
Paracyclopina sacklerae, Boxshall, Geoff A. & Jaume, Damià, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.279542 |
|
DOI |
https://doi.org/10.5281/zenodo.5693704 |
|
persistent identifier |
https://treatment.plazi.org/id/03E287D0-7007-AC78-FF25-FA58B1623D39 |
|
treatment provided by |
Plazi |
|
scientific name |
Paracyclopina sacklerae |
| status |
sp. nov. |
Paracyclopina sacklerae n. sp.
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Type material. Holotype female, 7 paratype females, 5 paratype males collected from sinkholes at Walengkabola village (5º 11.052’ S 122º 35.159’ E), Muna Island, Indonesia on 18 September 2007 by G.A. Boxshall and D. Jaume. Registration numbers: holotype [ MZB.Cru Cop.103], 4 paratype females and 3 paratype males [ MZB.Cru Cop.104] in Museum Zoologicum Bogoriense, 3 paratype females and 2 paratype males in Natural History Museum, London [ BMNH 2011.1167-1171].
Etymology. The species is named in honour of Mrs Theresa Sackler in recognition of her long term philanthropic support of the research and education work of the Natural History Museum, London.
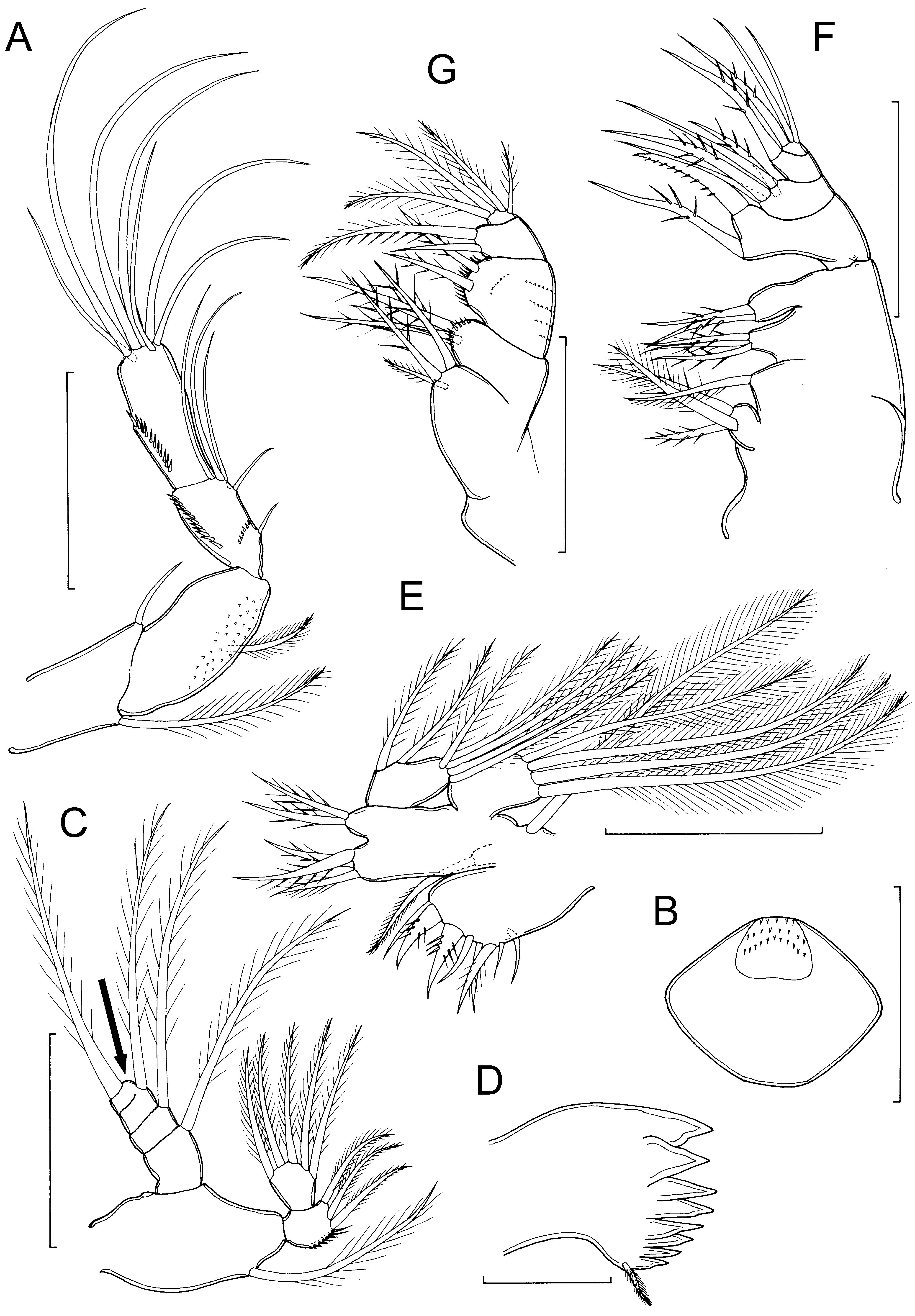
Description of adult female. Body cyclopiform ( Fig. 1 View FIGURE 1 A), divided into anterior prosome and posterior urosome with prosome–urosome boundary well defined at podoplean position. Prosome comprising cephalothorax and 4 free pedigerous somites; first pedigerous somite free, but partly concealed by posterior extension of dorsal cephalic shield. Epimeral angles of free second to fourth pedigerous somites rounded. Rostrum rounded, well developed. Nauplius eye not observed. Ratio of prosome to urosome length (including caudal rami) about 1.5:1. Urosome 5-segmented ( Fig. 1 View FIGURE 1 B), comprising fifth pedigerous somite, genital double-somite formed by fusion of genital and first abdominal somites, and 3 free abdominal somites. Genital apparatus comprising paired copulatory pores located within dorsolateral gonopores: seminal receptacles paired ( Fig. 1 View FIGURE 1 B). Egg sacs paired ( Fig. 1 View FIGURE 1 A), containing 7 to 9 eggs. Mean body length 0.549 mm, range 0.524 to 0.573 mm (based on 5 specimens).
Caudal rami ( Fig. 1 View FIGURE 1 B) 56μm long by 43μm wide, about 1.3 times longer than wide and bearing 6 caudal setae (seta I absent). Seta II plumose, 60 – 65 μm in length; outer distal angle seta III plumose, ca. 71–80 μm, but shorter than inner distal angle plumose seta VI (85–97 μm); inner apical seta V (268–274 μm) longer than outer apical seta IV (189–227 μm); dorsal seta VII plumose and 61–73 μm long.
Antennule 17-segmented ( Fig. 1 View FIGURE 1 C); probable segmental homologies as follows: segment 1 (I–II) double, segment 2 (III–V) compound, segment 3 (VI–IX) compound, segment 4 (X) free, segment 5 (XI) free, segment 6 (XII–XIV) compound, segment 7 (XV–XVI) double, segments 8 (XVII) to 16 (XXV) all free, apical segment 17 (XXVI–XXVIII) compound. Setal armature comprising: 3, 5, 7, 2, 2, 4, 2, 1, 1, 0, 1, 1 + aesthetasc, 1, 1, 1 + 1, 1 + 1, 6 + aesthetasc. Apical aesthetasc and adjacent seta arising from common base.
Antenna ( Fig. 2 View FIGURE 2 A) uniramous; with coxa and basis fused and separated from first endopodal segment by nonfunctional articulation; coxa-basis armed with long plumose inner seta and short outer seta representing exopod; endopod 3-segmented; first endopodal segment with 1 inner margin seta, second segment with 5 setae, the shortest and most proximal separated by gap from other 4, third endopodal segment with 7 setae; first endopodal segment ornamented with spinules on inner surface; second and third endopodal segments each ornamented with row of spinules adjacent to outer margin, second segment with additional transverse spinule row.
Labrum ( Fig. 2 View FIGURE 2 B) ovoid, with entire posterior margin; surface ornamentation of spinules on raised anterior zone.
Mandible ( Fig. 2 View FIGURE 2 C–D) comprising coxa with well developed gnathobase bearing numerous blades along oblique margin, and biramous palp: palp consisting of large basis bearing single plumose seta, 2-segmented endopod with 3 + 5 setae, and 4-segmented exopod bearing total of 5 sparsely plumose setae [figured specimen with only 4 exopodal setae, missing seta indicated by arrow in Fig. 2 View FIGURE 2 C].
Maxillule ( Fig. 2 View FIGURE 2 E) with large praecoxal arthrite bearing 8 marginal setal elements; coxal endite bearing single setae; coxal epipodite represented by long plumose seta; basis produced into 2 endites medially bearing 3 (proximal) and 2 (distal) spinulose setae; endopod comprising single expressed segment bearing 1, 1, 5 spinulose setae; exopod 1-segmented with 4 long plumose setae.
Maxilla well developed, 5-segmented ( Fig. 2 View FIGURE 2 F); praecoxa and coxa incompletely separated, bearing 4 inner margin endites, armed with 3, 1, 3, 2 spinulose setae (from proximal to distal); basis bearing powerful claw-like element and spinulose seta, ornamented with spinule rows; endopod 3-segmented, bearing 4, 1, 3 setal elements (from proximal to distal).
Maxilliped ( Fig. 2 View FIGURE 2 G) smaller than maxilla; 4-segmented; first segment (syncoxa) produced into two endites, proximal endite with 3 spinulose setae, distal endite armed with 2 setae and ornamented with row of spinules; second segment (basis) with 2 spinulose setae and row of spinules; endopod 2-segmented, first endopodal segment with 2 unequal spinulose setae and second with 4 spinulose setae.
Legs 1 to 4 biramous, with 3-segmented rami ( Fig. 3 View FIGURE 3 A–C) and with intercoxal sclerites present; free posterior margins of sclerites smoothly rounded, lacking any ornamentation on either surface in legs 1 to 3; intercoxal sclerite of leg 4 with 2 spinule rows on posterior surface. Spine and seta formula as follows:
coxa basis exopodal segments endopodal segments
leg 1 0-1 1-I I-1; I-1; III,I,4 0-1; 0-1; 1,2,3
leg 2 0-1 1-0 I-1; I-1; III,I,5 0-1; 0-2; 1,2,3
leg 3 0-1 1-0 I-1; I-1; III,I,5 0-1; 0-2; 1,2,3
leg 4 0-1 1-0 I-1; I-1; II,I,5 0-1; 0-2; 1,2,2
Coxa with spinule rows near outer distal angle in all legs (transversely orientated in leg 1 only); additional spinule rows present on coxa and basis of leg 4 only ( Fig. 3 View FIGURE 3 C). Outer margin spines on all exopods and inner spine on basis of leg 1 bilaterally ornamented with strips of serrated membrane. Inner margin of basis of legs 1 to 4 and outer margins of all endopodal segments ornamented with row of setules; outer margins of all exopodal segments ornamented proximally with row of spinules. Spinule rows located at articulation between all endopodal and exopodal segments. Terminal spine on exopod of leg 4 very powerfully developed ( Fig. 3 View FIGURE 3 C); spine 39μm in length with base 13μm in width, and 1.15 times longer than segment (length 34μm).
Fifth legs located ventro-laterally, comprising basal part incorporated into pedigerous somite armed with outer seta on small papilla and free exopodal segment ( Fig. 1 View FIGURE 1 B). Outer basal seta visible in dorsal view ( Fig.1 View FIGURE 1 A). Free exopodal segment 35μm long by 15μm in maximum width (measured at level of base of lateral spine), about 2.3 times longer than wide; bearing lateral margin spine (21μm long), outer distal spine (17μm), plumose apical seta (39μm) and inner distal spine (12μm); exopodal segment ornamented with sparse row of spinules along inner margin.
Description of adult male. Body cyclopiform ( Fig. 4 View FIGURE 4 A), divided into anterior prosome and posterior urosome as in female. Epimeral angles of free third pedigerous somites slightly produced. Ratio of prosome to urosome length (including caudal rami) about 1.7:1. Urosome 6-segmented, comprising fifth pedigerous somite, genital somite and 4 free abdominal somites. Genital somite ( Fig. 4 View FIGURE 4 B) bearing paired genital openings ventrally. Caudal rami ( Fig. 4 View FIGURE 4 A) 20μm long, by 16μm wide, about 1.25 times longer than wide and bearing 6 caudal setae (seta I absent), relative lengths of setae as for female. Mean body length 0.430 mm, range 0.408 to 0.461 mm (based on 3 specimens).
Antennules ( Fig. 4 View FIGURE 4 C) symmetrical, geniculate, 17-segmented; probable homologies: I–II, III–V, VI–VII, VIII, IX, X, XI, XII, XIII, XIV, XV, XVI, XVII, XVIII, XIX–XX, XXI–XXIII, XXIV–XXVIII; segment 11 (XV) with expanded sheath partly enclosing segment 12 (XVI). Setal armature comprising: 3, 4, 3, 2, 2, 0, 2, 1, 2, 1, 0, 1 (unilaterally spinulate), 1, 1 (unilaterally spinulate), 1 modified (fused to segment), 1 + 1 modified (fused to segment), 6 + aesthetasc.
Antennae to maxillipeds, and legs 1 to 4, as in female.
Fifth legs ( Fig. 4 View FIGURE 4 D) as in female except free exopodal segment 22μm long by 9μm in maximum width (measured at level of base of lateral spine), about 2.4 times longer than wide; bearing lateral margin spine (13μm long), outer distal spine (20μm), plumose apical seta (29μm) and inner distal spine (28μm); inner margin of segment smooth.
Remarks. The genus Paracyclopina was established in 1935 by Smirnov (1935) who designated his new species P. nana Smirnov, 1935 as the type species. At that time, the genus was included within the Cyclopinidae Sars, 1913 , a heterogeneous and probably paraphyletic family deliberately retained by Boxshall & Halsey (2004) even though proposals had already been made by Martínez Arbizu (2000a, b, 2001a, b) to break it up to form four other families, the Cyclopettidae Martínez Arbizu, 2000 , Giselinidae Martínez Arbizu, 2000 , Hemicyclopinidae Martínez Arbizu, 2001 and Psammocyclopinidae Martínez Arbizu, 2001 . Subsequently Martínez Arbizu (2006) proposed a fifth new family, the Schminkepinellidae , to include some other former members of the Cyclopinidae bringing the total number of “cyclopinid” families to seven (including the Cyclopinidae sensu stricto and the Pterinopsyllidae ). No comprehensive parsimony based test of the validity of the new families derived from the break up of the Cyclopinidae has yet been carried out. Indeed, no other publications have addressed this issue except Karanovic (2008) who considered that characters exhibited by the new cyclopinids he was describing from Australia exposed more of the characters used to support the break up of the Cyclopinidae sensu lato as unreliable and he recognised only the family Cyclopinidae . Clearly, uncertainty remains over the validity of some of these lineages as family level taxa.
Although its relationships with the family Oithonidae Dana, 1853 remain to be resolved, the Cyclopettidae is reasonably well supported and is recognised here. When he established the Cyclopettidae, Martínez Arbizu (2000a) tentatively included Paracyclopina as incertae sedis in his new family. Such caution was justified since all four of the species attributed to this genus were inadequately described. The four species recognised by Martínez Arbizu (2000a) as belonging to Paracyclopina were: P. nana , P. intermedia ( Sewell, 1924) , P. l o ng if urc a ( Sewell, 1924) and P. m i n u t a ( Sewell, 1934). The key characters used to support placement of Paracyclopina in a lineage with Cyclopetta Sars, 1913 were the 2-segmented endopod of the maxilliped and the lateral location of the leg 5 which comprises, in both sexes, one free exopodal segment carried on a slight pedestal representing the protopodal part of the limb that is incorporated into the somite. Martínez Arbizu (2000a) pointed out that the transformation of the inner apical and two outer margin spines on the exopod of leg 5 into setae was a synapomorphy of the core group of genera Cyclopetta , Paracyclopetta Wells, 1967 and Arctocyclopina Mohamed & Neuhof, 1985 . This transformation is not shared by Paracyclopina species, but the other synapomorphies support its placement in the Cyclopettidae ( Martínez Arbizu 2000a).
Cyclopetta orientalis Lindberg, 1941 was not mentioned by Martínez Arbizu (2000a): it was neither treated as a species of Cyclopetta , nor transferred to any other genus within the Cyclopinidae sensu lato. Although it was described as having a 1-segmented rather than a 2-segmented endopod on the maxilliped, it shares this form of laterally-located leg 5 and, in addition, has a 17-segmented antennule in the female, as in P. nana and P. m i nu ta. Accordingly Cyclopetta orientalis is here transferred to Paracyclopina as Paracyclopina orientalis ( Lindberg, 1941) n. comb. It is assumed here that the description of the maxilliped is mistaken and that P. orientalis has a 2- segmented endopod as found in the new species.
The new species, P. sacklerae n. sp., is closely related to P. orientalis . Both species possess a strikingly robust terminal spine on the exopod of leg 4 in both sexes and have three robust spines on the free exopodal segment of the female leg 5. No other Paracyclopina species share both of these features. There are several significant differences between these two species. Most obviously, the caudal rami are much shorter in P. sacklerae , only about 70% as long as the anal somite, whereas in P. orientalis the caudal rami are longer (125%) than the anal somite. The caudal rami themselves are more slender in P. orientalis , 3.5 times longer than wide compared to only 1.3 times longer than wide in P. sacklerae . Other differences include the shape of the antenna: in P. sacklerae the distal part of the antenna (second and third endopodal segments combined) is about equal in length to the proximal part (coxa-basis and first endopodal segment combined), whereas in P. orientalis the distal part is significantly shorter than the proximal part. The eggs sacs contained between 25 and 31 small eggs per sac in P. orientalis compared to 7 to 9 larger eggs per sac in the new species.
As highlighted by Martínez Arbizu (2000a), all four species he ascribed to Paracyclopina were found in brackish to fresh waters. Paracyclopina orientalis , transferred to Paracyclopina here, was collected in shallow salty waters near to the coast at Pondichery (“des mares saumâtres peu profondes, près du bord de la mer à Pondichéry ”) and in a lagoon at Oupalom ( Lindberg 1941). It seems likely that this species also inhabits brackish waters. Finally, the new species was found in small sink holes, about 2 m deep, located in the coastal limestone platform only 5 to 10 m inland from the shoreline on Muna Island. During low tide many of the sinkholes appeared dry, but at high tide, water was clearly visible in most of them. The copepods were caught using a long-handled plankton net in a water column that was up to 50 or 60 cm deep. Salinity measurements were not taken, but given the tidal influence and the proximity to the coast, it is highly likely that the water in the sinkholes was brackish but close to fully marine. The species of Paracyclopina appear to prefer low salinity and brackish water habitats, although P. m i n u t a was reported from “absolutely fresh” water in the Hooghly River, according to Sewell (1934).
The genus is known only from the Indo-Pacific. Four Paracyclopina species were described from the coast of India: P. longifurca and P. intermedia were originally described from Chilka Lake ( Sewell 1924) and the former is also known from the South India coast and from the canal system of the salt lakes at Chingrighatta ( Sewell 1934); P. minuta is known from the Hooghly River which bounds the salt lakes and associated canal system near Chingrighatta ( Sewell 1934); P. orientalis was collected at Pondichery and Oupalom ( Lindberg 1941). The new species, P. sacklerae , is from Indonesia. The type species, P. nana , was described from the mouth of the Suifun river near Vladivostok ( Smirnov 1935) and is also known from the coasts of Japan, South Korea and China ( Tai & Chen 1979; Chang 2009; Ueda et al. 2001).



| MZB |
Museum Zoologicum Bogoriense |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paracyclopina sacklerae
| Boxshall, Geoff A. & Jaume, Damià 2012 |
Cyclopetta orientalis
| Lindberg 1941 |
Paracyclopina orientalis (
| Lindberg 1941 |