
Hiatella umbonata, Zelaya and Marina Güller, 2023
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlad064 |
|
publication LSID |
lsid:zoobank.org:pub:9DCD56DB-3954-47CC-A2C5-483409D3958BC |
|
DOI |
https://doi.org/10.5281/zenodo.10474100 |
|
persistent identifier |
https://treatment.plazi.org/id/03E587A0-FFCE-FFDC-FEE5-FABBDB7CFBD8 |
|
treatment provided by |
Plazi |
|
scientific name |
Hiatella umbonata |
| status |
sp. nov. |
Hiatella umbonata View in CoL sp.nov.
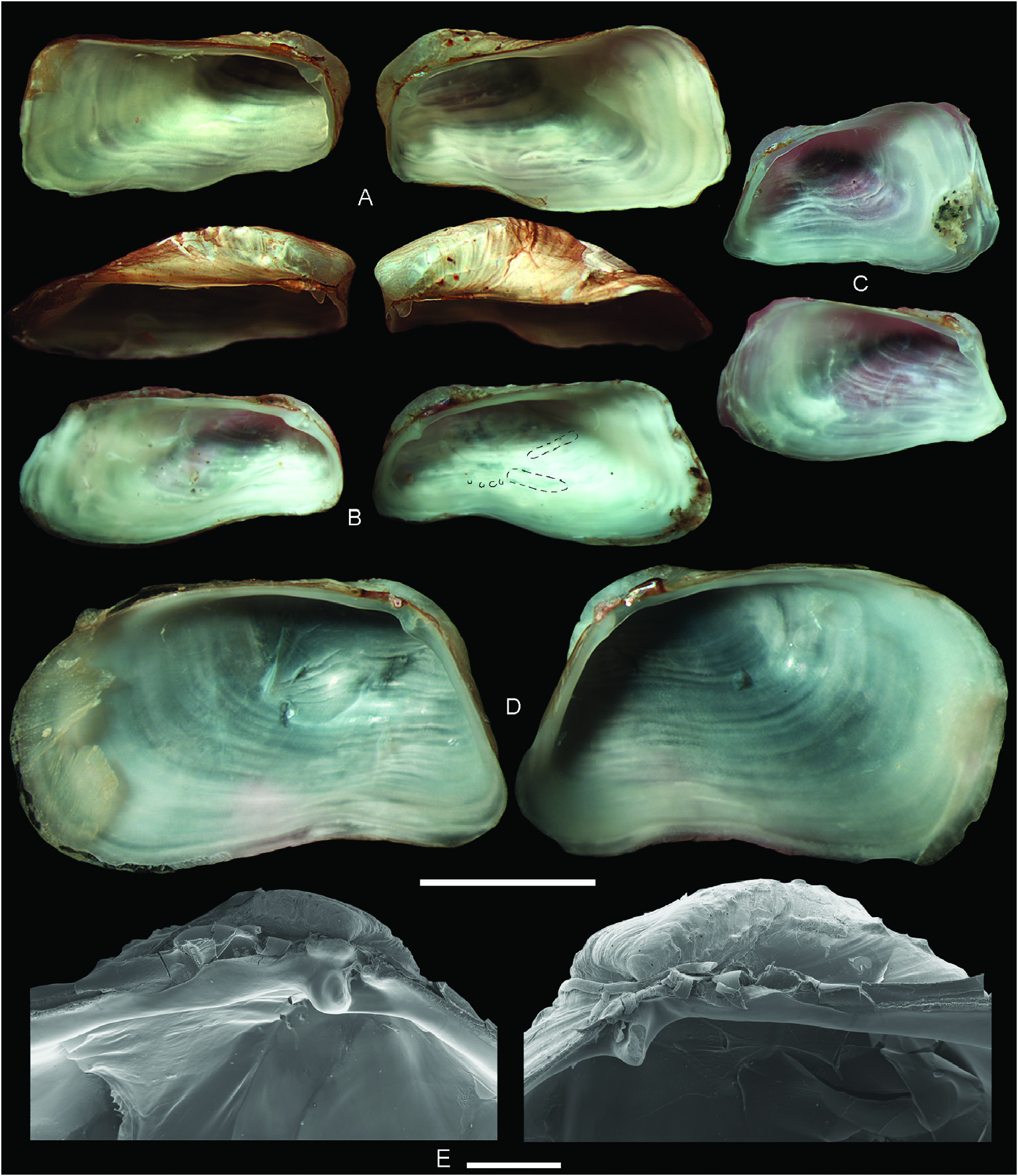
( Figs 3M–O View Figure 3 , 6C, D, H, K View Figure 6 , 7A–E View Figure 7 , 8 View Figure 8 , 9 View Figure 9 )
Zoobank LSID: urn:lsid:zoobank.org:act:7E5C1E17-9183-4BE1-8618-37FDC8B1C421
Saxicava arctica var. antarctica View in CoL : Melvill and Standen 1907: 151 (in part) (not Philippi, 1845).
Hiatella solida View in CoL : Branch et al. 1991: 47, 51 (not Saxicava solida Sowerby I, 1834 View in CoL ).
Hiatella sp : Zelaya 2005: 117.
Hiatella meridionalis View in CoL : Simone and Penchaszadeh 2008: 119–127 (not fig. 18); Schejter et al. 2010: 63–67 (not Saxicava meriodionalis d’Orbigny, 1846 ).
Hiatella View in CoL O: Güller et al. 2020: 143, fig. 3.
Type locality: 54°44 ʹ 49″ S, 59°56 ʹ 51″ W, Marine Protected Area Namuncurá I, Burdwood Bank, 177 m depth, B.O. Puerto Deseado, April 2016 GoogleMaps .
Type material: Holotype (MACN-In 44195: H9) and one paratype (MACN-In 44196: H10) from the type locality.
Etymology: The name derives from the peculiar umbo of this species.
Additional material examined: 94 lots (Supporting Information, ESM 1).
Diagnosis: Shell thick, inflated, with anterior end truncated or only slightly projected. Umbo wide, inflated, obliquely truncated, located at or close to anterior end. Outer shell surface with low and wide commarginal folds. Posterior area of shell wide, defined by a well-developed but narrow umbonal ridge. Hinge with teeth throughout the shell ontogeny. Nymph narrow.
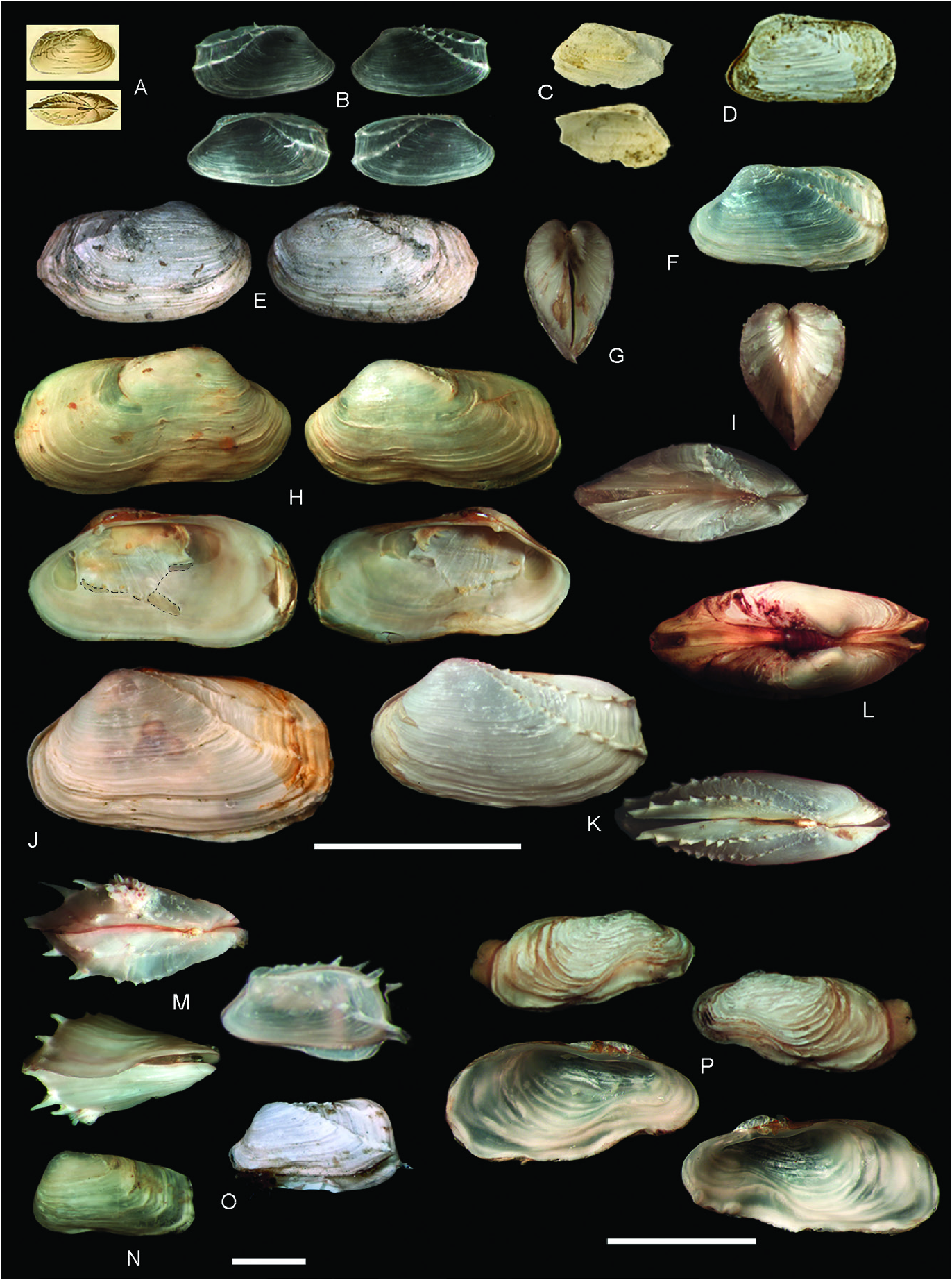
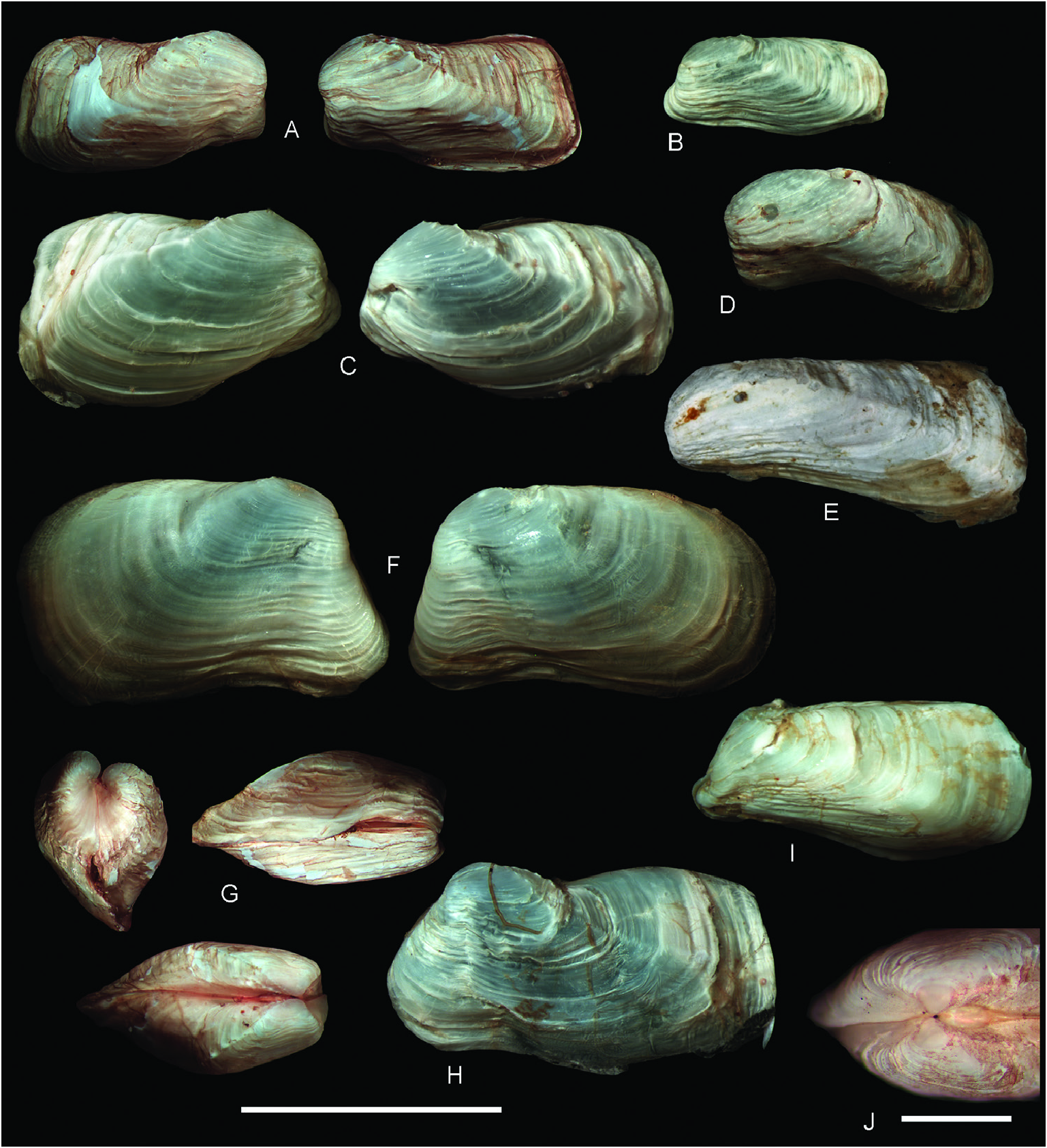
Description: Shell ≤ 23.0 mm long, usually low and rectangular, sometimes high and square or trapezoidal, sometimes greatly distorted; moderately to widely inflated, with the maximum width at about the posterior third of total shell length; very solid ( Figs 3M–O View Figure 3 , 8 View Figure 8 ). Markedly inequivalve throughout ontogeny: right valve larger and wider than less valve, ventrally overlapping over less valve. Posterior end greatly projected, sometimes curved or recurved; anterior end abbreviated, truncated, rounded or shortly pointed, lower than posterior end. Posterior area of shell wide, depressed to flat; usually defined by a narrow, rounded to keeled umbonal ridge, indistinct in some specimens. Ventral and posterior gapes between valves small. Anterior area usually with a narrow sulcus, running from the umbo to the ventral margin. Anterodorsal margin frequently indistinct from anterior margin; when distinct, straight and steeply sloping obliquely. Anterior margin short, straight to slightly curved. Ventral margin straight to slightly sinuous. Posterior margin high, obliquely straight to widely arched, exceptionally sinuous. Posterodorsal margin long, horizontally straight to markedly arcuated dorsally or ventrally; sometimes with a sinuation interlocking the opposite valve. Umbo prominent, wide, inflated, usually high but sometimes low, obliquely truncated, higher posteriorly than anteriorly; located at or close to anterior end. Right valve umbo larger than less valve umbo. Lunule elongated, depressed. Outer shell surface white, chalky. Dissoconch sculptured with irregularly distributed, low and rounded commarginal folds, increasing in solidness ventrally. Some folds (primary folds) relatively wide; space between primary folds with thinner (secondary) folds. Sometimes dissoconch showing strong growth disruption, giving the shell a gradated appearance. Posterior end with two radial ribs: one of them running from prodissoconch/dissoconch limit to the junction of the posterior and ventral margins (umbonal ridge) and the other to the junction of the dorsal and posterior margins; the laưer weaker than the former. In specimens <6 mm long, the two radial ribs bear hollow spines, which increase in size distally. In larger specimens, spines do not develop, and early formed spines (in the vicinity of the umbo) appear reduced to their bases, resembling tubercles. Periostracum thin, yellowish straw coloured, forming thin periostracal folds.
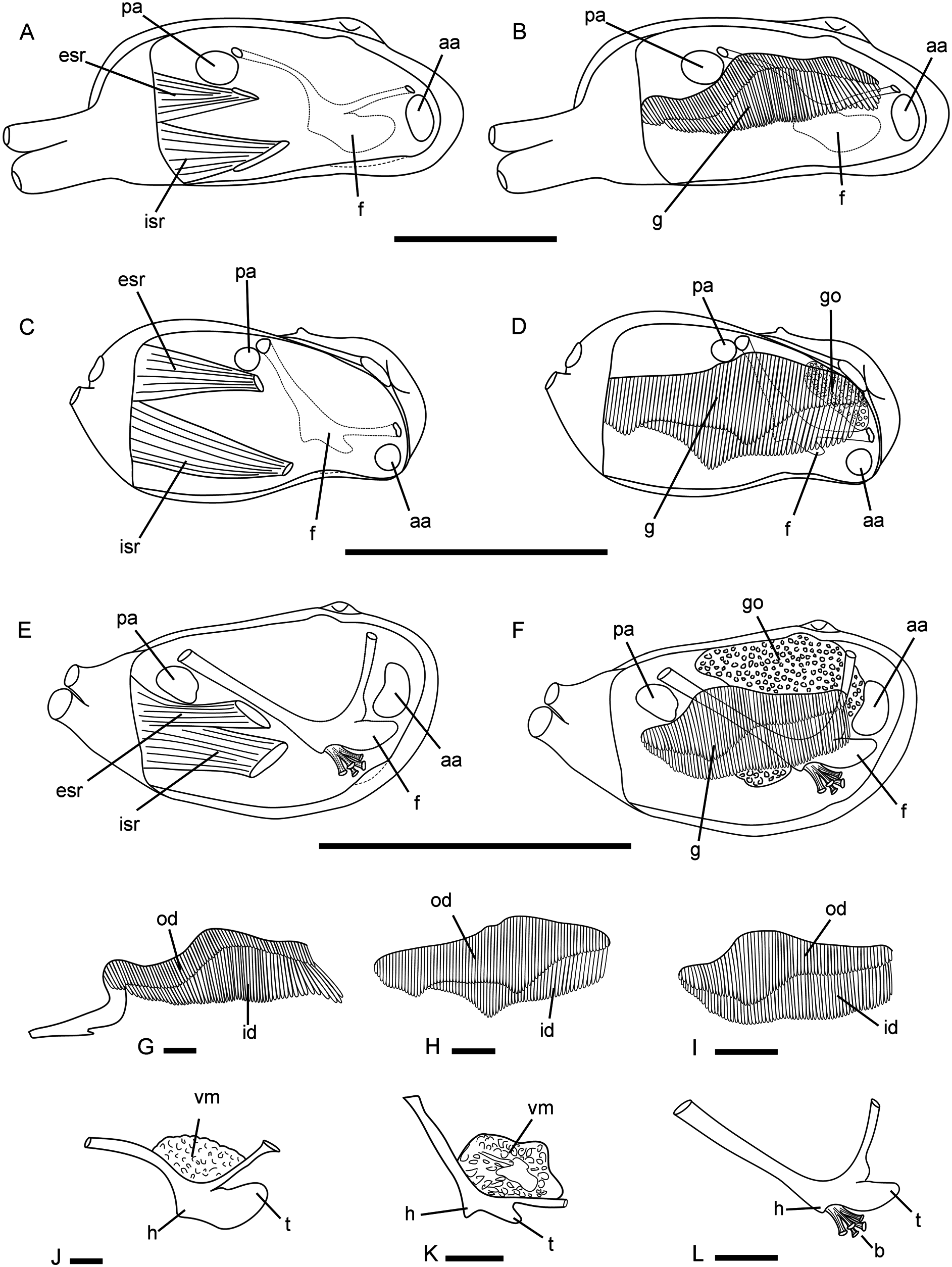
Hinge ( Fig. 9 View Figure 9 ): With a single, small cardinal tooth at each valve and a small, triangular socket to accommodate the tooth of the opposite valve. Less valve cardinal triangular, pustular to massive, high to low, sharply to bluntly pointed. Right valve cardinal delicate to solid, elongate, ventrally directed, smaller than less cardinal. Less valve socket anterior to cardinal tooth; right valve socket posterior to cardinal tooth. Teeth present throughout ontogeny. Nymph elongate, narrow, delicate. Ligament strong, external, opisthodetic. Pallial sinus deep. Inner shell surface porcellanaceous, white.
Anatomy ( Fig. 6C, D, H, K View Figure 6 ): Mantle margin fused for most of its length, with a small (anterior) pedal opening and two smaller (posterior) siphonal openings. Suture behind pedal opening large. Anterior and posterior adductor muscles small, subcircular in section, the anterior one anteroventrally located, the posterior dorsally displaced. Gills high (~80% of shell height), composed of two complete demibranchs at each side; less and right demibranchs posteriorly fused among them and to mantle margin. Outer demibranch gradually increasing in size backwards, with the maximum height at about the middle gill length. At anterior end, outer demibranch two-thirds the height of inner demibranch. At anterior half of the gill, ascending lamella of outer demibranch higher than descending lamella, and descending lamella of inner demibranch higher than ascending lamella. At the posterior half of the gill, inner and outer demibranchs, and ascending and descending lamellae of each demibranch, similar in height. Foot minute, composed of a compressed stalk, a small heel and a somewhat projected anterior ‘toe’; base narrow, flat, with byssal groove, extending for ~85% of foot sole. Byssus composed of numerous, narrow and flat byssus threads arising from a stout byssus seam; each byssus thread bearing a distal adhesive disc. Byssal gland functional in adult specimens. Siphons short, fused at the base, distally separated, grey to pinkish in living specimens ( Fig. 7A View Figure 7 ); both with numerous series of club-shaped papillae surrounding their openings. Inhalant siphon retractor muscle longer than exhalant siphon retractor muscle. Anterior and posterior labial palps small, triangular, with ≤ 10 sorting ridges. Gonad well developed in specimens> 8 mm long. Additional anatomical characters for this species were described by Simone and Penchaszadeh (2008: as ‘ Hiatella meridionalis ’).
Habitat: Seưlement occurs on the hydroid Symplectoscyphus subdichotomus (Kirchenpauer, 1884) ( Schejter et al. 2010) . In the northernmost distributional range, adult specimens were found as epibionts on the pectinid Zygochlamys patagonica (King, 1832) ( Fig. 7E View Figure 7 ), the gastropods Fusitriton magellanicus (Röding, 1798) ( Fig. 7B View Figure 7 ) and Trophon sp. , and polychaete tubes. In the southernmost distribution range, the species also appears as an epibiont on the hydrozoan Errina antarctica (Gray, 1872) and the kelp Macrocystis pyrifera (Linnaeus) C. Agardh, 1820 . Hiatella umbonata is also found nestling on barnacle shells and sponges ( Fig. 7C, D View Figure 7 ).
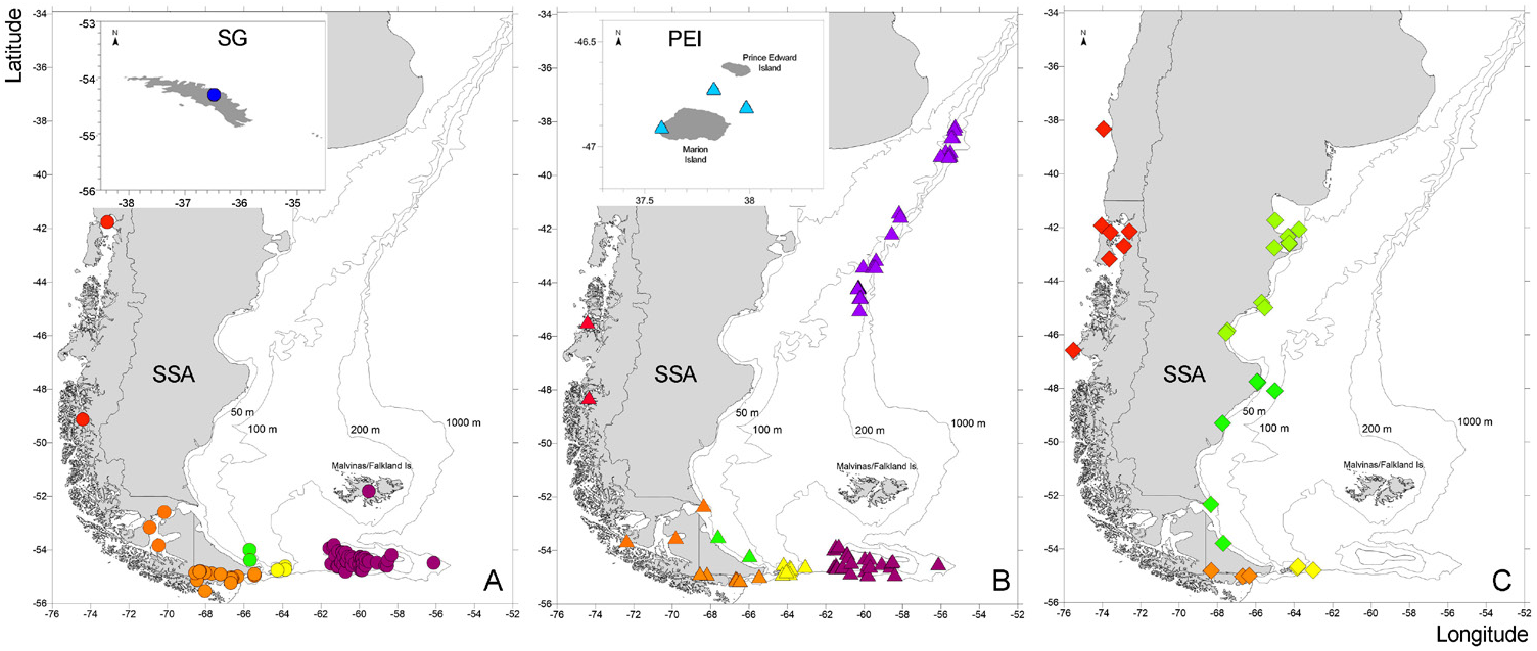
Distribution: Along the South American coast, from 45°27 ʹ 48″ S in the Pacific to the Atlantic coast of Tierra del Fuego; Magellan Strait; Isla de los Estados; Burdwood Bank; and extending northwards along the outer Argentine continental margin to 38°09 ʹ 03″ S. Also present in Prince Edward Islands ( Fig. 1B View Figure 1 ). From 7.6 to 785 m depth.
Remarks: Hiatella umbonata is widely variable in shell outline, projection of the anterior end, height of the umbo, development of commarginal and radial sculpture and solidness of hinge teeth. This variability is evident not only when considering specimens from different areas but also among specimens from a single site. However, the study of a large series of specimens and the combination of morphological, anatomical and molecular data allow us to confirm that all the variability described above represents the intraspecific variability of a single species.
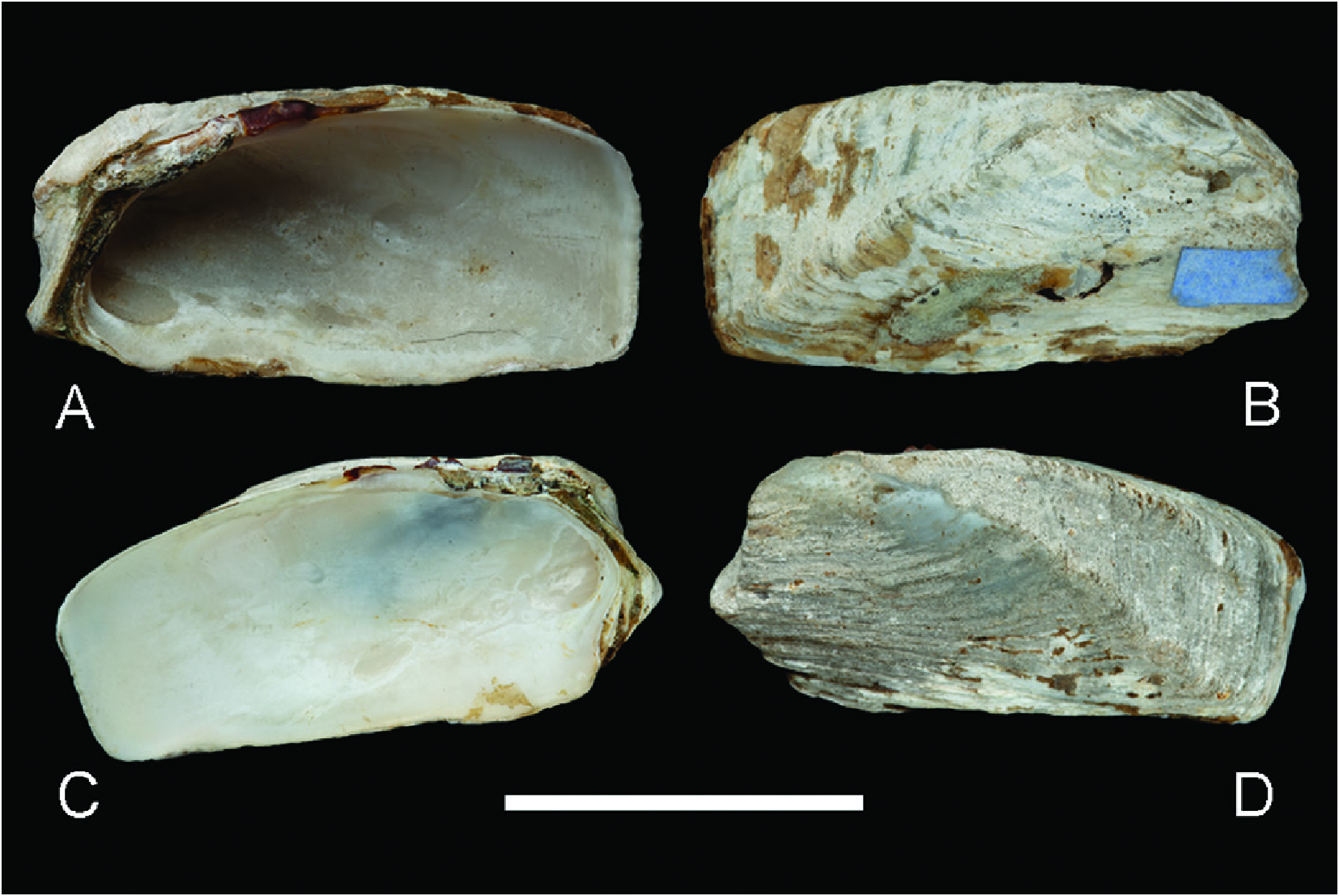
Hiatella umbonata View in CoL resembles H. solida (Sowerby I, 1834) View in CoL by having elongated shells and anteriorly displaced umbos. The laưer is a species that was based on specimens from ‘Sanctam Elenam’ (= Santa Elena, Ecuador); although the end of the original description reads ‘there are other varieties from Payta and the Isle of Muerte’ ( Perú). With the information available so far, it is impossible to determine whether Sowerby’s I (1834) ‘varieties’ correspond to subspecies of the same species he described from Ecuador or a different species. To increase the confusion, Sowerby II (1875, 1884) reported this species as coming from an unknown locality. The original description of H. solida View in CoL was not accompanied by an illustration. The first illustration was provided by Sowerby II (1875: species 6; subsequently copied by Sowerby II 1884: pl. 471, fig. 12), showing only the outer shell surface. Dell (1964) regarded the figured specimen (34.5 mm long) ‘as the type’ (= lectotype, according to the ICZN Art. 74.5); although alerting that there is no information on the provenance of this material associated with the lot. Photographs of the lectotype (NHMUK 20220347) of this species are reproduced herein ( Fig. 10 View Figure 10 ). Hiatella umbonata View in CoL clearly differs from the lectotype of H. solida View in CoL by having a more prominent umbo, narrower posterior radial ribs, smaller anterior adductor muscle scars, and the nymph situated close to the umbo instead of posteriorly displaced. Furthermore, all the specimens of H. umbonata View in CoL studied herein (independently of their size) show hinge teeth and the corresponding depression to accommodate the tooth of the opposite valve, whereas these elements are absent in the lectotype of H. solida View in CoL .
Hiatella umbonata View in CoL also resembles morphologically the Brazilian specimens identified by Narchi(1973) and Domaneschi and Narchi (1998) as H. solida View in CoL . Although the correct identification of that material deserves further study, the Brazilian specimens differ strikingly from the new species described herein by having two cardinal teeth in the less valve ( Narchi 1973), with at least one of the hinge teeth wearing away in larger specimens ( Domaneschi and Narchi 1998); a large and ovoid anterior adductor muscle; long and bright orange–yellow siphons; the outer demibranch half the height of the inner demibranch, and the labial palps with 16 ridges. Furthermore, the Brazilian specimens inhabit the intertidal zone, where they are found nestling or boring.
In the Magellan Region, H. umbonata View in CoL is frequently found coexisting with H. meridionalis View in CoL , from which it differs by having a thicker shell, with broader posterior area; shorter anterior end; wider, higher and more anteriorly displaced umbos; and stronger and more irregular commarginal sculpture. Anatomically, H. umbonata View in CoL differs from H. meridionalis View in CoL by having smaller adductor muscles. This fact is particularly evident in the case of the anterior adductor muscle, which in H. umbonata View in CoL is small, subcircular and restricted to the anteroventral part, whereas in H. meridionalis View in CoL it is large, occupying most of the anterior end, and ovoid, dorsally projected. Another difference arises in the gill, which is higher in H. umbonata View in CoL than in H. meridionalis View in CoL (~80% vs. 40% of shell height, respectively), with the outer demibranch reaching the maximum height in the middle of its length, whereas in H. meridionalis View in CoL the height of the outer demibranch is uniform throughout its length. Furthermore, H. umbonata View in CoL has a smaller foot, with a less projected anterior region than that of H. meridionalis View in CoL . All the above-mentioned anatomical differences are evident independently of the size of the specimens considered.
The morphological and anatomical characteristics recognized herein for H. umbonata are also recognizable in the specimens from off Mar del Plata, Buenos Aires Province, Argentina, identified by Simone and Penchaszadeh (2008) as H. meridionalis . Consequently, these specimens are here reassigned to H. umbonata . The material identified as Hiatella O in the Molecular Study section above, and by Güller et al. (2020), also corresponds to H. umbonata , as is also the case for some of the specimens from Burdwood Bank reported by Melvill and Standen (1907) as S. arctica var. antarctica . The specimens reported by Schejter et al. (2010: as H. meridionalis ) and Branch et al. (1991: as H. solida ) also seem to correspond to H. umbonata .
The persistence of hinge teeth in large specimens clearly allows the distinction of H. umbonata from the Northern Hemisphere ‘ H. solida ’, in which the teeth disappear with growth (e.g. Dell 1964).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hiatella umbonata
| Zelaya, Diego G. & Güller, Marina 2023 |
Hiatella meridionalis
| Schejter L & Bremec CS & Waloszek D 2010: 63 |
| Simone LRL & Penchaszadeh PE 2008: 119 |
Hiatella sp
| Zelaya DG 2005: 117 |
Hiatella solida
| Branch ML & Arnaud PM & Cantera J 1991: 47 |
Saxicava arctica var. antarctica
| Melvill JC & Standen R 1907: 151 |