
Afroedura gorongosa, Rödel, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4324.1.8 |
|
publication LSID |
lsid:zoobank.org:pub:B4Ff9A5F-94A7-4E75-9Ec8-B3C382A9128C |
|
DOI |
https://doi.org/10.5281/zenodo.6042038 |
|
persistent identifier |
https://treatment.plazi.org/id/A1F7519B-C809-41E2-B63A-9944C3E27524 |
|
taxon LSID |
lsid:zoobank.org:act:A1F7519B-C809-41E2-B63A-9944C3E27524 |
|
treatment provided by |
Plazi |
|
scientific name |
Afroedura gorongosa |
| status |
sp. nov. |
Afroedura gorongosa sp. nov.
( Figs. 4–5 View FIGURE 4 View FIGURE 5 )
Afroedura transvaalica (part) Broadley 1966, p. 111.
Holotype. ZMB 83293 (GNP 484), adult male, western flank of Mount Gorongosa , Gorongosa National Park, Sofala Province, Mozambique (1038 m a.s.l., 18˚28’04.3”S, 34˚02’53.2”E), collected by M.- O. Rödel and M.F. Barej, 11 December 2015.
Paratypes (three specimens). ZMB 83290 (GNP 438), adult female, with incision in left thigh; PEM R22220 (GNP 439, previously ZMB 83291), adult female, with incision in left thigh; ZMB 83292 (GNP 440; will later be deposited in the zoological collection of the E.O. Wilson Laboratory in Gorongosa National Park), adult male, with everted hemipenes and incision in left thigh; collected on 22 May and 21 July 2015 by J.A. Guyton and P. Naskrecki from gallery forest near Murombodzi Waterfall , Mount Gorongosa, Gorongosa National Park, Sofala Province, Mozambique (842 m a.s.l; 18°29'0.1''S, 34°2'34.6''E). GoogleMaps
Additional material (two specimens). ZMB 83288 (GNP 433), adult male, ZMB 83289 (GNP 434), adult female; both collected in April 2015 by H. Farooq, P. Naskrecki and J.A. Guyton from the top of Bunga Inselberg, Gorongosa National Park, Sofala Province, Mozambique (approx. 200 m a.s.l., 18˚35’58.1”S, 34˚20’34.8”E).
Etymology. The specific name refers to Mount Gorongosa and Gorongosa National Park in Central Mozambique, to which the species is endemic. We suggest Gorongosa Flat Gecko is a suitable common name. Afroedura , based on Oedura , is feminine and the specific epithet is treated as a noun in apposition.
Diagnosis. A member of the A. transvaalica -complex in possessing two pairs of enlarged scansors per digit and a strongly verticillate and flattened tail ( Jacobsen et al. 2014). Differs from other members of the complex by having 97–102 midbody scale rows (less than 95 in A. bogerti- complex, 113–120 (= 117) in A. loveridgei and 102–119 in A. transvaalica (102–118, = 109 in South Africa—Jacobsen et al. 2014; 108–119, = 114 in Zimbabwe adjacent to Mount Gorongosa); by the rostral bordering the nostril (nostril excluded from rostral in A. loveridgei ); by the anterior nasals being separated by 1–3 granules (always in broad contact in A. loveridgei ; usually in broad contact in A. transvaalica - <3% in South Africa populations, 18% separated by a single granule in Zimbabwe adjacent to Mount Gorongosa); in having 19–22 scales between the anterior borders of the eyes (15–19 in A. loveridgei ; 17–20 in South African A. transvaalica , and 15–19 in Zimbabwe populations adjacent to Mozambique); and in having a higher average number of precloacal pores in males (8–13, = 10; 6–10, = 8 in A. transvaalica ; 8–10, = 9 in A. loveridgei ).
Holotype description.
Morphology. Adult male; SVL 50.8 mm; tail 52.2 mm (partly regenerated), with a small incision in the left thigh for the removal of muscle tissue. Head and body dorsoventrally depressed; HL 12.3 mm, HW 9.0 mm, broadest at mid-eye and 1.37x longer than wide. Eye large (3.7 mm wide), pupil vertical with indented margins; circumorbital scales small and granular, becoming elongate, almost ciliate, at upper posterior margin. Snout rounded, 4.9 mm long, longer than distance between eye and ear openings (3.6 mm). Scales on top of snout granular, rounded and convex, the largest being 2/3rds the width of the scales on the back, which are granular and juxtaposed, with no intervening minute granules. Scales on snout almost twice as large as those on crown and throat. Scales on belly flattened, imbricate, more-or-less ovate at mid-ventrum, and twice the size of lateral granules and 1.5x those along backbone. Ear opening deep, oblique and roughly oval, only half as high as wide.
Limbs well developed, hindlimbs slightly longer than forelimbs, both without obvious mite pockets at posterior margin of limb insertions. All digits with a large pair of distal scansors, separated by a large, curved claw, and followed after a large gap (twice length of terminal scansor) by a smaller pair of scansors; infero-median row of digital scales enlarged transversely, particularly towards the scansors, where the terminal scale adjoining the first pair of scansors may be medially constricted, swollen and scansor-like; 12 scale rows under 4th toe.
Nostril pierced between rostral, 1 st supralabial and three nasal scales; the supranasal being much larger than the subequal postnasals and separated by a granule bordering the rostral, followed behind by two smaller granules. Rostral roughly rectangular but with its upper edges elongated due to extensions into the nostril. Nine supralabials on each side, the labial margin flexing dorsad at the rictus, with 1–2 minute scales proximal to the flexure. Nine infralabials on either side, with a small scale proximad to the flexure. At the lip, mental slightly narrower than adjacent infralabial, only 2/3rds the width of rostral, and in contact with two distinctly elongate postmental scales.
Approximately 20 scales across crown at level of front of eye; 13 scales between nostril and front of eye; 23 scales from front of ear to back of eye; 99 scales around midbody.
A roughly V-shaped row of 13 precloacal pores occurs eight scale rows anterior to the cloacal lip, with these scales more imbricate and triangular adjacent to the precloacal pores. Original portion of tail slightly dorsoventrally flattened and indistinctly verticillate, without obvious lateral constrictions; each verticil comprising 6–7 imbricate rows of scales dorsally and 5 imbricate scale rows ventrally, and with ventral scales approximately twice the size of those on dorsal surface. Hemipenal pouches well-developed, with an angular, ventro-lateral series of 3 enlarged tubercular scales on each side (largest nearest cloaca), and with two openings of the cloacal sacs on the posterior lip of the cloaca.
Colour in life (from photographs, Fig. 4a View FIGURE 4 and unpublished). Dark black-brown above with a series of five white dorsal spots (4–5 scales across) between the fore- and hind-limb insertions, that are surrounded by a chocolate brown zone that encompasses a series of 2–4 laterally arranged smaller pale spots (2–3 scales wide), and posteriorly by a diffuse pale region that forms an irregular band across the mid dorsum. Top of head, flanks and upper surfaces of limbs with small, scattered pale spots. Head with a dark brown band across the posterior edge of crown encompassing a small central pale spot; a vague, thin pale canthal stripe, extends on both sides from the nasal region to front of eye; upper and lower labials grey-brown with diffuse yellowish bars; iris reddish brown with a heavy dark brown reticulation, and pupil with crenulated edge. Original segment of tail mottled in dark brown and pale blotches; regenerated tail grey with vague darker blotches. Ventrum mottled grey-cream.
Colour in preservative. Dark black-brown above with irregular markings on back, comprising four equally spaced dark bars, each with a central white spot 4–5 scales wide, and an irregular, paler brown bar behind; top and sides of head, flanks and upper surfaces of limbs with small, irregular pale spots; ventral surface of throat, neck, belly, limbs and tail uniform dark grey. Side of head and labials dark brown, with a few scattered pale spots on upper labials, and vague pale edges to lower labials.
Variation in paratypes and additional material ( Figs. 4b, 4c View FIGURE 4 , 5 View FIGURE 5 ; Tables 3–4).
Morphology. SVL from 54.6 mm ( ZMB 83289) to 61.2 mm ( PEM 22220); head length 1.24–1.38 times head width (max. in paratypes 1.38); snout only 1.5 times diameter of eye in one specimen. The supranasals always separated by granules, usually with a single large granule in contact with the rostral between the supranasals, followed by 2–3 smaller granules in lateral contact; the first upper labial and rostral always enters the nostril, and the width of the rostral at the lip margin is always wider than that of the mental (except in ZMB 83290 were they are subequal); always two postmental scales; supralabials more than in holotype (10–11), infralabials (9–10); scales between anterior edge of eye 19–21; scales between nostril and anterior edge of orbit 12–13; scales between anterior edge of ear and rear margin of orbit 21–24; scales around mid-body 97–101; subdigital lamella beneath 4th toe 10–11; dorsal scales per tail verticel 7–8; ventral scales per tail verticel 5.
In one female ( ZMB 83290) the terminal, medially-constricted subdigital lamella of the 3rd toe of left foot is swollen and scansor-like, bearing small setae. Precloacal pores 8–13 in males, females without dimples in precloacal scales (as in A. transvaalica , but unlike A. loveridgei where five females had 5–9 dimpled precloacal scales, that lacked secretory activity, and only one had none).
Colour in life (based on female paratype PEM R22229). Boldly patterned, with six dorsal dark transverse bands, posteriorly edged with cream, the first on the neck, the second and sixth at the fore- and hindlimb insertions, respectively, and with three thin bands equally spaced between them on the body; background colour of body, head and upper surfaces of limbs flecked on light brown and cream with occasional pale spots 2–4 scales wide; a dark canthal stripe runs through the rostral to the front of the eye, with a paler zone above; the scales of the anteriordorsal region of the orbital rim are bright yellow, with a faint yellowish infusion of the crown scales between the eyes; the original tail is boldly barred dorsally with seven, subequally-spaced, dark brown-black bars with a yellow posterior edge; belly uniform cream with scattered yellowish scales, greyer on ventral surfaces of limbs and tail.
Colour in preservative. Similar to holotype, but dorsum pale brown to grey; larger dorsal markings usually reticulated, occasionally forming irregular transverse bands (6–7 from occiput to between hindlimbs); smaller dorsal markings vary from vague to distinct. A few specimens have irregular pale blotches along the posterior edges of the darker dorsal patches/bands, and also scattered on the dorsal surfaces of the limbs and back. Dorsal surface of regenerated tails usually with irregular pale and dark blotches that may form wavy bands, and which become flecks or grey blotches below.
Size. Largest male (ZMB 83288)—SVL 56.5 mm, tail (partially regenerated) 55.7 mm; largest female (PEM R22220)—SVL 61.2 mm, tail (original) 64.0 mm. It is thus intermediate in size between A. transvaalica (malemax. SVL 72 mm, female—max. SVL 64 mm) and A. loveridgei (male—max. SVL 59 mm, female—max. SVL 58 mm). Only one of six specimens had an original tail (105% SVL).
Hemipenis (based on ZMB 83292). Approximately 5 mm long, with a simple, smooth, possibly longitudinally flounced pedicel; capped with two large cups covered with fine calyculate ornamentation; sulcus simple and draining into the conjoined base of the distal cups.
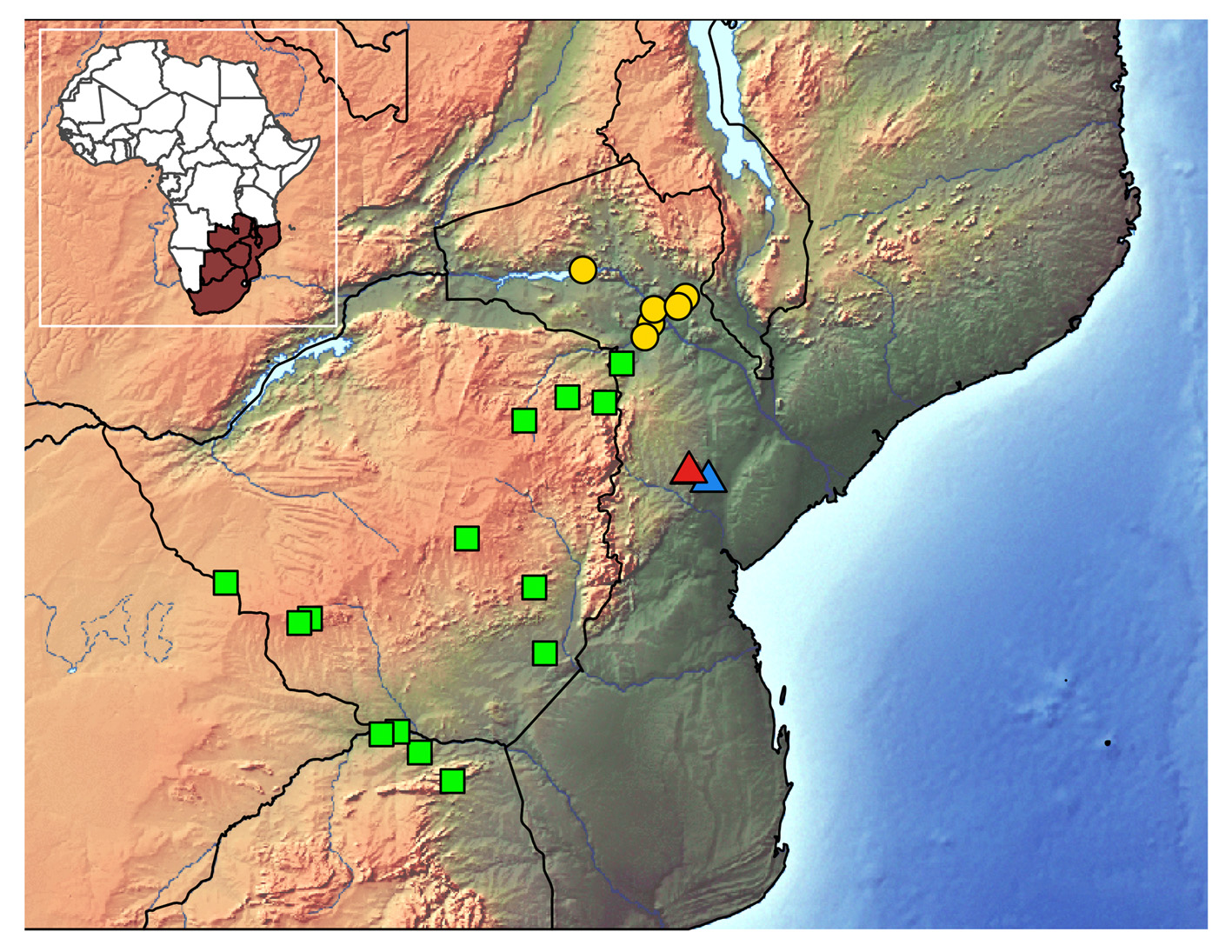
Distribution. Known from only three localities in the Mount Gorongosa region ( Fig. 2 View FIGURE 2 ). The holotype and paratypes, plus various non-collected individuals, have been found on the western flanks of Mount Gorongosa between about 900 and 1100 m a.s.l. ( Figs. 6a–c View FIGURE 6 ). Access to further potential areas on the mountain, i.e. rock faces at the eastern slopes, was not possible due to security concerns. The additional material and various non-collected specimens have been observed at a lower altitude on the Bunga Inselberg (about 212 m a.s.l.) in the north-western corner of the National Park, approximately 40 km from the type locality ( Fig. 6d View FIGURE 6 ). No tissue samples from the Bunga individuals were available, and the assignment of these individuals to the new species was thus based on morphological similarity.
Natural history. The Gorongosa National Park is situated in Central Mozambique approx. 100 km south-west of the Zambezi River, and is in the southern part of the great African Rift Valley. Mount Gorongosa is an isolated granite mountain massif rising to 1863 m a.s.l., north-west of the National Park’s plains. At its highest elevations the massif’s plateau is covered with montane grasslands and forest patches dominated by cypresses ( Widdringtonia nodiflora ); the upper slopes of the mountain (900–1600 m) support a belt of moist evergreen forest. The type series was collected in an area between 900 and 1100 m on the western flanks of the mountain ( Figs. 6a–c View FIGURE 6 , 7 View FIGURE 7 ). The paratypes were collected in a small alcove beneath a boulder pile in the closed-canopy riverine forest beside the Murombodzi Waterfall, and thus in comparatively cool and moist microclimates ( Figs. 6c View FIGURE 6 , 7 View FIGURE 7 ). Some rocks with lizards present were located within the splash zone of the waterfall. Other specimens were observed but not collected at this site. The holotype was collected from a deep narrow crack in a large granite rock (i.e. 8 m ³) in highly degraded grassland below the edge of the rainforest and almost fully exposed to the sun ( Fig. 6b View FIGURE 6 ). It was the only specimen seen at that site. At both sites the new species lived in syntopy with the cordylid Smaug mossambicus (FitzSimons, 1958) .
The additional non-type material comes from the Bunga Inselberg ( Fig. 6d View FIGURE 6 ), a partly eroded inselberg composed mostly of trachyte in the north-western corner of the national park, covered and surrounded by dense Miombo woodland at a relatively low altitude (approx. 212 m a.s.l.). Here the geckos were observed during the day and at night in several places in deep cracks between very large boulders. No geckos were observed outside of these cracks. Syntopic lizards were Smaug mossambicus , the skink Trachylepis margaritifera (Peters, 1854) and the gecko Hemidactylus platycephalus Peters, 1854 . Further sympatric, but not syntopic geckos were: Lygodactylus cf. capensis (Smith, 1849) , Pachydactylus punctatus Peters, 1854 and Chondrodactylus turneri (Gray, 1864) .
The new species was observed regularly between April and December 2015. Like A. transvaalica , it is tolerant of conspecifics and frequently several specimens, and sometimes as many as over 20, were observed sheltering in deep rock crevices. One adult female (ZMB 83290) contained two eggs of approximately 11.5 x 7.5 mm.
Conservation Status. The population of the new gecko present on the lower slopes of Mount Gorongosa is threatened by illegal deforestation taking place within the riverine forest adjacent to Murombodzi Waterfall. Slashand-burn removal of tall, old growth trees around the rocks may cause changes to the humidity and water availability in these currently very moist, shaded microhabitats that seem to be preferred by the lizards. Reforestation efforts led by Gorongosa National Park and introduction of shade-grown coffee as an alternative to slash-and-burn agriculture still practiced around the mountain will hopefully slow or halt the loss of the native riverine and evergreen vegetation in that area. On Bunga Inselberg the population of A. gorongosa is currently protected within the core area of the National Park and no immediate threat to its survival exists, though deforestation for agriculture is slowly encroaching on the area. However, based on the existence of only a few known sites and small area of occupancy, i.e. the presumed restriction to a fragmented range on Mount Gorongosa and potentially some rocky areas surrounding the mountain, as well as the ongoing high rate of habitat degradation and conversion on the mountain, the species may be considered highly threatened, potentially justifying an assessment as Endangered or even Critically Endangered. Further surveys and formal IUCN Red List assessment are required to confirm this.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.