
Macrothrix oviformis Ekman, 1900
|
publication ID |
https://doi.org/10.1080/00222930701689937 |
|
persistent identifier |
https://treatment.plazi.org/id/03E32C46-B146-FF89-FE60-FA10D9E410BB |
|
treatment provided by |
Felipe |
|
scientific name |
Macrothrix oviformis Ekman, 1900 |
| status |
|
Macrothrix oviformis Ekman, 1900 View in CoL
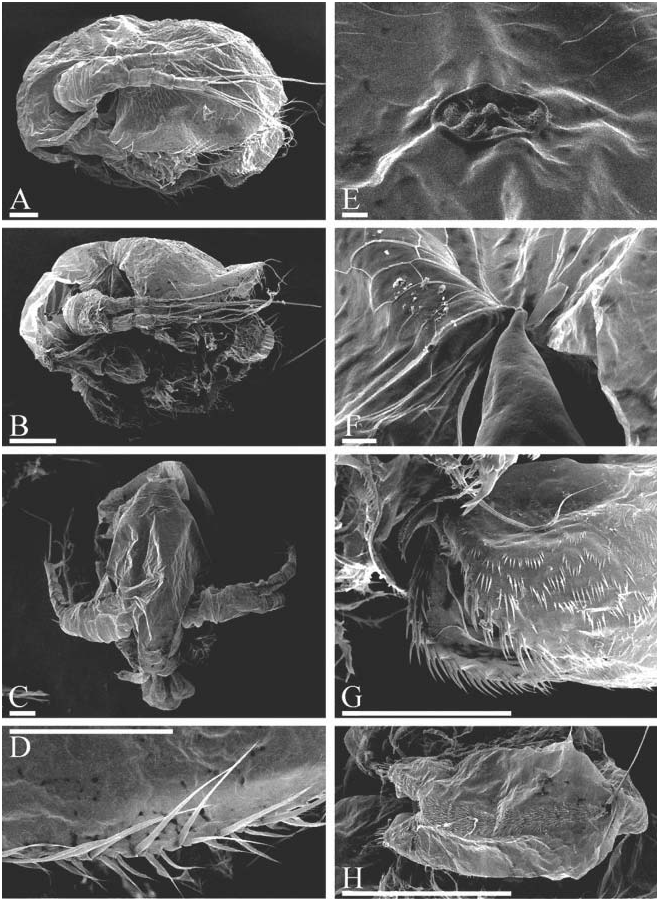
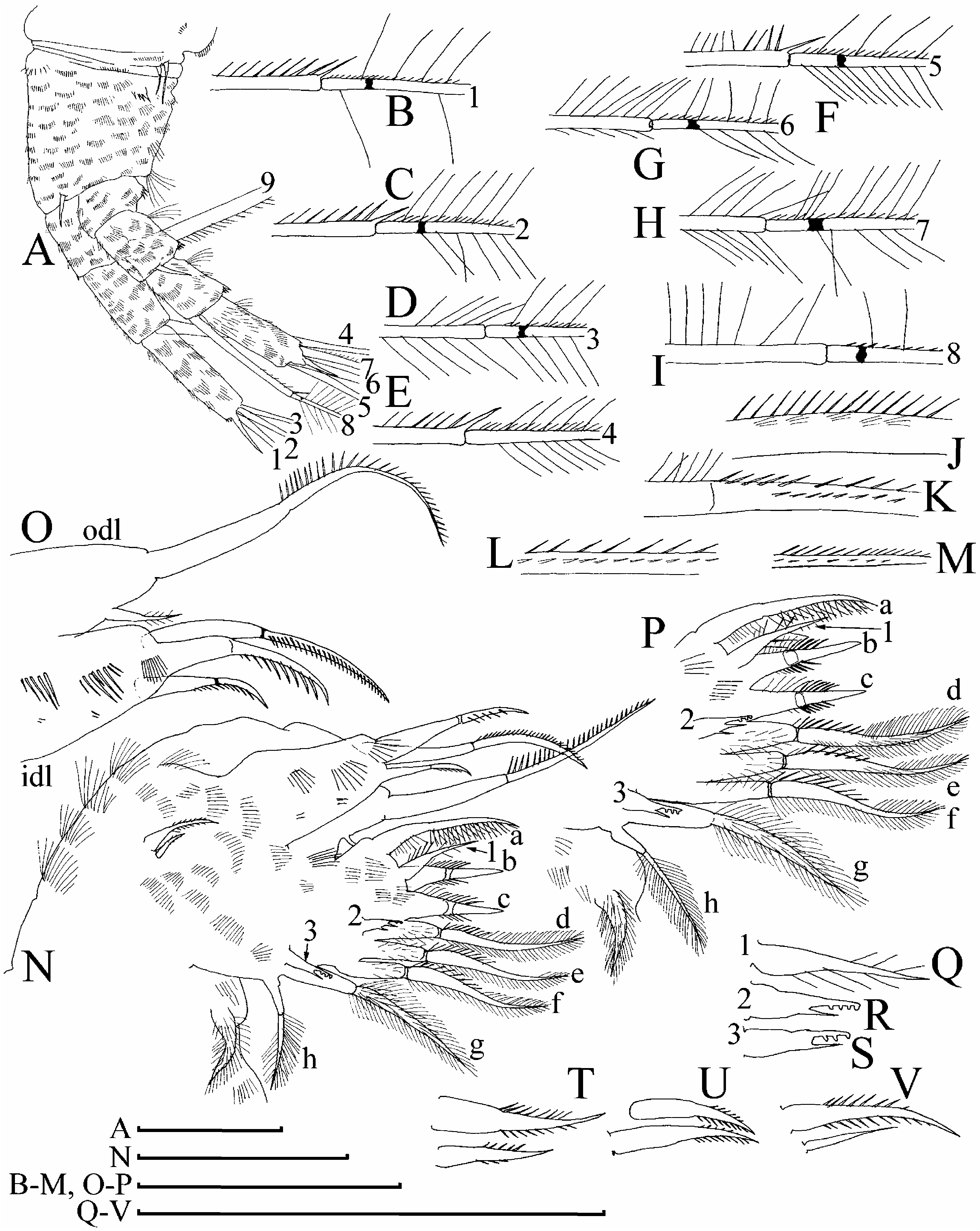
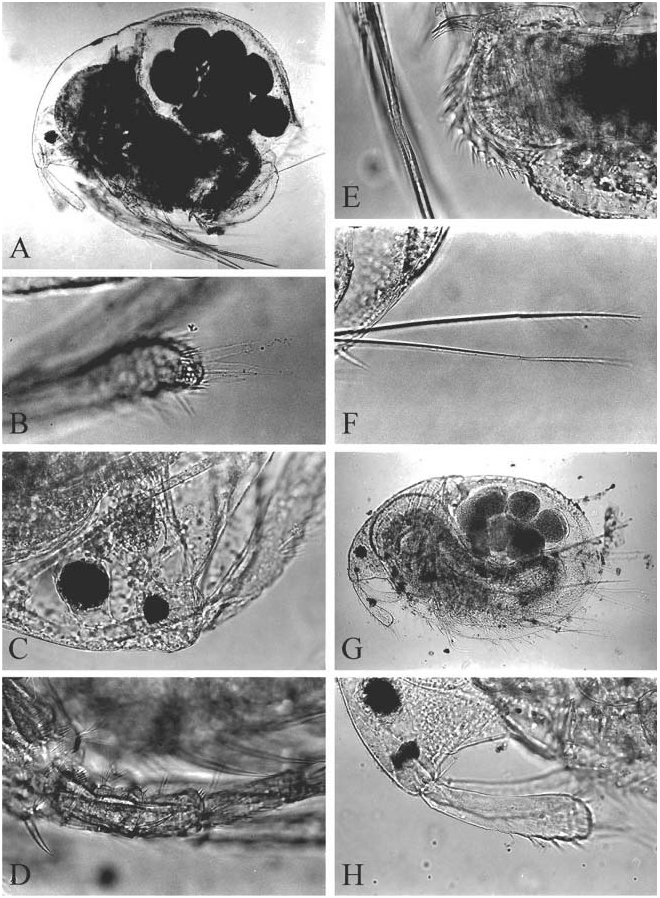
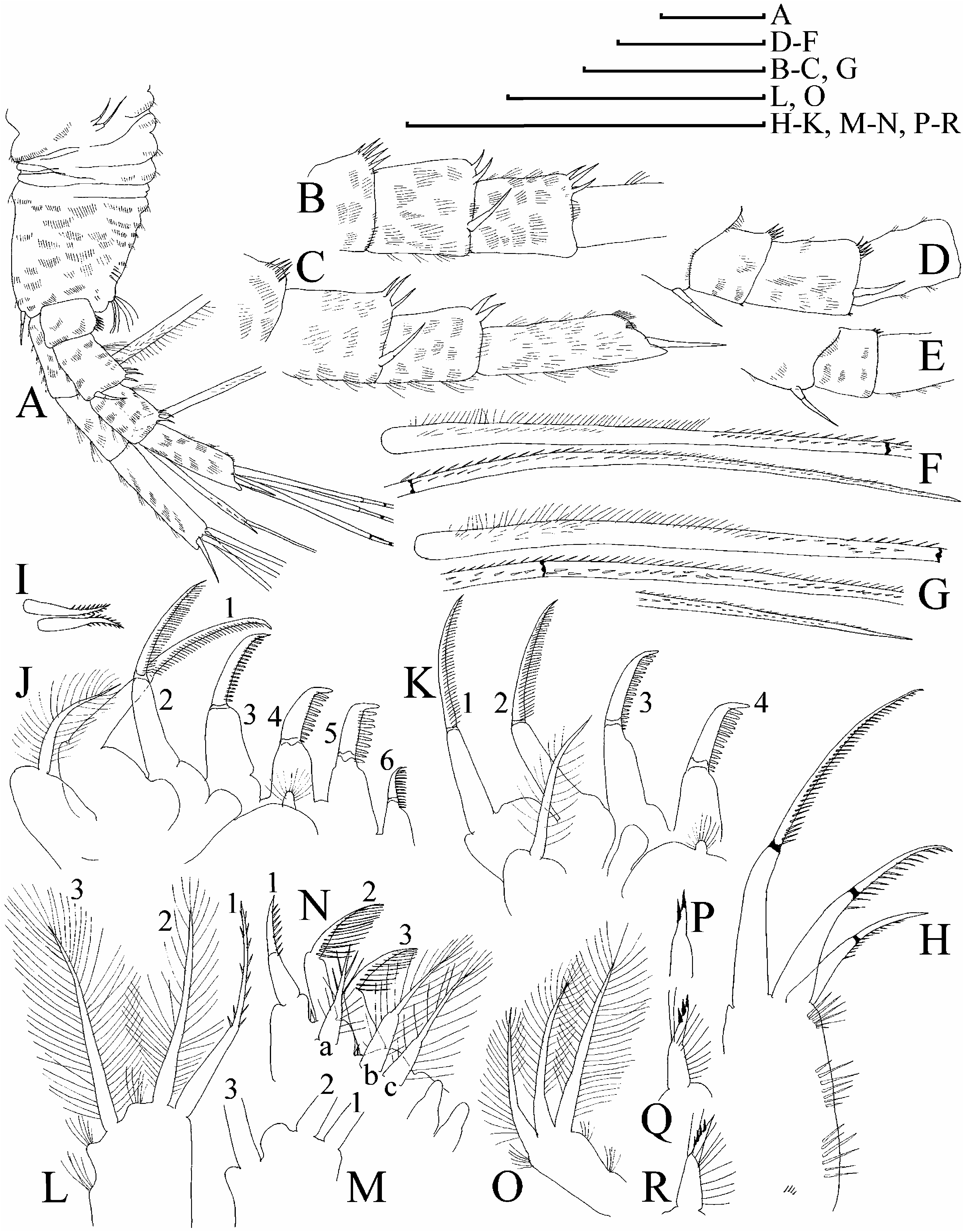
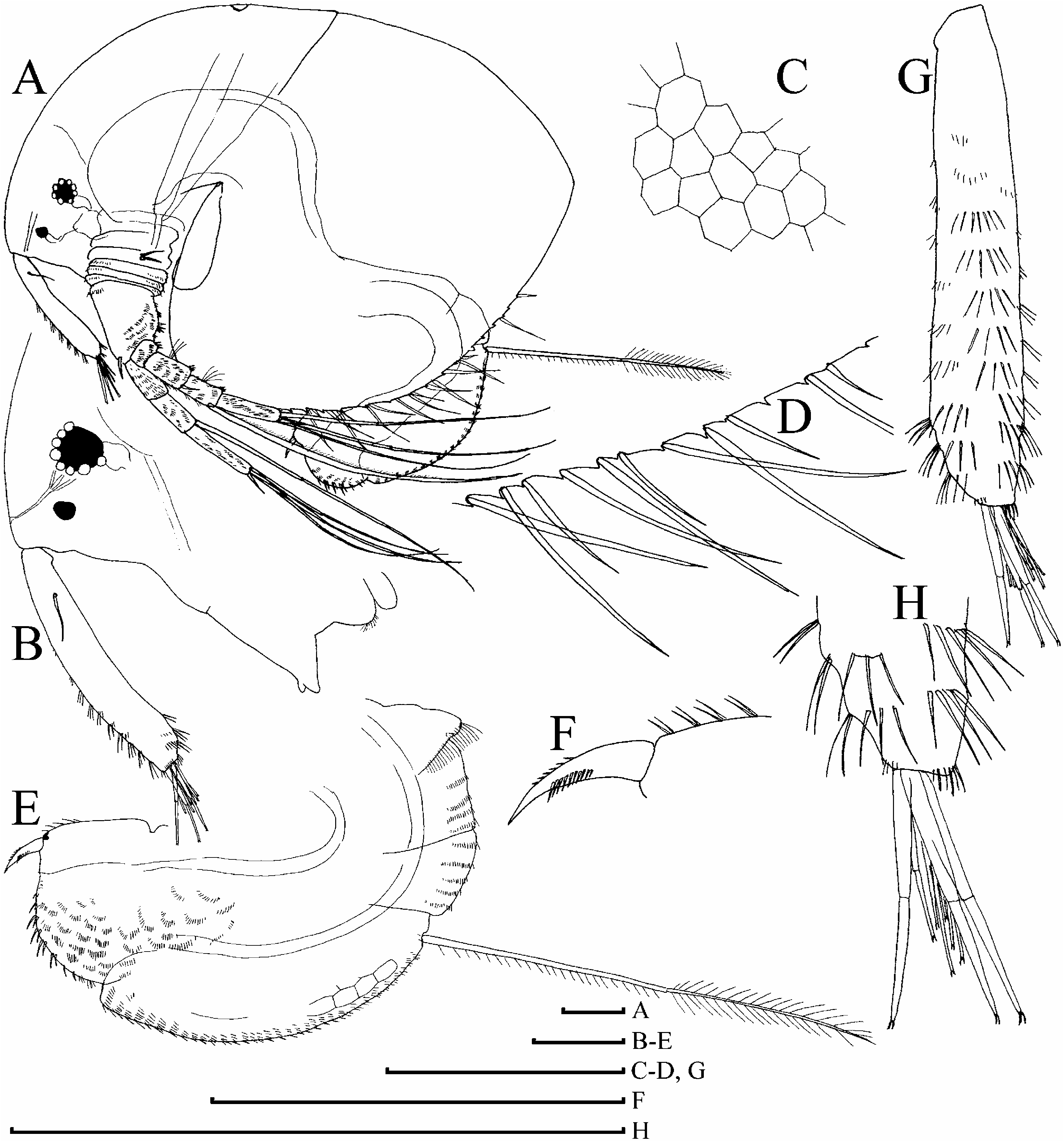
( Figures 6D–H View Figure 6 , 7–12 View Figure 7 View Figure 8 View Figure 9 View Figure 10 View Figure 11 View Figure 12 , 13A–F View Figure 13 )
Macrothrix oviformis Ekman 1900, p 71 View in CoL –73, Plate 4, Figures 17–19 View Figure 17 View Figure 18 View Figure 19 ; Olivier 1962, p 225, Plate 19, Figures 3–5 View Figure 3 View Figure 4 View Figure 5 ; Smirnov 1976, p 91, 94, Figure 64.
Macrothrix ciliata Vávra 1900, p 18 View in CoL –19, Figure 3 View Figure 3 ; Olivier 1962, p 225, Plate 16, Figures 6 View Figure 6 , 7 View Figure 7 . Macrothrix odontocephala Daday 1902, p 272 View in CoL –274, Plate 9, Figures 18–20 View Figure 18 View Figure 19 View Figure 20 ; Olivier 1962, p 225, Plate 18, Figure 6 View Figure 6 , Plate 19, Figures 1 View Figure 1 , 2 View Figure 2 ; Smirnov 1976, p 95, Figure 66.
Macrothrix propinqua Sars 1909, p 5 View in CoL –15, Plate 1; Pesta 1928, p 80; Olivier 1962, p 230, Plate 19, Figures 6 View Figure 6 , 7 View Figure 7 , Plate 20, Figure 1 View Figure 1 .
Macrothrix hirsuticornis Norman and Brady View in CoL in Ekman 1905, p 7 –8; Harding 1941, p 319; Olivier 1962, p 225 –226, Plate 17, Figures 1–3 View Figure 1 View Figure 2 View Figure 3 ; Dartnall and Hollwedel 2007, p 1273 – 1274, Figures 18 View Figure 18 , 19 View Figure 19 .
? Macrothrix inflata Daday 1902, p 271 View in CoL –272, Plate 10, Figures 13–16 View Figure 13 View Figure 14 View Figure 15 View Figure 16 , Plate 11, Figure 1 View Figure 1 ; Olivier 1962, p 226, Plate 17, Figures 4–6 View Figure 4 View Figure 5 View Figure 6 ; Smirnov 1976, p 89, Figure 60.
not Macrothrix odontocephala Daday 1902 View in CoL in Daday 1906, p 39; Daday 1913, p 321, 345, 347, 348, 351, 352, 356.
Type locality
‘‘...einer Lagune in der Nähe von Rio Turbio’ ’ ( Ekman 1900) in Santa Cruz, Argentina .
Type material
Most probably lost. Absent from Ekman’s Collection in SMNH ( Kotov and Gololobova 2005).
Type series of junior synonyms examined here
Macrothrix ciliata Vávra, 1900 View in CoL — neotype (selected here): juvenile female 0.53 mm from Port Stanley, Falkland Islands, coll. 18 August 1902 by South Polar Expedition of 1901–1903, collection of S. Ekman, tube SMNH 5874. Neotype label: ‘‘ Macrothrix ciliata Vávra, 1900 View in CoL , 1 juv. ♀ from Port Stanley, Falkland Islands, NEOTYPE’’.
Macrothrix inflata Daday, 1902 View in CoL — syntypes: four females from unknown locality in Santa Cruz, Argentina, coll. by F. Sylvestri, slide DAD II/P-418. We found that Smirnov (1992) was correct when he said that the slide is dried, and the specimens are in a bad state.
Macrothrix odontocephala Daday, 1902 View in CoL — type material: lost. Specimens in slide DAD D V-96; II/P-420 and tube DAD D 1917-142; II-457 (see Forró and Frey 1982), misidentified as ‘‘ M. odontocephala View in CoL ’’ by Daday (1906), contain material from Mongolia, apparently belonging to Palaearctic M. hirsuticornis View in CoL .
Macrothrix propinqua Sars, 1909 View in CoL — lectotype (selected here): parthenogenetic female 1.05 mm from unknown locality in South Georgia, GOS F 12298a. Paralectotypes (selected here): 127 parthenogenetic, ephippial females and males, the same locality with the lectotype, GOS F 12298b; one dissected parthenogenetic female on eight slides, GOS F 12298c-j; one dissected male on five slides, GOS F 12298k-o.
Other material examined
Continental South America. Argentina ( Santa Cruz): a pond (or an old river channel) at Mission Salesiana, Highway 3, 12 km north of Rio Gallegos , coll. 19 January 1989 by D. G. Frey, DGF 8713 ; drainage channel, Highway 3, 2 km north of P. Maria, coll. 20 January 1989 by D. G. Frey, DGF 8714 ; seepage pools in dry bottom of Brazo Rico of Lago Argentino , coll. 25 January 1989 by D. G. Frey, DGF 8745 ; Altwässer, Rio Coig , 19 km west of Estancia La Vanguardia, coll. 26 January 1989 by D. G. Frey, DGF 8748 ; a small creek, tributary of Rio Coig , west of Esperanza, coll. in 26 January 1989 by D. G. Frey, DGF 8749 ; Lago Largo, on road from Rio Turbio to Rio Gallegos , coll. 27 January 1989 by D. G. Frey, DGF 8755 ; pools near crossing of Rio el Zorro on road from Rio Turbio to Rio Gallegos, coll. 27 January 1989 by D. G. Frey, DGF 8757 ; valley pond dammed by road, 13 km north of Estancia Monte Dinero on road from C. Virgenes to Rio Gallegos, coll. 28 January 1989 by D. G. Frey, DGF 8759 . Chile ( Magallanes): Laguna Melliza, Parque Nacional de los Torres del Paine , coll. 14 January 1989 by D. G. Frey, DGF 8764 ;
grassy meadow, 8 km north of turnoff to Monte Aymond, coll. 15 January 1989 by D. G. Frey, DGF 8780.
Tierra del Fuego ( Argentinian part). Freshwater lake near ocean, on Pt. Almte Broun Road, coll. 14 January 1989 by D. G. Frey, DGF 8666 ; Lago Fagnano, coll. 15 January 1989 by D. G. Frey , DGF 8679 ; a lagoon along Highway 3, 8 km north of Hosteria Kaiken, coll. 15 January 1989 by D. G. Frey, DGF 8689 ; northern and middle pond of three ponds, a few km south of Rt. E crossing of Rio Mac Lennan, coll. 17 January 1989 by D. G. Frey, DGF 8701 and 8702; flooded grassland near where Rt. B takes off from Highway 3, coll. 18 January 1989 by D. G. Frey, DGF 8703 .
Falkland Islands. Unknown water body near Port Stanley, coll. 18 August 1902 by South Polar Expedition of 1901–1902, SMNH 5875 View Materials and 32918 ( neotype locality) .
South Georgia. Unknown locality, NHM 1962.4.3.2.
Signy Island (South Orkney Island group). Heywood Lake, coll. March 1964 by R. B. Heywood, BAS 64 View Materials /257; Pumphouse Lake , coll. February 1964 by R . B. Heywood, BAS 64 View Materials /260; unknown locality, coll. 1964 by R. B. Heywood , BAS no number.
Antarctic Peninsula. Horseshoe harbour (‘‘Penola’’?), Graham Land, coll. 19 February 1937 by British Graham Land Expedition 1934–1937, NHM 1940.3.5.41–42.
Amended diagnosis
Parthenogenetic female. In lateral view body subovoid (height/length50.71– 0.78 in largest adults), cervical depression expressed (although shallow), posterior boundary of head forming or not forming a ‘‘step’’ on dorsum, dorsal margin of valves without any serration or with minute serration ( Figures 6H View Figure 6 , 7A, H, J–L View Figure 7 , 8A View Figure 8 , 9A View Figure 9 , 13A View Figure 13 ). Posterodorsal angle obtuse, lies in large adults ventral to middle of body height. Ventral margin ( Figures 8C, D View Figure 8 , 9E–H View Figure 9 ) with setae as in M. boergeni . In anterior view, body compressed laterally, with a distinct dorsal keel ( Figure 9B View Figure 9 ). No dome above eye. Ocellus small ( Figure 13C View Figure 13 ). Dorsal organ small or of moderate size. Labrum with a thick, rounded, moderately projected apex, supplied with few low tubercles, which may be absent ( Figures 7B View Figure 7 , 8B View Figure 8 , 9C View Figure 9 ).
Abdomen with a distinct, setulated dorsal projection ( Figures 8E View Figure 8 , 9I View Figure 9 ). Postabdomen subovoid, with rounded distal extremity, without a ‘‘heel’’ basally, with ill-defined reticulation on sides, or without it ( Figures 8E View Figure 8 , 9I View Figure 9 ). Ventral margin slightly convex, with few series of short denticles ( Figures 8F View Figure 8 , 9L View Figure 9 ). Dorsal margin distinctly bilobed; preanal margin with transversal series of minute setules, anal margin with groups of thicker setules ( Figure 13E View Figure 13 ). Postabdominal seta with distal segment densely armed with long setules; proximal segment with numerous, relatively long setules (although shorter than on distal one) ( Figures 7E View Figure 7 , 8E View Figure 8 , 9M View Figure 9 , 13F View Figure 13 ). On outer side of postabdominal claw, a series of 7–14 fine spinules, with size increasing distally; medial row of about six to eight denticles; inner row with numerous denticles, organized in two successive series, subdivided by a specially large denticle ( Figures 7C, D View Figure 7 , 8F View Figure 8 , 9J–L View Figure 9 ).
Antenna I widened distally, slightly curved, without a subapical external angulation; sensory seta at distance of about 2–2.5 antennular diameters (at base) from antenna I joint; on anterior face about five to six transverse rows of strong spinules, and rows of finer setules, but no reticulation ( Figures 7B View Figure 7 , 8B, G View Figure 8 , 9N, O View Figure 9 , 13B View Figure 13 ). Nine short aesthetascs (longest shorter than half of antenna I length), two of them significantly larger than the rest. On the latter, sometimes there are some thin-walled, delicate structures ( Figure 9O View Figure 9 ), which could be additional sensory elements, but there is a chance that these are only epibiotic bacteria, as in the case of M. boergeni . Antenna II with distal burrowing spine on basal segment longer than proximal segment of exopod ( Figures 10A View Figure 10 , 13D View Figure 13 ). Length of all apical swimming setae subequal, their armature represented in Figure 10B–I View Figure 10 . Lateral seta on proximal endopod segment larger than other setae, with robust denticles in the middle ( Figures 7F–G, I, N, P View Figure 7 , 8I–K View Figure 8 , 10J– N View Figure 10 ). A spine on second segment of exopod approximately half as long as next segment. On posterior side of segments 1–3 of exopod there are minute additional denticles.
Limb I with longest apical seta of outer distal lobe ( Figure 11A, B View Figure 11 ) having distal segment unilaterally armed with relatively fine, spaced setules; inner-distal lobe with three bisegmented setae of different size, unilaterally setulated in distal part, robustness of armature different in different setae, smallest one with whole distal segment setulated ( Figure 11C View Figure 11 ), or these setules present only in proximal portion of distal segment ( Figure 11B View Figure 11 ); two ejector hooks of similar size; a remainder of gnathobase I with single fully setulated setae. On limb II ( Figure 11D, E View Figure 11 ), scrapers 1–2 with delicate feathering, scrapers 3–7 with robust denticles of size characteristic for the genus; a solitary posterior seta near gnathobase present; filter plate II with four setae, without a rudiment of fifth seta. On limb III ( Figure 11F View Figure 11 ) epipodite small, exopod with a distal group of three long setae, seta 1 shortest, armed with short setules; setulated projections proximally to seta 3 and between setae 2 and 3; inner-distal limb portion ( Figure 11G View Figure 11 ) with seta 1 with short and robust denticles; seta a with fine setules basally and robust spinules distally, seta b longer than c; basal endite posteriorly with four soft setae. Limb IV ( Figure 11H View Figure 11 ) with exopodite small, bearing only a distal group of three bilaterally feathered setae of subequal size; on inner-distal portion of this limb ( Figure 11I View Figure 11 ) seta 1 with strong setules basally and two to five robust denticles distally; posteriorly row of five long setae. On limb V there are three setae on inner margin ( Figure 11J View Figure 11 ).
Juvenile female. Body less high (height/length50.64–0.68), subquadrangular, with almost straight dorsal margin ( Figures 6E–G View Figure 6 , 7O View Figure 7 , 8L, M View Figure 8 ).
Ephippial female. In contrast to adult parthenogenetic female, body less high (height/ length50.65–0.72), postero-dorsal angle more prominent and lies in level of middle of body height ( Figure 12A View Figure 12 ). Body less compressed laterally and lacking a dorsal keel ( Figure 12B View Figure 12 ). Dorsal portion of valves forming an ephippium, slightly pigmented in brownish colour, without clear border between it and rest of valves. Dorsal wall of carapace forms a special dark, chitinized plate. Two eggs in ephippium.
Male. Body subquadrangular in lateral view. Apex of labrum with a series of tubercles; a reticulation on sides of postabdomen fully absent, gonopores open on its distal margin, distalmost denticle in median series on postabdomen claw significantly larger than the rest. Antennular sensory seta located there at distance of about 1.5 antennular diameters (at base) from antenna I joint; at the same level, a large male seta, with length of half of antenna I length, bisegmented, with basal segment short and naked, and distal segment fully setulated; 14 relatively short terminal aesthetascs, two aesthetascs slightly larger than the rest. Limb I with inner-distal lobe bearing four setae, additional male seta large; copulatory hook large, with three ridges distally, copulatory brush seta on a low pedestal, bisegmented, with blunt tip to distal segment.
Size. Females up to 1.14 mm, males up to 0.56 mm.
Redescription of adult male
Body subquadrangular in lateral view, height/length50.63–0.69, dorsal margin without a distinct cervical depression, no ‘‘step’’ in posterior boundary of head ( Figure 12C View Figure 12 ). Dorsal margin of valves not elevated significantly above dorsal margin of head. Postero-dorsal angle obtuse, rounded, lies dorsal to middle of body height.
Head with dorsal margin evenly convex, no dome above eye; ventral margin slightly concave to straight, without ridges, no projection at base of labrum ( Figure 12D View Figure 12 ). Compound eye large, ocellus even smaller than in female (its size about one-third of eye diameter), located closely to tip of rostrum. Dorsal organ small, located on posterior part of head.
Labrum in lateral view approximately triangular, with a moderately projected apex bearing several small tubercles, and setulated distal labral plate.
Valve surface with a delicate reticulation. Dorsal margin without any serration, ventral margin as in female, but marginal setae relatively longer ( Figure 12E, F View Figure 12 ).
Postabdomen in lateral view ( Figure 12G View Figure 12 ) with subquadrangular distal portion and without a ‘‘heel’’ basally. Ventral margin inflated, without series of denticles. Dorsal margin distinctly bilobed, preanal margin regularly convex, with short transversal series of minute setules. On anal margin groups of setules with size significantly larger than those on postanal margin, laterally to them there are series of finer setules. A reticulation on sides of postabdomen fully absent. Gonopores open on distal margin, ventrally to postabdominal claws.
Postabdominal seta longer than postabdomen, with short distal segment, densely armed with long setules; basal segment with numerous, shorter setules.
Postabdominal claw small, directed distally, less elongated than in female, slightly and regularly bent dorsally, with pointed tip and wide base in lateral view ( Figure 12H View Figure 12 ). On claw, a series of seven to ten fine denticles on external side; medial row with about five to six denticles, distalmost one significantly larger than the rest.
Antenna I almost straight, with maximum width in basal portion, this part a homologue of subapical external angulation; antennular sensory seta located at distance of about 1.5 antennular diameters (at base) from antenna I joint ( Figure 12 View Figure 12 IJ). On anterior surface of antenna I, at level of sensory seta, a large male seta, half length of antenna I, bisegmented, with basal segment short and naked, and distal segment fully setulated. About six to seven transverse rows of spinules on anterior surface of antenna I, but no reticulation. Fourteen relatively short terminal aesthetascs (longest one-quarter length of antenna or even shorter), apex of each with a conical depression on distal side ( Figure 12K, L View Figure 12 ), two aesthetascs slightly larger than the rest.
Antenna II with relative size larger than in female; basal segment of exopod relatively shorter than in female; distal burrowing spine on basal segment and a spine on the second segment of exopod significantly longer than in female ( Figure 10O View Figure 10 ).
Limb I with outer distal lobe as in female; inner-distal lobe with four setae, additional male seta large; copulatory hook large, with three ridges distally ( Figure 12M View Figure 12 ). Copulatory brush seta on a low pedestal, bisegmented, with blunt tip to distal segment.
Juvenile (pre-reproductive) male. Body shape ( Figure 7Q View Figure 7 ) similar to adult; height as in adult male (height/length50.61–0.66), postabdomen ( Figure 7R View Figure 7 ) as in female, distal denticle of median row on postabdominal claw ( Figure 7S View Figure 7 ) not as large as in adult male; antenna I with nine aesthetascs and shorter male seta ( Figure 7T, U View Figure 7 ); male seta on inner-distal lobe of limb I smaller and located at a distance from other setae, copulatory hook shorter than in adult ( Figure 7V View Figure 7 ).
Size. Parthenogenetic females from Lago Largo, on road from Rio Turbio to Rio Gallegos, Santa Cruz ( DGF 8755) 0.37–0.71 mm; from ponds a few km south of Rt. E crossing of Rio Mac Lennan, Tierra del Fuego ( DGF 8701) 0.43–0.90 mm; from Port Stanley, Falkland Islands ( SMNH 32918) 0.50–1.06 mm; parthenogenetic females from unknown locality in South Georgia ( GOS F 12298b) 0.36–1.05 mm, ephippial females 0.67– 0.79 mm, juvenile males 0.42–0.49 mm, adult males 0.47–0.56 mm. According to Sars (1909), female length reaches 1.14 mm.
Taxonomic notes. The southernmost portion of the South American continent, South Atlantic (Subantarctic) islands and Antarctic Peninsula are inhabited by only a single hirsuticornis -like species, for which several formal names were suggested: M. oviformis Ekman, 1900 , M. ciliata Vávra, 1900 , M. inflata Daday, 1902 , M. odontocephala Daday, 1902 , and M. propinqua Sars, 1909 .
Two species ( M. oviformis Ekman and M. ciliata Vávra ) were published within the same year; the article of Ekman (1900) appeared on 22 October 1900 (this information present on the title page of the issue where it was published). Attempts to establish when Vávra’s (1900) paper appeared, including special requests to several libraries in Germany (including Hamburg, where it was published), were unsuccessful. According to case 21.3.2 of the International Code of Zoological Nomenclature ( International Commission on Zoological Nomenclature 2000), the latter article must be regarded as published on ‘‘the last day of the year’’, namely on 31 December 1900. So, M. oviformis Ekman, 1900 has priority, and M. ciliata is its junior synonym.
First description of M. oviformis Ekman, 1900 was relatively detailed for that time. The distinctive characters listed by Ekman (1900, p 71–73) are now considered of dubious value. The first description contains some important characters such as (1) a depression bordering dorsal margin of head and valves; (2) nine aesthetascs of different size; (3) two dorsal projections on abdomen, and his illustrations adequately represent others such as (1) shallow cervical depression; (2) presence of numerous setules on proximal segment of postabdominal seta; (3) long setules on distal segment of postabdominal seta. Specimens very similar to that illustrated by Ekman (1900) were found at localities close to the type locality (Rio Turbio), as well as at other localities in continental South America.
Macrothrix ciliata Vávra, 1900 View in CoL was described from the Falkland Islands. Most of the characters listed by Vávra (1900, p 18–19) are characters of a juvenile hirsuticornis View in CoL -like species. A neotype of this taxon is here selected from Ekman’s (1905) sample collected in the vicinity of the type locality; a juvenile female is selected, confirming Vávra’s ideas on this species. There is no doubt that Macrothrix ciliata View in CoL is a junior synonym of M. oviformis View in CoL .
Macrothrix odontocephala Daday, 1902 View in CoL is an apparent junior synonym of M. oviformis View in CoL . Similar morphotypes with a tooth-like projection, or with a ‘‘monk’s hood’’ (in terminology of Fox 1962) on posterior portion of head (‘‘ var. groenlandica Lilljeborg, 1901 View in CoL ’’) are known also for Palaearctic M. hirsuticornis View in CoL ( Fox 1962; Flössner 1967, 1972, 2000). Several samples have been examined from Santa Cruz, from where the taxon was described, revealing several ‘‘ odontocephala View in CoL ’’-like populations (with a tooth at posterior border of head) of M. oviformis View in CoL . In contrast, Daday’s (1906) ‘‘ M. odontocephala View in CoL ’’ from Mongolia is a ‘‘toothed’’ morphotype of M. hirsuticornis View in CoL .
Macrothrix inflata Daday, 1902 View in CoL is more problematic. Females from a single population from continental South America (DGF 8780, unfortunately, with few specimens), lacking a depression between head and valves, also have some other specific traits ( Figure 7A–G View Figure 7 ): (1) setules on seta on proximal segment of antennal endopod are relatively small, and (2) the dorsal organ is very large as compared with populations with clear border between head and valves. This may indicate that M. inflata View in CoL is a valid species, but this conclusion must be confirmed by examination of better material; however, there is a high probability that the inflata View in CoL -like morphotype is only an extreme example of the oviformis View in CoL variability. Ekman (1900) clearly stated that M. oviformis View in CoL has a distinct border between head and valves, so in the case that two taxa are recognized in South America, M. oviformis View in CoL is widely distributed and M. inflata View in CoL rare.
Macrothrix propinqua Sars, 1909 View in CoL was described in detail, but no real differences from other species described from the South Atlantic region were reported. Re-examination of Sars’ (1909) type material led to the conclusion that his taxon is an apparent synonym of M. oviformis View in CoL .
Two other species described from the southern part of South America, M. goeldi Richard, 1897 View in CoL and M. magna Daday, 1902 View in CoL , are not members of the hirsuticornis View in CoL -group. Macrothrix goeldi Richard, 1897 View in CoL is a member of the laticornis- or spinosa- group, because the first description ( Richard 1897, p 287–289) contains a clear reference to distinct denticulation at the dorsal margin of the valves. Macrothrix magna Daday, 1902 View in CoL was regarded by Smirnov (1992) as a junior synonym of M. hirsuticornis View in CoL , but this is incorrect. The male of M. magna View in CoL has a very thin antenna I with a great distance between male seta and antennular sensory seta. This is apparently a relative of the triserialis-rosea group, sensu Dumont et al. (2002). At the same time, M. magna View in CoL has postabdominal seta with a long distal segment, so the former is not a junior synonym of M. triserialis View in CoL s. str. or M. elegans Sars, 1901 View in CoL (the latter is one of the most common species in the more southern portion of South America; Kotov et al. 2004). Macrothrix magna View in CoL needs to be revised, as a species outside of the hirsuticornis- like group. Olivier (1962) listed differences between all species described by previous authors, but these differences are dubious.
Distribution. Southernmost portion of continental South America, Tierra del Fuego, Falkland Islands, South Georgia, South Orkney Islands, and the Antarctic Peninsula. All records from the South Atlantic listed by Pugh et al. (2002) refer to this species. It is necessary to note that in these localities M. oviformis is a quite common species. Most probably Macrothrix from the South Shetland Islands ( Toro et al. 2007) and Palmer Land ( Pugh et al. 2002) also belong to this species.












No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Macrothrix oviformis Ekman, 1900
| Kotov, Alexey A. 2007 |
Macrothrix propinqua
| Olivier SR 1962: 230 |
| Pesta O 1928: 80 |
| Sars GO 1909: 5 |
Macrothrix odontocephala
| Daday E 1913: 321 |
| Daday E 1906: 39 |
Macrothrix hirsuticornis
| Dartnall HJG & Hollwedel W 2007: 1273 |
| Olivier SR 1962: 225 |
| Harding JP 1941: 319 |
| Ekman S 1905: 7 |
Macrothrix inflata
| Smirnov NN 1976: 89 |
| Olivier SR 1962: 226 |
| Daday E 1902: 271 |
Macrothrix oviformis
| Smirnov NN 1976: 91 |
| Olivier SR 1962: 225 |
| Ekman S 1900: 71 |
Macrothrix ciliata Vávra 1900 , p 18
| Smirnov NN 1976: 95 |
| Olivier SR 1962: 225 |
| Olivier SR 1962: 225 |
| Daday E 1902: 272 |
| Vavra W 1900: 18 |