
Allosorex stenodus FEJFAR , 1966
|
publication ID |
https://doi.org/10.37520/fi.2020.006 |
|
persistent identifier |
https://treatment.plazi.org/id/03E3CA45-FFAA-8620-D72C-FDADFEA51A81 |
|
treatment provided by |
Diego |
|
scientific name |
Allosorex stenodus FEJFAR , 1966 |
| status |
|
Allosorex stenodus FEJFAR, 1966
Text-figs 2–7 View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig , 9 View Text-fig , 10 View Text-fig
1966 Allosorex stenodus nov. gen., nov. sp.; Fejfar, pp. 227–237, Taf. 23–27, Abb. 1–9.
1985 Allosorex stenodus ; Fejfar and Heinrich, p. 220.
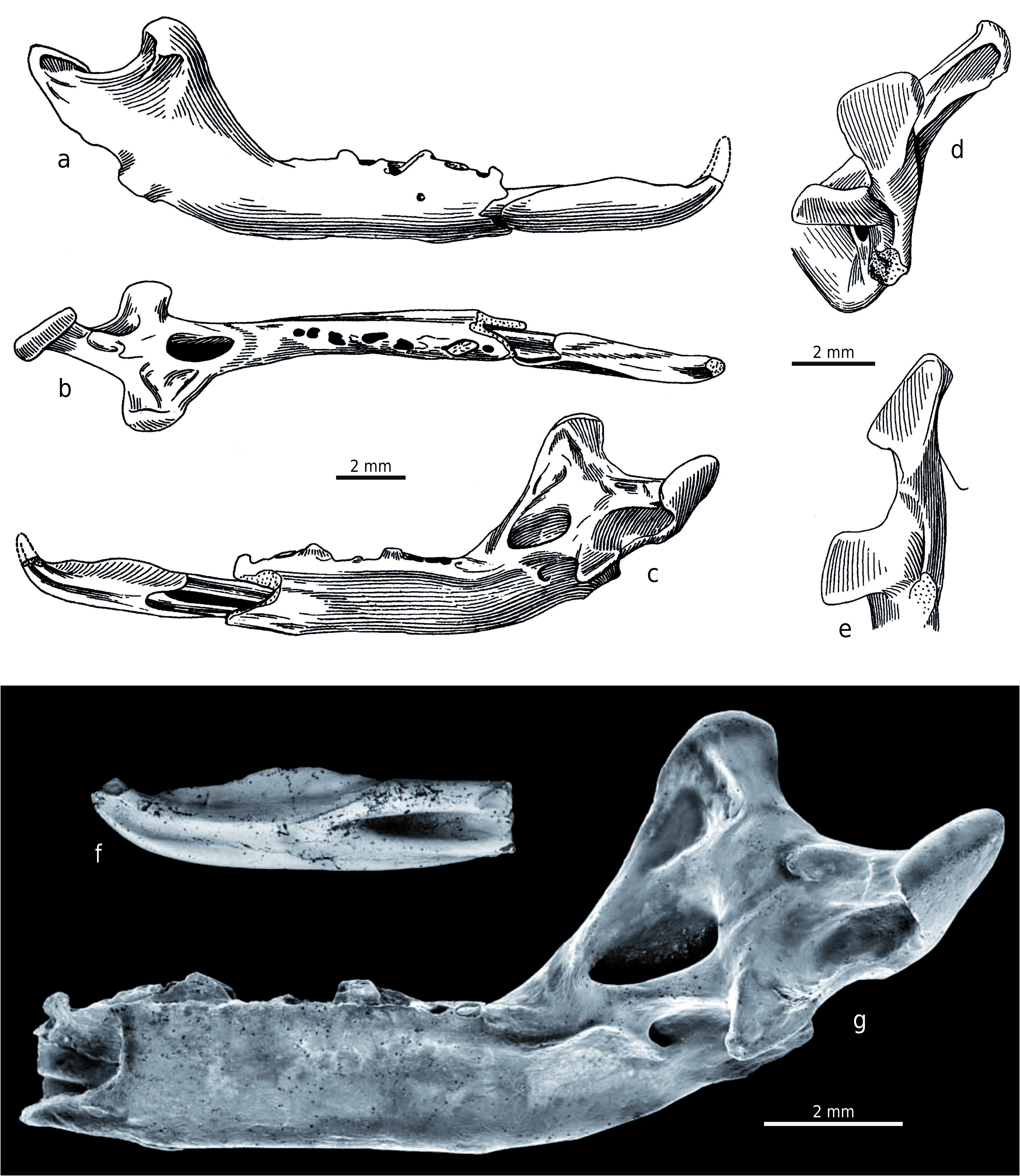
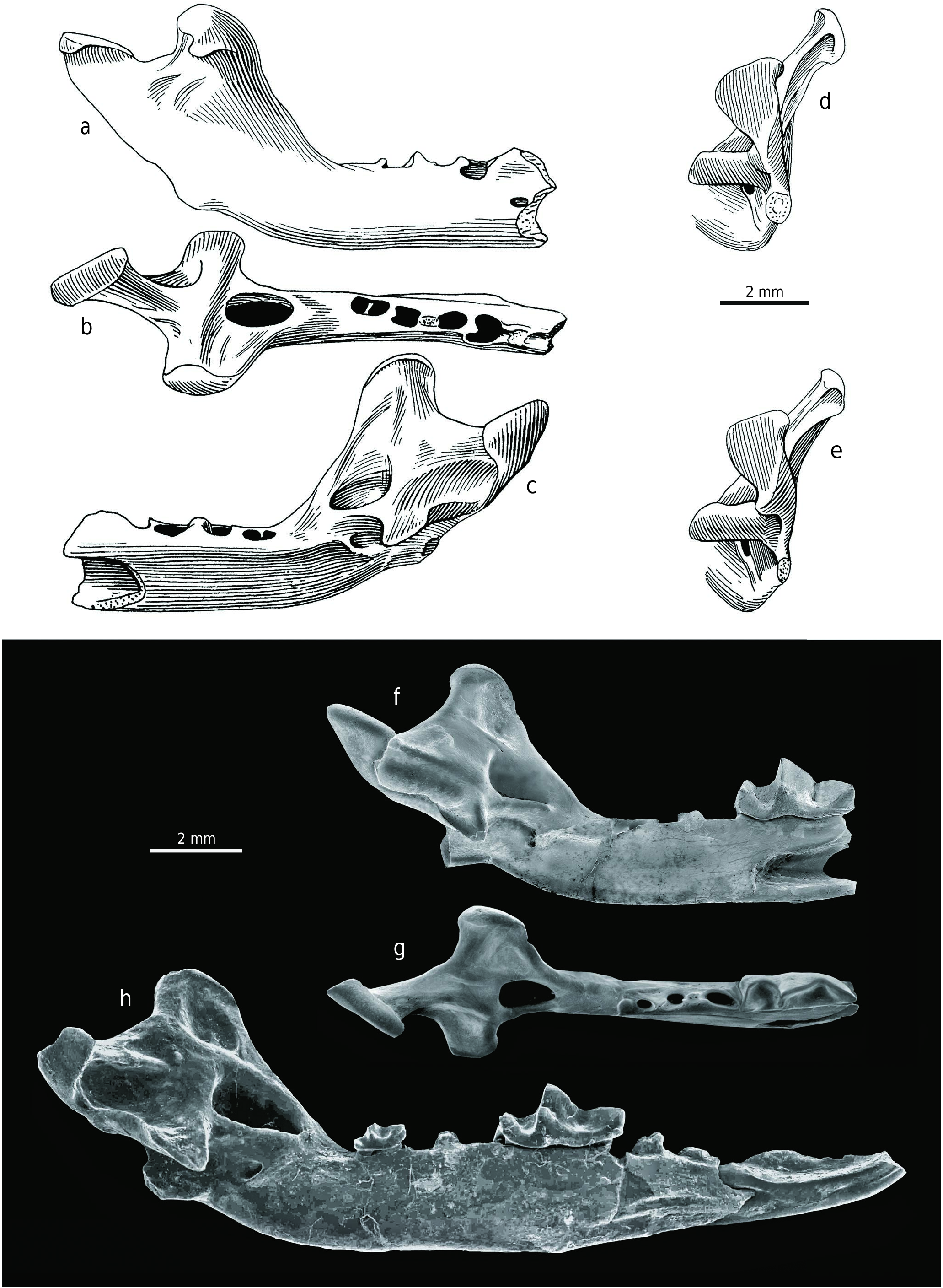
H o l o t y p e. Right mandible with the incisor and alveoli for a2–m3, without the angular process (Z 28208 – OF 651273 View Materials ; Fejfar 1966: Abb. 1, Taf. 23.1–3, Taf. 24).
P a r a t y p e. Incomplete right mandible with antemolar (currently absent), m1 and m2, and alveoli for m3, without the angular process (Z 28209 – OF 652356 View Materials ; Fejfar 1966: Abb. 2, Taf. 23.4, Taf. 25, Taf. 26.5).
T y p e l o c a l i t y. Ivanovce, fissures 6512, Slovakia
(early Pliocene, late Ruscinian, MN 15).
E m e n d e d d i a g n o s i s. Large soricid (Lm1 = 2.59–2.88 mm) with dental formula 1.?.3./1.2.3. The fissident I1 has a curved cutting edge on the talon; the P4 parastyle with its high and sharp paracone is larger than the protocone; M3 is short and wide. The acuspulate to bicuspulate hookshaped i1 has a distinct lingual groove; a2 is single-rooted; low molars lack the entoconid, the long paralophid forms a distinct sharp cutting edge (except in m3) and the low metaconid is situated close to the high protoconid; the m3 talonid is distinctly reduced. The molar enamel exhibits the Soricella -schmelzmuster. The coronoid process is low; the interarticular area is very short to almost absent; the triangular to rectangular upper condylar facet is positioned caudally and the lower condylar facet is strongly displaced forward (see also Fejfar 1966: 224–226 and Reumer 1984: 18).
S t r a t i g r a p h i c a l d i s t r i b u t i o n. Early Pliocene, late Ruscinian (MN 15).
G e o g r a p h i c a l d i s t r i b u t i o n. Central Europe (probably an endemic form, which so far is known maybe only from Ivanovce locality).
S i t e. Ivanovce, vertical fissures 6510 and 6520, and horizontal fissures 6512 and 6523 ( paratype locality) (early Pliocene, late Ruscinian, MN 15 ), Slovakia .
M a t e r i a l. I dext. fragment (Z 28191 – OF 6512 ) ; I dext. (Z 28192 – OF 6523 ) ; I sin. (Z 28193 – OF 652360 View Materials ) ; 2 fragmented right upper incisors and 2 fragmented left upper incisors (Z 28194/1–4 – OF 6523 ) ; 2 fragmented i dext. (Z 28195 – OF 652361 View Materials , Z 28196 – OF 652362 View Materials ) ; i dext. fragment (Z 28197 – OF 652363 View Materials ) ; i sin. fragment (Z 28198 – OF 6510 ) ; i sin. without the root ( Z 28199 View Materials – OF 6523 ) ; i sin. fragment (Z 28200 – OF 6523 ) ; P4 sin. (Z 28201 – OF 6523 ) ; M3 sin. (Z 28202 – OF 6523 ) ; 3 m 1 dext. (Z 28203 – OF 652364 View Materials , Z 28204 and Z 28205 – OF 6523 ) ; m2 dext. (Z 28206 – OF 652 ) ; m2 sin. fragment (Z 28207 – OF 6512 ) ; right mandible with incisor and alveoli of a2 to m3 (Z 28208 – OF 651273 View Materials , holotype) ; incomplete right mandible with m1 and m2 (antemolar is lost) and alveoli of m3 (Z 28209 – OF 652356 View Materials , paratype) ; incomplete toothless right mandible with alveoli of m1 to m3 (Z 28210 – OF 652358 View Materials ) ; 2 right mandible fragments (Z 28211 – OF 652357 View Materials , Z 28212 – OF 6523 ) ; left mandible with the incisor, m1 and m3 (Z 28213 – OF 6523 ) ; fragment of left mandible with the fragmented incisor, antemolar (a2) and m2 (Z 28214 – OF 6523 ) ; left mandible with m1 (Z 28215 – OF 652366 View Materials ) ; toothless left mandible (Z 28216 – OF 6523 ) ; 3 left mandible fragments (Z 28217 – OF 651274 View Materials with alveoli of m2 and m3; Z 28218 – OF 6520; Z 28219 – OF 6523 ) ; and fragment of humerus dext. (Z 28220 – OF 6523 ) .
The entire fossil record consists minimally of 7 individuals (MNI). The fragment of left m2 (Z 28207 – OF 6512) was used for the enamel microstructure analysis (W. von Koenigswald). The left m1 ( OF 652367 View Materials ) previously mentioned in this collection ( Fejfar 1966: 231–232, Abb. 7a–e, Taf. 26.3–4) is currently absent.
The teeth crowns are not pigmented, fossilized in white to yellowish-white. The fossilization of mandibles is brownish-white, with black manganese plaques.
D e s c r i p t i o n o f t h e h o l o t y p e. The damaged right mandible, with the incisor and alveoli of a2 to m3 (Z 28208 – OF 651273 View Materials ; Text-fig. 2 View Text-fig ), is approximately as large as the mandible of large soricids such as Beremendia fissidens (PETÉNYI, 1864) . The incisor portion of the mandible is broken off and the horizontal ramus is slender. The small mental foramen is situated under the m1 middle portion and the mandibular foramen opens below the central part of the long and broad internal temporal fossa, which lies just above the level of the dental row. The fossa is oval shaped, very deep and distinctly bordered in its upper (dorsal) and lower (ventral) parts. The shallowly concave medial surface from the upper edge of this fossa to the wide top of the coronoid process has several irregular ridge-like tuberosities in its anterior half for the attachment of the temporal muscles. The concavity of the coronoid medial surface appears as a dorsal continuation of the internal temporal fossa and the upper edge of this fossa is thus homologous with the “limula” of other soricid types. The external pterygoid fossa is situated at the level of the internal temporal fossa and it has a completely unusual position between this fossa and the condyle. Its posterior boundary forms the lingual boundary of the upper condylar facet, and the fossa is also the deepest at this site. It gradually becomes more shallow anteriorly and slopes towards the back edge of the internal temporal fossa. Finally, the imprecisely-bounded external pterygoid fossa is generally deep and long. This ventrally borders the dorsal edge of the lower condyle and dorsally limits the lingual wall of the upper condyle process. This peculiar morphology has no similar analogy in other soricids. The external temporal fossa is a shallow depression, and this is also clearly visible in the paratype. The fossa lies on the lateral side between the broad coronoid process and the upper condylar facet, under the upper sigmoid notch. The coronoid process is relatively low, superiorly shortened and lies in a blunt 121° angle just behind the m3 alveolus. Its long base is the broadest at the upper level of the condyle and it is caudally and strongly laterally inclined, similar to the carnivore coronoid process. Its anterior edge is slightly concave and the posterior edge is very short, forming the anterior arm of the upper sigmoid notch. The coronoid process head is long, wide, and rounded with a distinct muscular bar and a caudally-oriented lateral coronoid spicule. Both mandible articulation facets on the condyle appear strange and these are very specialised being completely separated far from each other in vertical and horizontal directions; they are not joined to a lamina and there is no interarticular area for connection. The distinct and unusually thick triangular upper condylar facet is more caudally shifted. The lower condylar facet is strongly displaced forward and its medial portion is situated below the posterior margin of the internal temporal fossa. The relative height of the upper condylar facet is visible in caudal view, together with the lateral inclination of the coronoid process. The angular process is broken off and its base is laterally flattened, but the process was most likely medially-hooked.
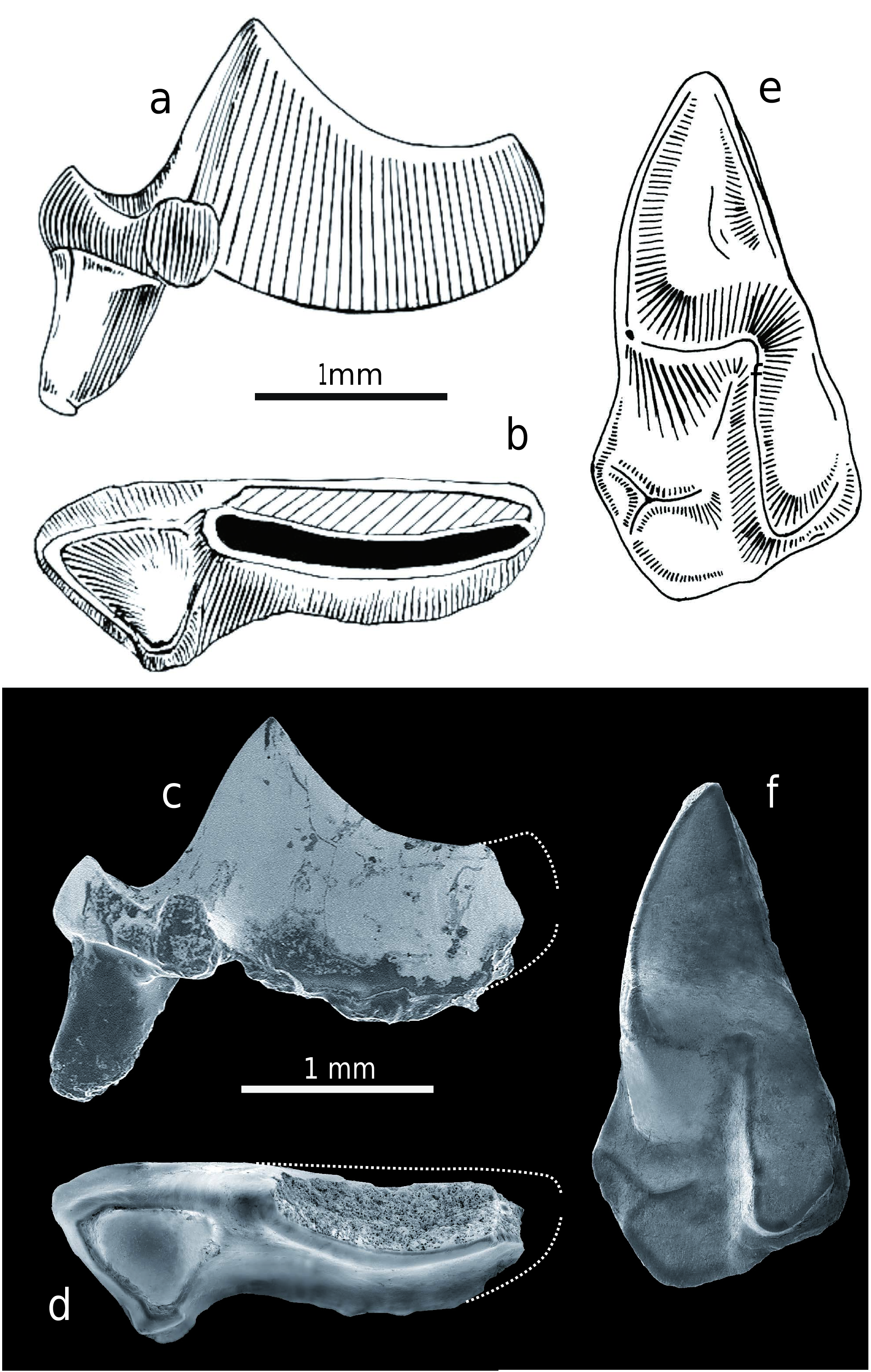
The crown of the narrow lower incisor (i1) is hookshaped, with its upwardly curved apex broken off. It has a distinct longitudinal groove on the symphysial portion (a blood-groove?) and a sharp dorso-labial edge with an irregularly, weakly waved (bicuspulate-like) dorsal margin. The symphysial wall of this cutting edge is covered with very thin enamel and the crown buccal wall has short parallel vertical wrinkles. The crown length is 6.45 mm.
Based on the preserved alveoli, the a2 antemolar was only single-rooted. The well-preserved alveoli of the double- rooted molars indicate that especially the m1 and m2 were very weakly anchored in the mandible, and this is certainly related to their sectorial function of being mechanically unaffected by chewing processes.
D e s c r i p t i o n o f t h e p a r a t y p e. The incomplete right mandible with antemolar (currently lost), m1 and m2, and alveoli for m3 and without the angular process (Z 28209 – OF 652356 View Materials ; Text-fig. 3 View Text-fig ) is approximately as large as the holotype. The incisor portion of the mandible is broken off, together with the anterior part of the slender horizontal ramus, which is also damaged below the m1 (currently separated from the mandible with a piece of its horizontal ramus). Although the mental foramen and angular process are unpreserved, the overall mandibular morphology is very similar to that of the holotype and differs only in the following details: the rounded fossa is broader; a small circular opening is situated in the posterior portion of the external pterygoid fossa; the coronoid process lies in a less blunt angle of 115° and its anterior edge is straight; the coronoid process head is not so long and the caudally oriented postero-lateral coronoid spicule is more robust; and finally the upper condylar facet is smaller than the holotype facet and the lower one is larger.
The separated first lower molar (m1) is unusually slender in occlusal view and damaged in the postero-lingual portion. The typical soricid V-shaped trigonid basin is completely expanded here, and the long paralophid is transformed into a carnassial-like blade similar to that in predatory carnivores, and it has a shallow notch. The trigonid buccal wall is very steep and its lingual slope is only moderately inclined. The U-shaped talonid basin is slightly more open lingually than seen in the second lower molar. The oblique crest resembles the paralophid and begins at the posterior wall of the protoconid. Only part of the hypolophid is preserved because the whole postero-lingual crown portion is broken off. The protoconid is the highest cusp and the others are approximately the same height, with only the metaconid protruding slightly above them when unworn. This conelike cusp, however, is the smallest of all trigonid cusps and is situated close to the protoconid. The cingulum is only indicated, or slightly developed, on the lingual side and also on the antero-buccal side under the paraconid. The anterior root is longer than the damaged posterior root and is more laterally flattened. Finally, the larger molar width lies at the distal part of the trigonid.
The second lower molar (m2) is similar in morphology to m1, but it is smaller. The trigonid basin is expanded, but not as much as in m1, and it is wide open lingually; the U-shaped talonid basin is also lingually open because no entoconid crest is developed. The oblique crest begins directly under the protoconid and passes through the low hypoconid into the straight hypolophid which ends in the distinct cone-like entostylid. The latter parallels the missing entoconid in both position and function. The protoconid is high and comparatively higher than in m1. The cone-like metaconid is situated close to the protoconid and although it is low, it is higher than the paraconid and the entostylid. The long paralophid with shallow notch resembles the carnassial cutting edge, but not as distinctly as in m1. The cingulum is only indicated on the lingual edge of the crown base and slightly developed on the antero-buccal side under the paraconid. Finally, the largest molar width is at the trigonid.
No mesoconid is developed on the preserved lower molars. Based on preserved alveoli, the third lower molar (m3) was double-rooted, with the anterior root larger than the posterior one.
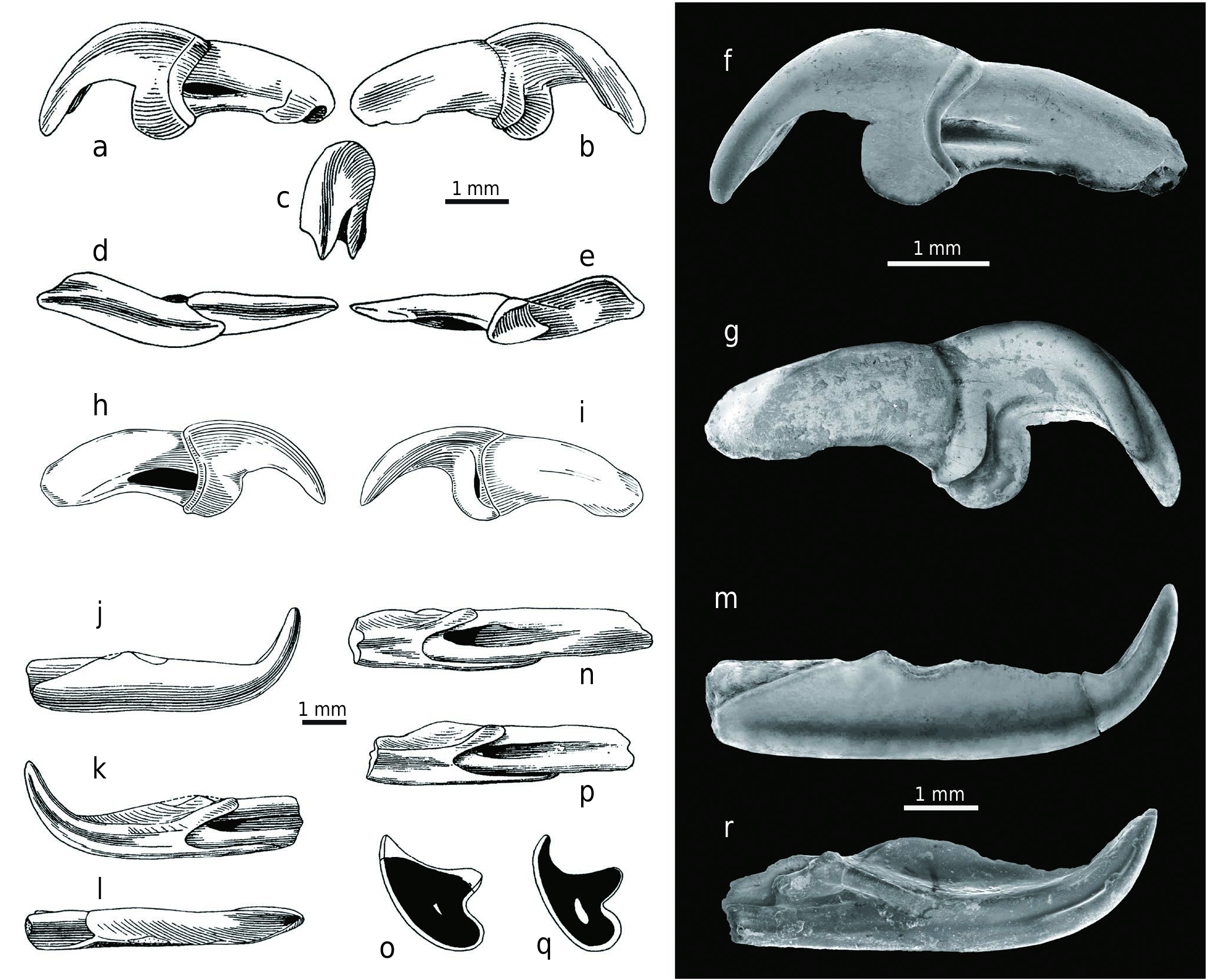
D e s c r i p t i o n o f d e n t i t i o n. Twenty nine additional teeth of Allosorex , including upper and lower incisors, upper fourth premolar, upper third molar, lower antemolar, and lower molars are available ( Text-figs 4–6 View Text-fig View Text-fig View Text-fig , Tabs 1–3). The upper incisors (I sup.) in the explored sample comprise only two complete teeth and five fragmented specimens. The narrow apex is extremely sickle-like and transversely widened in shovel-like shape in ventral view. The teeth are not fissident, and instead of the usual soricid talon with a lateral tip, there is a strange blade developed with a semi-circular cutting edge which connects to the apical posterior edge. The cingulum on the crown base under the talon is less robust on the buccal side than on the medial one where a similar structure to the talon blade can be formed (as in Z 28192 – OF 6523). The laterally flattened root has a short longitudinal groove on the buccal side and a small opening for a blood vessel at the distal end.
Upper antemolars (A1–A3) are unknown in the fossil record of the species from this site.
The preserved portion of P4 sin. (Z 28201 – OF 6523) has a possible hypercarnivorous condition, not recognised in other soricids. Its long buccal edge makes almost a 45° angle with the anterior edge. The paracone is very high and narrow, laterally flattened cusp, with the long sharp posterior cutting edge, slightly curved medially, and most likely running into a metastyle crest. The metacone/metastyle part of the crown is unfortunately broken off, and while the buccal wall of the relatively high cutting blade is heavily worn (or damaged?), the inner wall is smooth with a worn narrow stripe along the cutting edge. This resembles the similar phenomenon in the lower carnassials of large felids or hyenids. The triangular parastyle is somewhat larger than the conical, lingually situated protocone and it is connected to the paracone by a distinct parastylar crest. A crest also connects the parastyle with the protocone and another one connects the protocone with the paracone. All these three crests bounds a conspicuous anterior basin. The hypocone and the hypoconal flange are not preserved; but based on the P4 morphology of the related Viretia gracilidens species, the hypocone could be reduced or even completely absent. This is consistent with the assumed hypercarnivorous appearance of the discovered tooth fragment. The pronounced short cingulum is present on the buccal side below the parastylar crest and it passes anteriorly into the parastyle anterior crest. The anterior root is small and the length of the preserved premolar portion on the buccal side is 2.17 mm.
Only one triangular M3 sin. (Z 28203 – OF 6523) was discovered at the site. The tooth is short and wide, with the large and well-developed paracone extended by a decreasingly distinct and moderately anteriorly bent crest that runs up to the pointed (parasytyle) buccal margin. The metacone is markedly weaker, but not low, and it is connected to the paracone by a blunt crest. The antero-lingual basin is almost flat, with only an indication of a tiny blunt accessory cusp and the basin is anteriorly and lingually surrounded by a cingular ridge. Of the four roots, only the robust but shorter buccal root and the antero-lingual root are preserved, while the postero-lingual and tiny central ones are broken off. The M3 crown is 1.36 mm long and 2.69 mm wide.
The most prominent characteristic of the lower incisors (i inf.; six isolated fragments and two fragmented teeth in mandibles) is the narrow crown apex which is extremely upwardly curved. The crown has a distinct longitudinal groove on the symphysial side which is wedged almost in the middle of the crown and branches distally. A very sharp enamel edge runs from the buccal base of the apex; its dorsal margin is straight in Z 28197 – OF 652363 View Materials or relatively weakly waved as in the Z 28195 – OF 652361 View Materials ( holotype) or Z 28196 – OF 652362 View Materials . Most teeth, however, have unserrated acuspulate crowns. The distal border of the crown is posteriorly elongated on the buccal side and extends below the middle of the second lower antemolar (a2/p4). In common with other soricids, the crown has a deep indentation (sinuosity) on the symphysial side and its anterior margin terminates in the first third of the crown. A deep longitudinal groove is situated at the root’s symphysial side and the small opening for a blood vessel is visible at the distal end. Some incisors have no enamel on the crown dorsal side (see Fejfar 1966: Abb. 5e), and this lack is paralled in molars where the lingual walls of the trigonid or talonid basins have only very thin enamel or no enamel covering near the ridges .
From lower antemolars – a1 and a2 – only the latter one is preserved in the explored material and this is generally considered the p4 ( Reumer 1984). An a1 alveolus can be seen in Z 28213 – OF 6523 and Z 28214 – OF 6523 and this indicates smaller size than a2. The a2 root is inclined posteriorly (similarly like a1 root) and a2 crown is slightly hidden below the mesial margin of the m1. The a2 and m1 crown bases are almost at the same level, and this feature differs from other soricids. The a2 outline is oval in occlusal view and has a shallow fold (indentation) at the slightly wider distal margin. The only relatively high and narrow main cusp (protoconid) is clearly bi-crested – both crests are sharp and while the anterior crest runs straight anteriorly, the posterior runs posteriorly and lingually. The cusp is slightly inclined lingually, and the lateral view identifies a mesially arched anterior crest and the concave posterior crest bounding a small shallow syncline which is a homologue of the talonid basin in molars. The cingulum is well developed and absent only on the mesial edge. The buccal wall is much smoother than the lingual. The a2 from the paratype (see Fejfar 1966: Abb. 2, 6, 10a, e) is now lost and the only (faintly damaged) a2 is preserved on the fragmented left mandible with the incisor fragment and m2 (Z 28214 – OF 6523). The crown length is 1.38 mm and the width is 0.91 mm.
1
1
1
1
1/6
3
3
2
4
5
6
6
1/6
6
6
The lower molars gradually decrease from m1 to m 3 in length, width, and height and the length difference between m1 and m2 is particularly significant. While the molars’ lingual side is straight, the buccal side is slightly buccally arched. In occlusal view, their narrow shape related to the elongated trigonid is very striking compared to the normal molar shape seen in other soricids.
Three isolated right m1s and two left m1s remain in the mandibles, and these have similar morphology described in the paratype m1 above, with the difference that the crowns’ posterior portions are preserved. The hypolophid is straight and connects the low hypoconid with the high entostylid (sensu Reumer 1984), and this feature parallels the entoconid of other insectivores in both position and function. Although the entoconid is absent, a weak entoconid crest can be developed. The cingulum is slightly developed on the mesial margin under the paraconid in all m1s. The m1 from the left mandible fragment (Z 28213 – OF 6523 ) has an almost indistinguishable metaconid and indication of a small entoconid in front of the conspicuous entostylid .
The m2 morphology resembles that described in the paratype. This is especially relevant in the left m2 analysed fragment (Z 28207 – OF 6512 ) and the damaged m2 from the left mandible fragment (Z 28214 – OF 6523 ), while the isolated right m2 (Z 28206 – OF 6523 ) has no notch on the paralophid .
The only preserved and slightly damaged m3 is still situated in a left mandible (Z 28213 – OF 6523). This is a strongly reduced tooth with its length approximately half the m2 length and a third of m1. The molar is tiny and narrow, with a lingually opened and distinctly reduced and posteriorly narrowed talonid where only a small cuspid is developed (entostylid?). The protoconid is relatively robust, while the metaconid is reduced and situated very close to the protoconid. The paraconid forms only a semi-arched anterior elevation with the flat inner surface, and there is no carnassial blade-shaped paralophid developed between the paraconid and the protoconid. The weak cingulum is indicated only at the antero-buccal margin. Of the two m3 roots, only the anterior one is laterally flattened. The m3 measurements are as follows: the crown length is 1.08 mm, the trigonid width is 0.56 mm, and the talonid width is approximately 0.35 mm.
The position and shape of especially the two anterior lower molars in the mandible is certainly related to their sectorial (not-chewing) function. Moreover, this odontological specialisation correlates with the highly specialised morphological construction of the mandibles, and it is quite remarkable that the majority of the discovered mandibles are toothless, thus indicating weaker embedment in the jaws. While this phenomenon is rarely mentioned in other fossil soricids, edentulous insectivore mandibles are quite common in the fossil record.
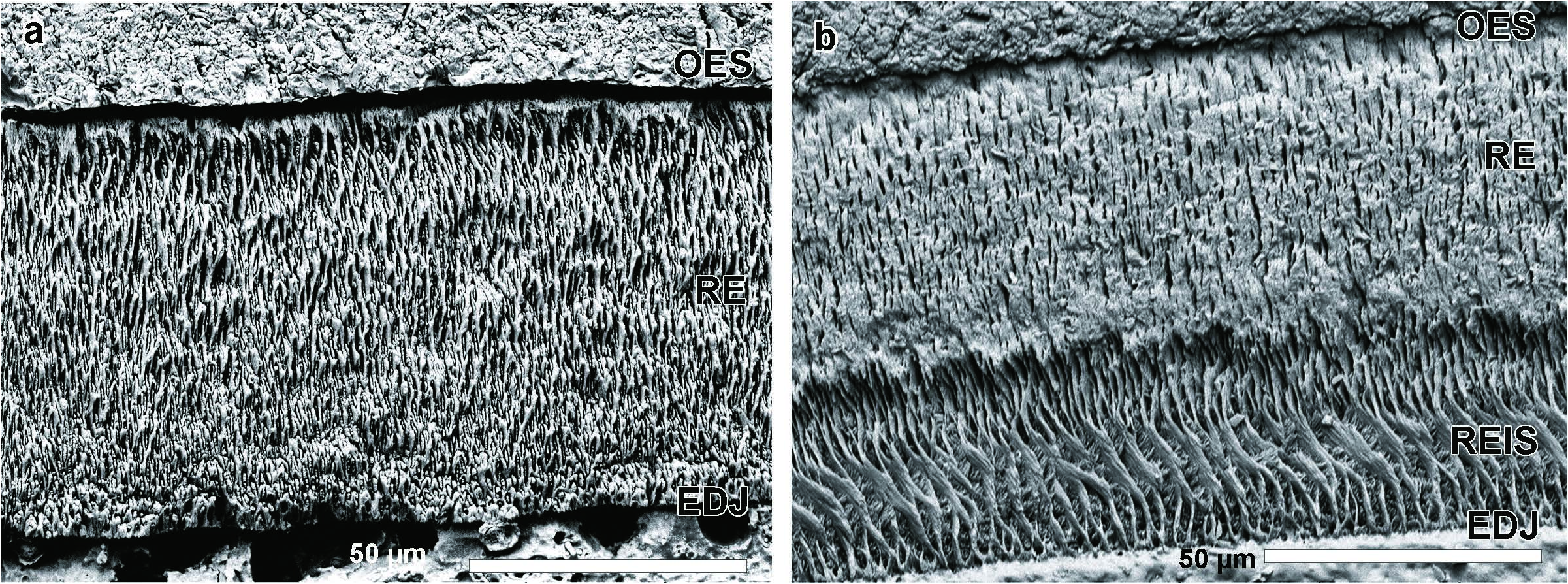
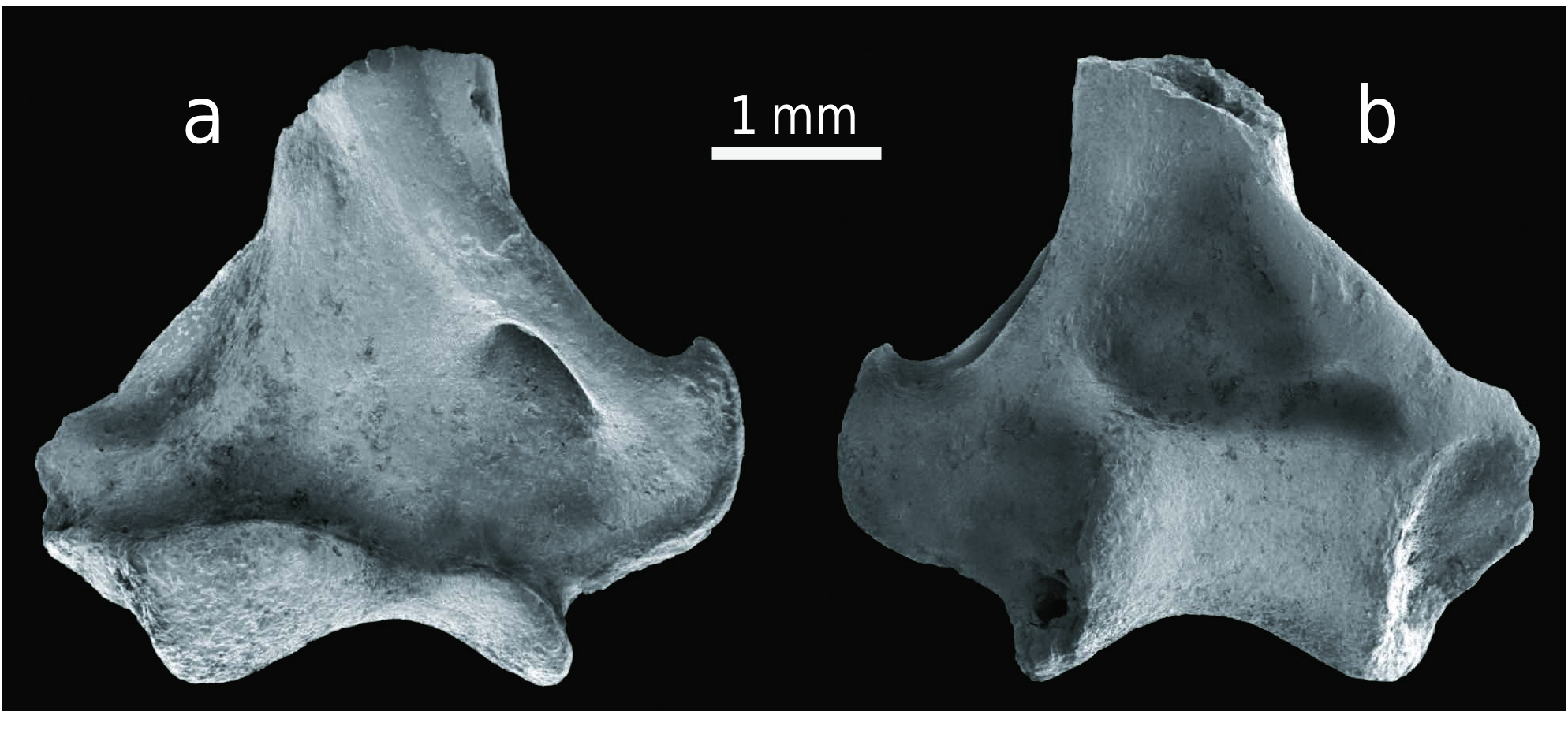
T h e s c h m e l z m u s t e r o f A l l o s o r e x. As previously mentioned, the microstructural character of paralophid enamel between the paraconid and protoconid is important in soricid differentiation ( Koenigswald and Reumer 2020). The schmelzmuster in Blarina , and similarly differentiated genera, distinctly changes along the paralophid. The paralophid enamel in Allosorex stenodus from Ivanovce is one-layered and composed solely of radial enamel. The prisms in the transverse section are seen in oblique cross-sections, as they arise from the enamel-dentinjunction (EDJ) towards the outer enamel surface (OES). The crystallites of the interprismatic matrix (IPM) are oriented at an angle to the prisms and anastomose between them. This gives the typical appearance of radial enamel. No specializations could be recognized, neither a separated inner layer with interrow sheets or a prism-less outer enamel ( Text-fig. 7a View Text-fig ). Therefore, Allosorex represents the Soricella - schmelzmuster – the least derived schmelzmuster in soricids. The simplicity of this one-layered schmelzmuster is well recognizable when compared to the two-layered schmelzmuster of Blarina brevicauda . ( Text-fig. 7b View Text-fig ).
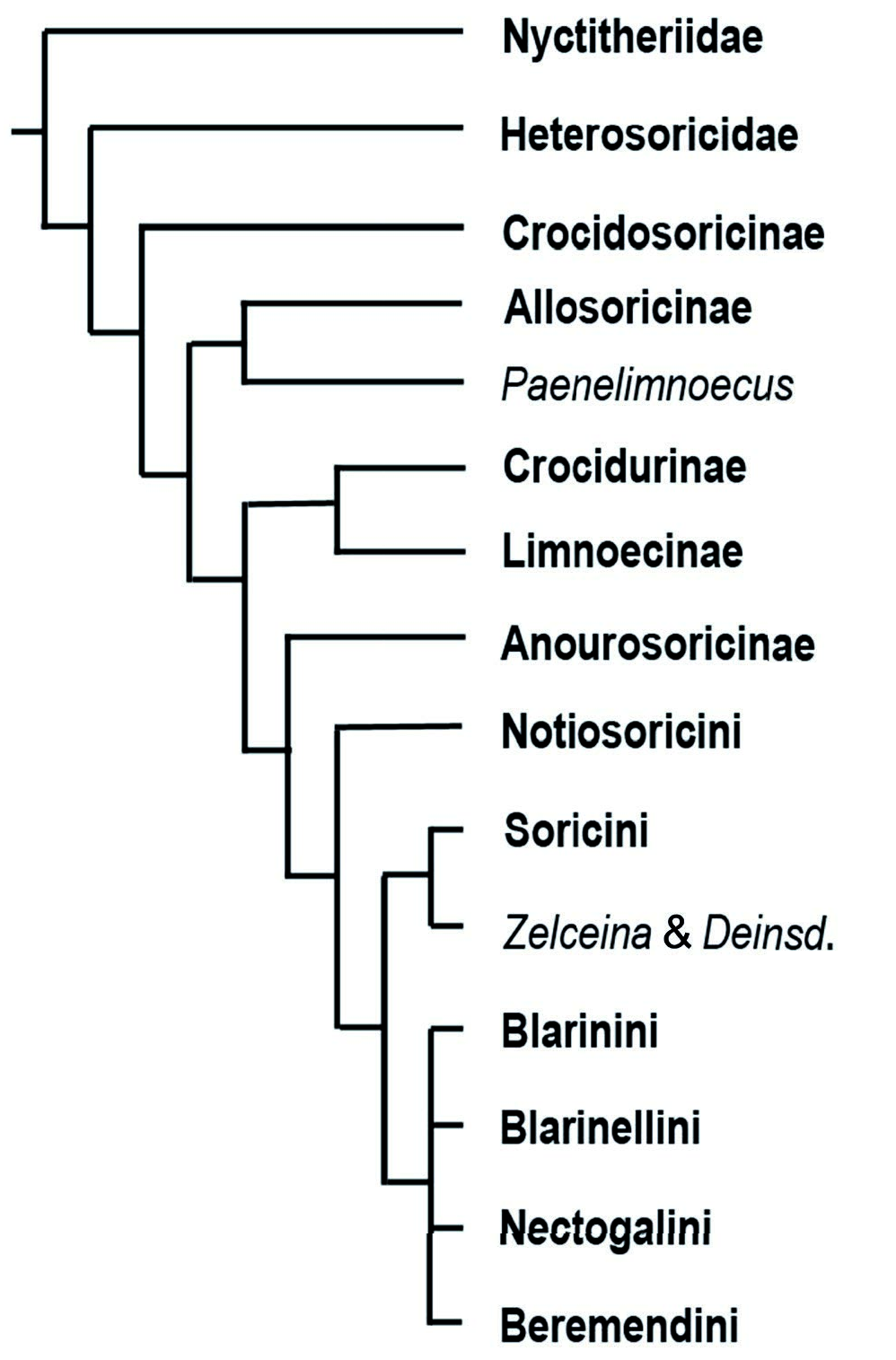
According to many investigated genera, the variability in the schmelzmuster is limited ( Koenigswald 1980, 1997, Koenigswald and Reumer 2020). Therefore, the Allosorex genus can be characterised by the basal Soricella - schmelzmuster. Furthermore, the occurrence of the Soricella -schmelzmuster can be used – in combination with other characters – to discuss Allosorex phylogenetic position. The presence of this underived Soricella -schmelzmuster excludes Allosorex from groups with a more differentiated schmelzmuster. Allosorex forms most likely an early side branch of Soricidae ( Text-fig. 8 View Text-fig ). Thus, the enamel supports the isolated position of Allosorex among the late Miocene and Pliocene soricids and strengthens the separate Allosoricinae subfamily based on morphological reasoning ( Fejfar 1966).
D e s c r i p t i o n o f m a n d i b l e s. The mandible morphology ( Text-fig. 9 View Text-fig , Tab. 4) is very similar to both holotype and paratype; with only the following variations: the internal temporal fossa is ovally to triangularly rounded and very deep (Z 28217 – OF 651274 View Materials , Z 28218 – OF 6520, Z 28211 – OF 652357 View Materials , Z 28215 – OF 652366 View Materials ), distinctly bordered on all sides (Z 28217 – OF 651274 View Materials , Z 28218 – OF 6520, Z 28213 – OF 6523, Z 28215 – OF 652366 View Materials ); the medial surface above the internal temporal fossa is less shallow than in the holotype (Z 28213 – OF 6523 and Z 28215 – OF 652366 View Materials ) and quite smooth without tuberosities (Z 28211 – OF 652357 View Materials , Z 28210 – OF 652358 View Materials , Z 28216 – OF 6523, Z 28219 – OF 6523 ); the coronoid process forms an angle from approximately 105° (Z 28218 – OF 6520 ) through 111° (Z 28215 – OF 652366 View Materials ) , 116° (Z 28217 – OF 651274 View Materials ) , 118° (Z 28216 – OF 6523 ) , 120° (Z 28210 – OF 652358 View Materials ) to 140° (Z 28211 – OF 652357 View Materials ) and its anterior edge is straight (Z 28218 – OF 6520 ) or straight to slightly concave (Z 28217 – OF 651274 View Materials ) ; the caudo-ventrally oriented lateral coronoid spicule is more robust than in the holotype, but not as much as in the paratype (Z 28213 – OF 6523 and Z 28215 – OF 652366 View Materials ) or it can also be reduced (Z 28216 – OF 6523 and Z 28219 – OF 6523 ) ; a small circular opening is situated in the posterior portion of the external pterygoid fossa (Z 28218 – OF 6520 ) which can be uniformly deep throughout its length and distinctly bordered, especially on the ventral side (Z 28216 – OF 6523 ) , or it can be almost as deep as the internal temporal fossa (Z 28219 – OF 6523 ) ; and the upper condylar facet is rectangular (Z 28213 – OF 6523 ) .
D e s c r i p t i o n o f t h e h u m e r u s. The right humerus fragment (Z 28220 – OF 6523; Text-fig. 10 View Text-fig ) more or less consists only of the distal epiphysis with a spiral fracture in the diaphyseal portion. The condyle has irregular hourglass-shape in cranial view and it is divided by a relatively distinct groove into a larger capitulum and smaller trochlea, and this latter is very high on the caudal side. The coronoid and radial fossae are shallow and indistinctly separated from one another. The triangular olecranon fossa is also shallow, but relatively larger than the fossae on the cranial side. While the lateral and medial supracondylar ridges are distinctly developed, the slightly damaged lateral epicondyle is smaller than the medial epicondyle which is hooked and has a distinct edge. The lateral supracondylar foramen is relatively large, roughly oval and ventrally narrow. The following measurements were determined: the maximum distal epiphysis transverse diameter is 4.44 mm, the trochlea diameter is 2.06 mm, the trochlea upper transverse diameter is 1.69 mm and the trochlea maximum height is 1.56 mm.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.