
Paralimnadia cygnorum ( Dakin, 1914 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4161.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:8B9BDEA7-5F2B-465C-B2A8-757B733CCCE7 |
|
DOI |
https://doi.org/10.5281/zenodo.4685589 |
|
persistent identifier |
https://treatment.plazi.org/id/03E4878E-FFD6-FFEA-FF70-007B139CF813 |
|
treatment provided by |
Plazi |
|
scientific name |
Paralimnadia cygnorum ( Dakin, 1914 ) |
| status |
|
Paralimnadia cygnorum ( Dakin, 1914) View in CoL
( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 , 10 View FIGURE 10 )
Eulimnadia cygnorum Dakin, 1914: 295 View in CoL (list), 299, pl.1, figs. 9–13; Glauert, 1924:59. Limnadia cygnorum View in CoL .— Webb & Bell, 1979: 243, tab. 1; Brtek, 1997:57 (list). Paralimnadia cygnorum View in CoL .— Rogers et al., 2012: 838.
Type material. Neotype: WAM C61743, male 6.1 mm long by 4.1 mm high, Bradby Nature Reserve , Serpentine, Western Australia, 32°21’29.45”S, 115°58’26.89”E, 15 September 2011 GoogleMaps , BVT.
Other material examined. Western Australia: Bradby Nature Reserve, Serpentine, 32°21’29.45”S, 115°58’26.89”E, 15 September 2011, BVT, 2 males, 6.4 × 4.4 mm, 6.2 × 4.1 mm, 2 females 6.0 × 4.3 mm, 5.9 × 4.1 mm, WAM 61744 GoogleMaps ; Bradby Nature Reserve , Serpentine, 32°21’29.45”S, 115°58’26.89”E, 15 September 2011, BVT, 68 specimens, WAM C71745 GoogleMaps ; Bradby Nature Reserve , Serpentine, 32°21’29.45”S, 115°58’26.89”E, 7 September 2003, BVT, 88 specimens, WAM C71746 GoogleMaps ; Albany Highway , 10 August 2002, M. Davis, 21 specimens, WAM C71747 ; Esperance , 64 km NE, lake beach pool near Kau Rock, 33°24’32”S, 122°19’48”E, 29 January 2007, BVT, 48 specimens, WAM C61748 GoogleMaps .
Diagnosis. Egg spherical with 30–50 deep polygons each centred on a longitudinal groove and with steep internal slopes and edges frilly, perhaps extended into narrow outgrowths. Male rostrum larger than occipital outgrowth.Telsonic spines 16–20. Cercopod with <8 short setae on basal 40%. Clasper with projection at mediodistal corner of hand <10% of distal clapser width and first large plap with only 2 palpomeres though 2nd clasper with 3 palpomeres.
Description of neotype. Male. Head ( Fig. 10 View FIGURE 10 B) with ocular tubercle prominent, the compound eye occupying about 60% of its diameter. Rostrum robust, dorsal surface-frons angle about 100°, protruding about twice as much as ocular tubercle, and basal thickness about twice that of ocular tubercle. Rostrum apex broadly rounded and with growth lines. Ocellus slightly smaller than compound eye, triangular and lying at base of rostrum. Dorsal organ posterior to eye by about half its height, pedunculate and asymmetrical, not quite as high as ocular tubercle.
First antennae ( Fig. 10 View FIGURE 10 B) a little longer than peduncle of second antennae and also rostrum, and with nine lobes. Second antennae with spinose peduncle and dorsal flagellum with 12 antennomeres and ventral flagellum with 13 antennomeres, and dorsally with 1–3 short spines and ventrally with 1–5 longer setae. Distal antennomeres with minimal spines and maximal setae.
Carapace ( Fig. 10 View FIGURE 10 A) elongated oval with highest point midlength, pellucid but darker dorsally, and with hardly any indication of growth lines. Adductor muscle scar at about 45° to carapace long axis, only visible when animal removed from carapace.
Thoracopods. Eighteen pairs of thoracopods. Claspers ( Fig. 10 View FIGURE 10 D) with palm trapezoidal with a distinct endite III distomedially. Apical bulb spherical with many stout denticles and spines pointing medially, also laterally bearing small palp with many short thin spines apically. Finger arcuate with blunt apex bearing suctorial disc anteriordorsally and many rounded pits ventrally. Both long palps of claspers inserted on apical edge of palm, first with 2 palpomeres, second with 3, palpomere junctions inerm. Long palp of first clasper about 1.25× length of palm and long palp of second clasper 2.25× length of palm. Flattened palaform apices of both palps with numerous limp setae. Other thoracopods of typical structure for Paralimnadia , decreasing is size and complexity posteriorly. Dorsum of last 10 segments with 1–3 spines distomedially.
Telson ( Fig. 10 View FIGURE 10 C) with spine rows bearing about 18 pairs of spines, the first few and last few the longest. Spines with spinulae. Telsonic filaments originating from mound little higher than dorsal floor of telson positioned between fourth and fifth spine pairs. Steep declivity posterior to mound followed by slow descent to cercopod base. Cercopods almost as long as dorsum of telson; basal 40% hardly thinning to small spine then distal 60% thinning rapidly to acute apex. About 7 setae of moderate length (length 1.5 × basal cercopod diameter). Setae geniculate and plumose. Numerous denticles on dorsolateral edges of distal 60% of cercopod.Ventroposterior corner of telson with denticulated triangular protrusion.
Female. Head ( Fig. 10 View FIGURE 10 F) with ocular tubercle prominent, with compound eye occupying about 90% of it. Rostrum asymmetrical, bulkier than the head, protruding about as much as the ocular tubercle, but basal width about 1.5× that of tubercle. Ocellus middorsal on rostrum, about half size of eye. Dorsal organ posterior to eye by about half of its height, pedunculate and asymmetrical and about threequarters length of ocular tubercle.
First antennae ( Fig. 10 View FIGURE 10 F) and peduncle of second antennae subequal in length, with 5 lobes. Second antennae largely as in male.
Carapace ( Fig. 10 View FIGURE 10 E) as in male, though slightly more vaulted dorsally.
Thoracopods. Eighteen thoracopods of typical Paralimnadia structure. Trunk dorsum with segments 1–9 inerm, segments 8–14 with 3–7 setae/spines medioterminally and segments 15–18 with 1 spine distomedially.Thoracopods 9 and 10 with long flabellum dorsally.
Telson ( Fig. 10 View FIGURE 10 G) as in male, though with about 22 posterior row spines, otherwise with cercopod similar.
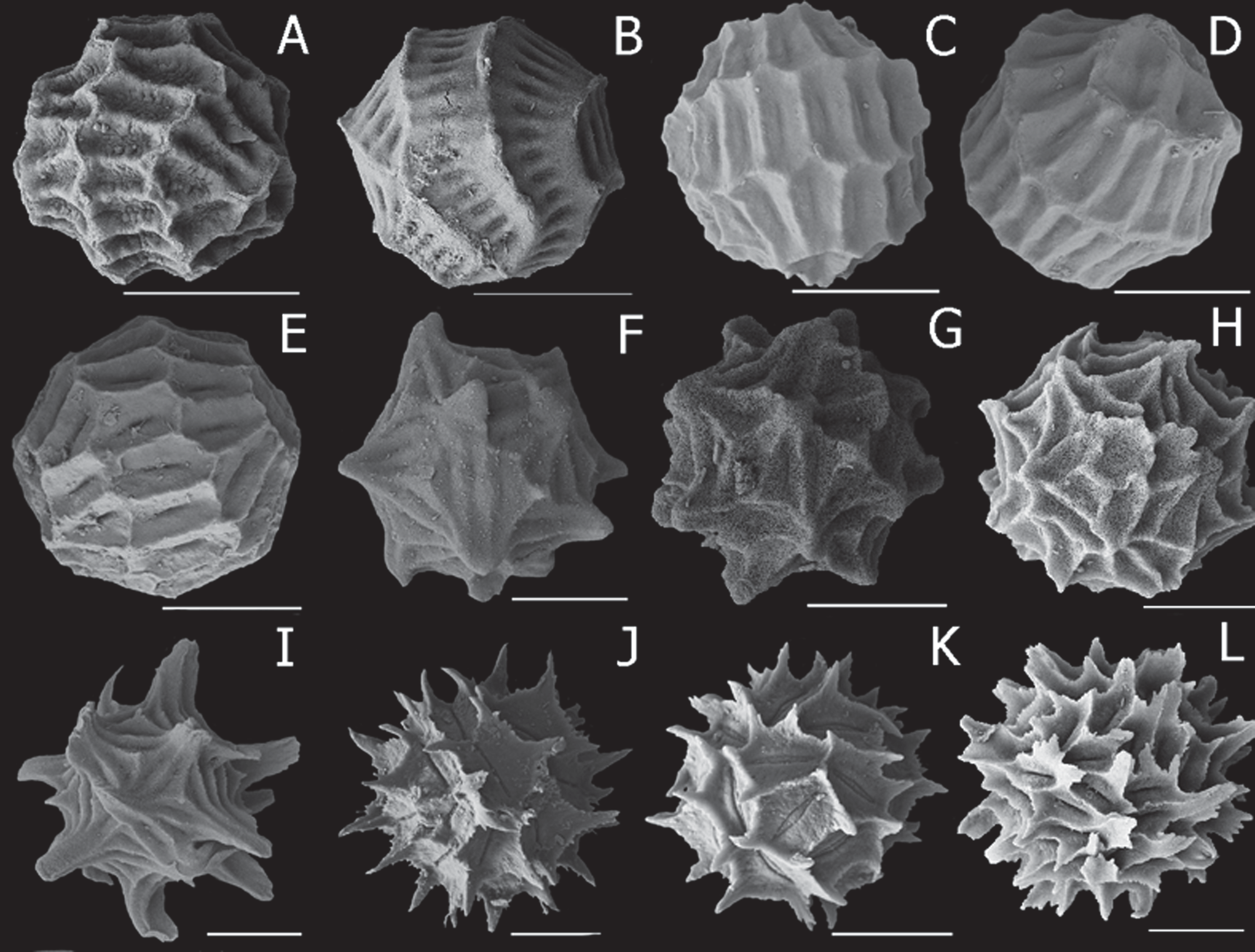
Egg ( Fig. 5 View FIGURE 5 K, L). Egg spherical (diameter 315 and 324 µm in two populations and 216 µm and 235 µm in two other populations, n = 20) with many projections from the 30–50 polygons. Each polygon elongated around central groove and with projections arising from most angles, longer in some populations than others. Groove usually deep within sloping floor and polygon walls steep and narrow.
Variability. Specimens from the four sites exhibited the usual variation in meristic characters with ±1 for antennal units and ±2 for cercopod setae and ±3 for telsonic spines. Rostra thick-narrow, proportions in the cercopod seem relatively invariant. These characters are slightly different from what Dakin’s diagrams of male structures show, but within typical levels of variability.
Differential diagnosis. The egg with its deep polygonal structure is unique among Paralimnadia . Other Paralimnadia with cercopod setae short and fewer than 10 include P. stanleyana , P. hyposalina n. sp., P. monaro n. sp., P. multispinosa n. sp., and P. saxitalis n. sp. Paralimnadia stanleyana is separated by having variable length cercopod setae and a large projection at the mediodistal corner of the hand. Paralimnadia hyposalina n. sp. has a much smaller projection and has spines at lower palpomere junctions; P. monaro n. sp. also has these palpomere spines and both large palps with an incompletely annulated second palpomere, P. multispinosa n. sp. has>22 telsonic denticles and an inermic section of the basal part of cercopod and P. saxitalis n. sp. has only 7 first antennal lobes and also spines at lower palpomere junctions.
Remarks. Dakin’s (1914) description and diagrams of this species could be applied to many species of Paralimnadia , certainly to those with a robust triangular male rostrum, a typical Paralimnadia clasper, a telson with about 20 telsonic spines and few cercopod setae. Unfortunately, the syntypes in the Western Australian Museum (C339, 3 specimens) have disintegrated and are beyond study. There are just two clues to its identity. The first is the few (four in Dakin’s 1914: pl. 1, fig. 12) short cercopod setae. The second clue is the type locality is Cannington near the Swan River, Perth. Two collections from the sandy coastal plain of Perth and likely to be similar to the site at Cannington (now urbanised) are available and thought to be of this species (and certainly not of other described species). A neotype has been erected from one, and two other collections from further south in Western Australia also contain similar specimens.
Distribution. To date, Paralimandia cygnorum is known mainly from coastal sandy locations, from about Perth to Esperance ( Fig. 4 View FIGURE 4 ).


| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paralimnadia cygnorum ( Dakin, 1914 )
| Timms, Brian V. 2016 |
Eulimnadia cygnorum
| Rogers 2012: 838 |
| Brtek 1997: 57 |
| Webb 1979: 243 |
| Glauert 1924: 59 |
| Dakin 1914: 295 |