
Macrothrix ruehei, Kotov, 2007
|
publication ID |
https://doi.org/10.1080/00222930701689937 |
|
persistent identifier |
https://treatment.plazi.org/id/03E32C46-B16A-FFB4-FE16-FB48DFD3172E |
|
treatment provided by |
Felipe |
|
scientific name |
Macrothrix ruehei |
| status |
sp. nov. |
Macrothrix ruehei View in CoL sp. nov.
( Figures 17 View Figure 17 , 18 View Figure 18 ) Macrothrix hirsuticornis Norman and Brady in Rühe 1914, p 55–56 (only populations from Île Amsterdam); Brehm 1958, p 29, Figure 6 View Figure 6 .
Etymology
The species is named after Friedrich Eduard Rühe, well-known German investigator of Cladocera , who identified this species as M. hirsuticornis from Île Amsterdam ( Rühe 1914).
Type locality
A pond at Baie Americaine , Île de la Possession, Crozet Islands, French Subantarctic Territories. The type series was collected 24 February 1968 by L. Davies .
Type material Holotype: a parthenogenetic ♀ in 90% alcohol, NHM 2004.2309. Label of holotype:
‘‘ Macrothrix ruehei n.sp.; pond, Baie Americaine , Possession Is., Crozet Islands, coll. 24.02.1968 by L. Davies, 1 parth. fem., HOLOTYPE’’. Paratypes (all from Crozet): 151 parth. ♀♀ from the type locality, coll. 24 February 1968 by L. Davies, NHM 1968.7.10.2 (tube); one parth. ♀ from the type locality, coll. 24 February 1968 by L. Davies, NHM 1968.7.10.2.b (slide); one dissected parth. ♀ from the type locality, coll. 24 February 1968 by L. Davies, NHM 2004.2310 (six slides); one parth. ♀ from locality ‘‘ Net. Riv. du camp’’, Île de la Possession, coll. 16 January 1968 by L. Davies, NHM 1968.7.10.3 (tube); 20 parth. ♀♀ from unknown locality in Île de l’Est, coll. 28 February 1969, NHM 1970.4.30.1 (tube) .
Other material studied
Marion Island: six parthenogenetic females from loc. 2494, coll. 1965 by B. J. Huntley (Marion Island Expedition 1965–1966), tube NHM 1971.6.8.96–103.; one parth. ♀ from loc. 3345, coll by E. Z. Bakker, slide NHM 1971.6.8.85.; 16 parth. ♀♀ from loc. 3346, coll. by Marion Island Expedition 1965–1966, tube NHM 1971.6.8.55–95.
New Amsterdam Island (Île Amsterdam): 119 parthenogenetic females from unknown locality, coll. 27 March 1903 by Deutch Südpolar-Expedition, tube ZMHU 17465 View Materials ; one parth. ♀ from unknown locality, coll. 27 March 1903 by Deutch Südpolar-Expedition, slide in ZMHU, no number accessed .
Diagnosis (based exclusively on Crozet specimens)
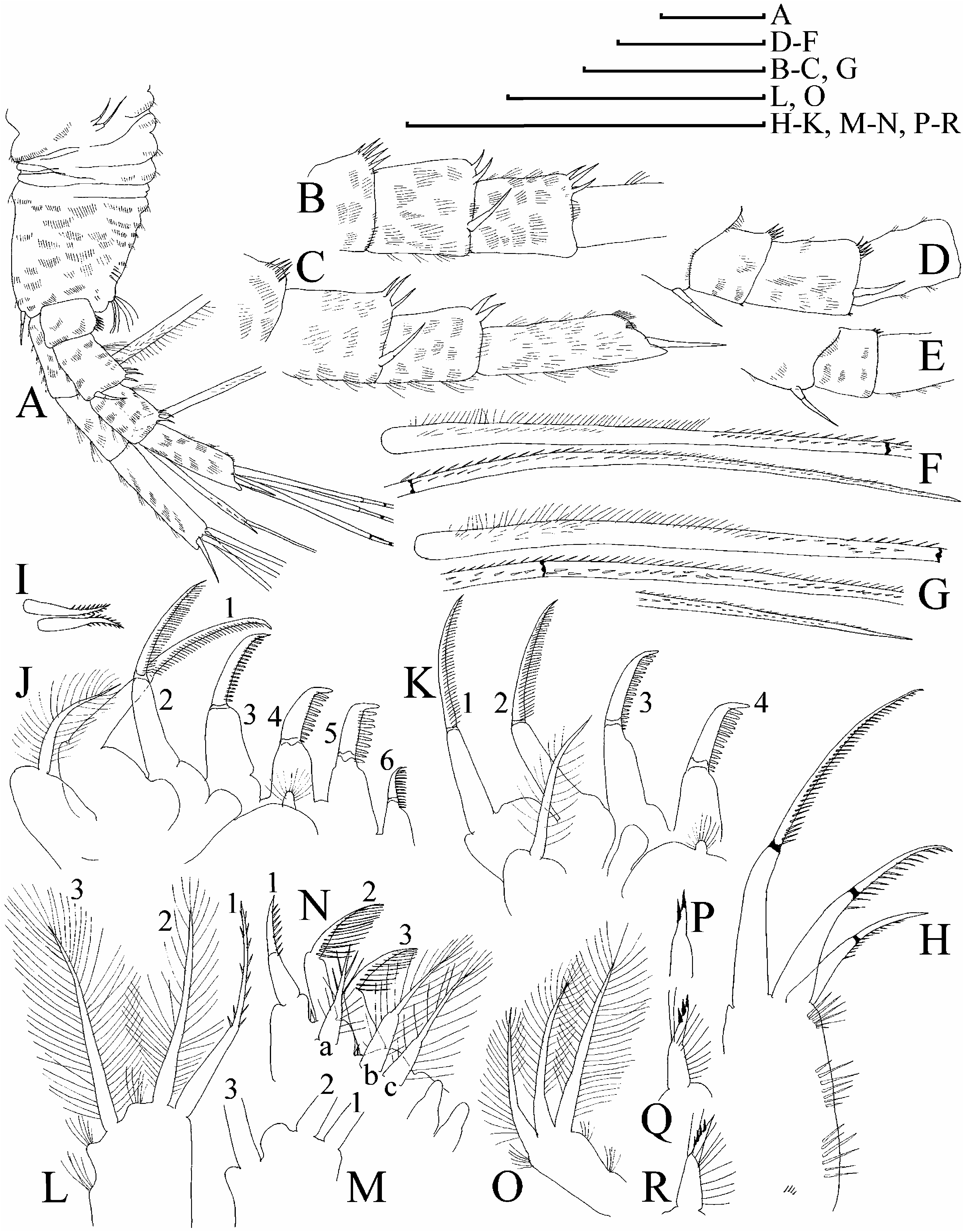
Parthenogenetic female. In lateral view body subovoid (height/length50.62– 0.67 in large adults), cervical depression present, dorsal margin breached by a ‘‘step’’ in posterior boundary of head, forming a blunt tooth, dorsal margin of valves not elevated under dorsal margin of head, without any serration ( Figure 17A View Figure 17 ). Postero-dorsal angle as rounded triangle, lies in level of middle of body height. Body compressed laterally, with a low, but sharp dorsal keel ( Figure 17B View Figure 17 ). No dome above eye; ocellus small ( Figure 17D View Figure 17 ). Dorsal organ ovoid, small. Labrum with a thick, rounded, moderately projected apex lacking any tubercles ( Figure 17D, E View Figure 17 ). Armature of ventral margin as in other species ( Figure 17F, G View Figure 17 ).
Postabdomen without ‘‘heel’’ basally, and without a reticulation on sides. Ventral margin straight, with few series of small denticles. Postabdominal seta with a short distal segment, armed with relatively short setules; basal segment without any setules ( Figure 17L, M View Figure 17 ). On external side of postabdominal claw, a series of six to nine moderately robust denticles; medial row of about seven to nine denticles; inner row with numerous denticles, organized in two successive series ( Figure 17H, I View Figure 17 ), on inner side two pectens of denticles ( Figure 17K View Figure 17 ).
Antenna I widened distally, straight or slightly curved, without a subapical external angulation; sensory seta at distance of about 1.5–2 antennular diameters (at base) from antenna I joint; on anterior face about five to seven transverse rows of spinules, but no reticulation ( Figure 17N, O View Figure 17 ). Nine short aesthetascs, two or three of them significantly larger than the rest. Antenna II with distal burrowing spine on basal segment as long as or longer than proximal segment of exopod ( Figure 18A View Figure 18 ). Length of all apical swimming setae subequal, their armature as in M. boergeni . Lateral seta on proximal endopod segment larger than other setae, lacking robust denticles in middle ( Figure 18G View Figure 18 ). A spine on second segment of exopod half as long as next segment ( Figure 18B, C View Figure 18 ), on posterior side of segments 1–3 of exopod there are series of relatively large additional denticles.
Limb I outer distal lobe with longest apical seta having distal segment unilaterally armed with robust setules; inner-distal lobe with three bisegmented setae of different size, unilaterally setulated in distal part, smallest one with whole distal segment setulated, or these setules present only in proximal portion of distal segment ( Figure 18H View Figure 18 ); two ejector hooks of similar size ( Figure 18I View Figure 18 ). On limb II, scrapers 1–2 with delicate feathering, scrapers 3–7 with robust denticles of size characteristic for the genus ( Figure 18J, K View Figure 18 ); a solitary posterior seta present near gnathobase; filter plate II with four setae, without a rudiment of fifth seta. On limb III epipodite with five setae; a distal group of three long setae, seta 1 armed with robust denticles ( Figure 18L View Figure 18 ); setulated projections proximally to seta 3 and between setae 2 and 3; on inner-distal limb portion, seta 1 with short and robust denticles; seta a with fine setules basally and robust spinules distally, seta b only somewhat longer than c ( Figure 18N View Figure 18 ); basal endite posteriorly with four soft setae. Limb IV with exopodite small, bearing only a distal group of three bilaterally feathered setae of different size ( Figure 18O View Figure 18 ); on inner-distal portion of this limb seta 1 with strong setules basally and two to six robust denticles distally ( Figure 18P, Q View Figure 18 ); posteriorly, row of five long setae. On limb V there are three setae at inner margin.
Differences of juvenile female. In contrast to adult, body more elongated, with less-developed tooth on posterior border of head, with valve dorsal margin less convex, with antennae II and swimming antennal setae longer, rows of setules on antenna I and II weakly developed ( Figure 17C View Figure 17 ).
Ephippial female, male. Unknown.
Differences of Île Amsterdam populations. Île Amsterdam females ( Figures 17J, K, O View Figure 17 ; 18D–F, M, R View Figure 18 ) have (1) smaller, more numerous (up to 10), denticles projected exactly dorsally on outer surface of postabdominal claw; (2) antenna I somewhat more widened distally; (3) shorter aesthetascs of antenna I with smaller differences between smallest and largest members; (4) very small additional denticles on segments 1–3 of antenna II; (5) more prominent projections between setae 2 and 3 and distally to seta 3 on exopodite III; (6) more numerous denticles (up to six) on seta 1 of inner margin of limb III. Except for difference 1, the rest of the specific traits of Île Amsterdam populations seem to be subject to strong variability among Macrothrix populations. Populations from all three groups of islands examined here are considered conspecific.
Size. Holotype 0.84 mm, juvenile and adult parthenogenetic females from the type series, Crozet 0.44–0.97 mm, from Île Amsterdam 0.65–1.00 mm.
Taxonomic notes. Rühe (1914) was the first to observe M. ruehei sp. nov., which he identified as ‘‘ M. hirsuticornis ’’, although he mixed in the description populations from Île Amsterdam and Kerguelen, belonging to different species in present understanding. Brehm (1958) made a drawing of the head of ‘‘ M. hirsuricornis ’’ from Île Amsterdam with widely rounded labral apex. This is undoubtedly M. ruehei sp. nov. There is no doubt about the conspecificity of populations from Crozet, Marion, and Île Amsterdam, but due to small differences between them, all material not from Crozet is ‘‘expressly excluded from the type series’’ (case 72.4.1 of the International Code of Zoological Nomenclature ( International Commission on Zoological Nomenclature 2000)).
Distribution. French Southern Territories: Crozet, Marion, Île Amsterdam, but not Kerguelen, where its congener, M. boergeni , occurs. Apparently, M. ruehei sp. nov. is a very common species in small water bodies on the aforementioned islands. Note that it is not clear which species of Macrothrix (recorded by Dartnall (1995)) inhabits Heard Island.


| ZMHU |
Zoologisches Museum der Humboldt Universitaet |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.