
Grismadox elsneri, Perger & Rubio & Pett, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5168.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:784A7D6E-934D-4D8F-ACEA-015D022F307F |
|
DOI |
https://doi.org/10.5281/zenodo.6899762 |
|
persistent identifier |
https://treatment.plazi.org/id/01D3DC32-0C88-458A-8440-48012FFCE6A3 |
|
taxon LSID |
lsid:zoobank.org:act:01D3DC32-0C88-458A-8440-48012FFCE6A3 |
|
treatment provided by |
Plazi |
|
scientific name |
Grismadox elsneri |
| status |
sp. nov. |
Grismadox elsneri sp. nov.
(LSID: urn:lsid:zoobank.org:act:01D3DC32-0C88-458A-8440-48012FFCE6A3 )
Figs 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5
Type material. Holotype ♂ from BOLIVIA: Santa Cruz department, Santa Cruz de la Colina, Urubo (-17.7608°; -63.24°), 432 m a.s.l., 28 Dec 2019, leg. R. Perger, Cerrado-like savanna grass adjacent to fragment of Chiquitano forest (IBSI-Ara 1710) . Paratypes: 1♂, 1♀, same data as holotype (IBSI-Ara 1711) ; 1♂, 5♀, same location as holotype, 25-30 Apr 2021, leg. R. Perger ( CBF).
Etymology. The specific epithet, elsneri , is a patronym in honor of Hans-Peter Elsner in recognition of his efforts to operate an ecologically sustainable and employee-friendly ranch in the Beni department, Bolivia, and his support of several research projects on his property. One such project led to the discovery of an unknown Grismadox species that was described by Pett et al. (2022).
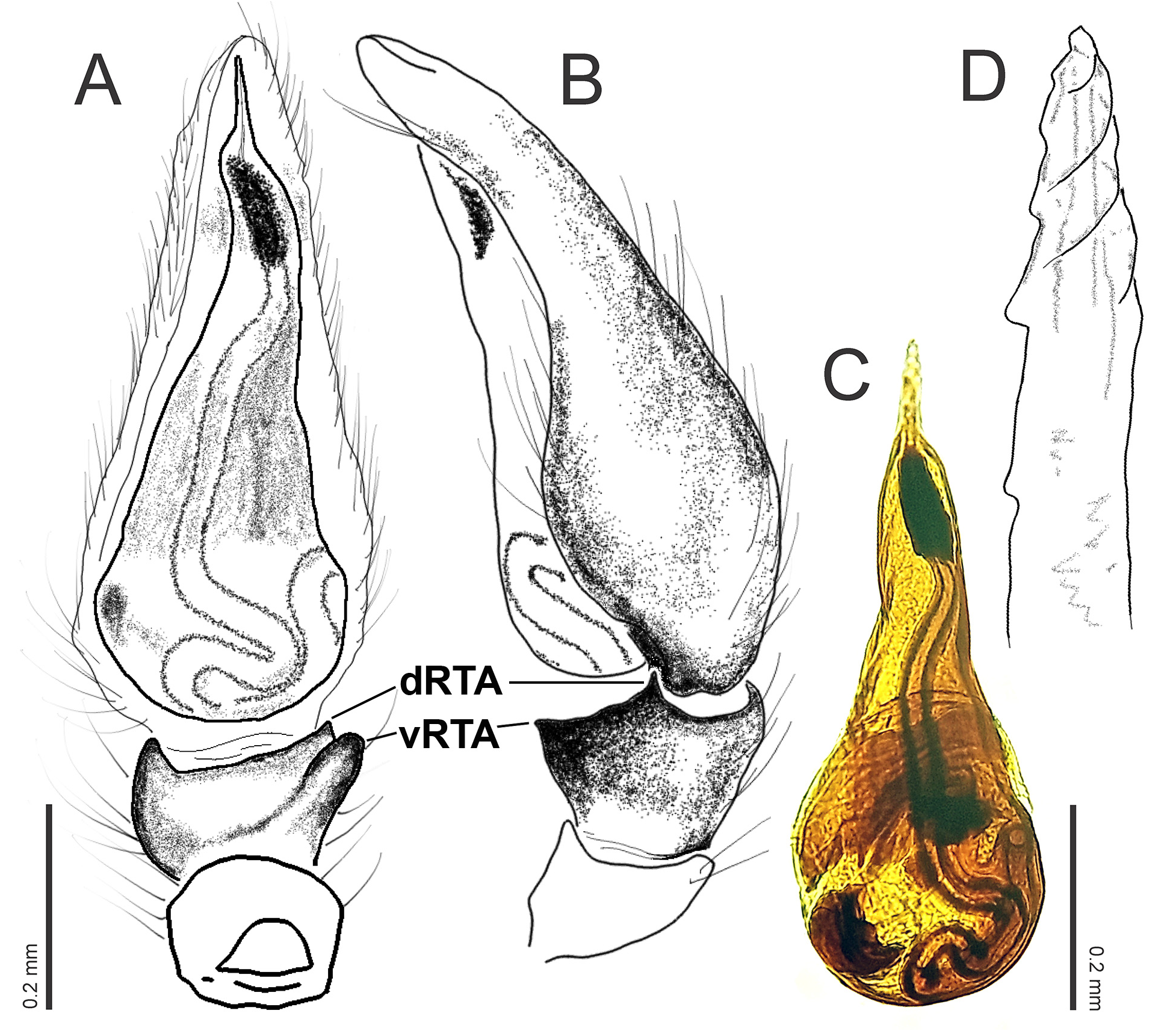
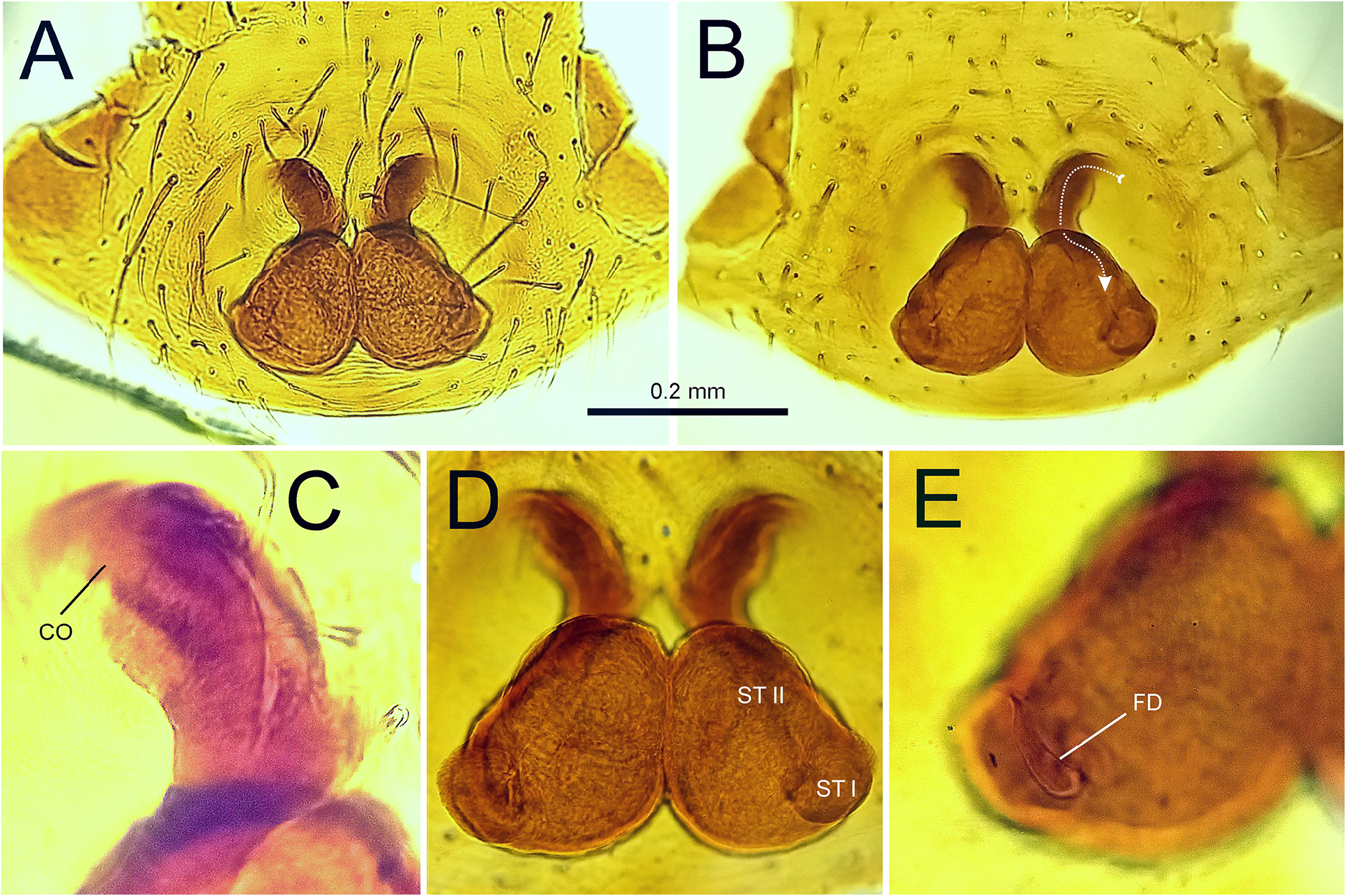
Diagnosis. Grismadox elsneri sp. nov. can be readily separated from congeners by having a comparably short and obovate abdomen (abdominal index 60–67.5; in congeners 38–50) with an indistinct to moderate constriction ( Figs 2 View FIGURE 2 , 4 View FIGURE 4 ). The reproductive structures of specimens of G. elsneri sp. nov. differ from other Grismadox species by the following combination of characters: dRTA small, vRTA blunt and rounded in ventral view ( Fig. 3A, B View FIGURE 3 ), E slender and pointed, coils less conspicuous ( Fig. 3D View FIGURE 3 ), and ST breast-shaped ( Fig. 5 View FIGURE 5 ).
Description of male holotype.
Body length 3.97; carapace length 1.96, width 1.11, carapace width index 56.63; cephalic width 0.67, cephalic width index 60.36; sternum length 0.81, width 0.61, sternum index 75.3; abdomen length 1.75, width 1.05, abdominal index 60; petiole length 0.14, width 0.25; dorsal sclerite length and width as in abdomen; epigastric sclerite length 0.48, width 0.63; ventral sclerite length 0.85, width 0.58; inframamillary sclerite length 0.19, width 0.41. AER 0.44; AME–AME 0.08; AME–ALE 0.03; PER 0.61; PME–PME 0.15; PME–PLE 0.10.
Color and microsculpture. Dorsal integument dark-grey blackish (faded to dark-brown due to storage in ethanol), weakly shiny, smooth; microsculpture reticulate, with evenly distributed fine pits; chelicerae brown, moderately shiny; microsculpture of abdomen reticulate, with evenly distributed pits that are denser and larger than on carapace; coxae II and III translucent white, remainder of coxae dark brown; trochanters I–III translucent whitish-yellow, with dark-brown margins; femora I and II translucent whitish, broad black bands along edges, proximal fifth somewhat darker, particularly on femur I; remainder of leg I and II yellowish to light-brown; femora and tibiae III and IV dark brown, edges lined with bands of short, appressed, white feathery setae, patellae III and IV translucent white ventrally, metatarsi and tarsi III and IV light brown.
Setation. Integument densely covered by appressed, whitish, simple and feathery setae except two rounded, bare areas on thoracic part, setae providing an anthracite-greyish, somewhat silvery appearance in live condition ( Fig. 7B View FIGURE 7 ) (setae mostly abraded due to storage in ethanol), several relatively long, forward-pointing, dark setae on front of cephalic area.
Anterior pair of abdominal setae simple, not sclerotized to spines, dark, first pair strongly reduced, almost as short as feathery setae, second pair short, indistinct; abdomen with three transverse bands of light grey, feathery setae, two on proximal half of abdomen and one broader band at the middle, with feathery setae larger and denser than in the other bands, posteriorly followed by broad, bare area, and distal fourth with separate, long, erected light grey/whitish and brassy/goldish setae.
Carapace. Obovate, with cephalic area somewhat narrowed laterally, front slightly convex; carapace widest in middle, evenly narrowing in posterior direction, posterior margin straight ( Fig. 2A, C View FIGURE 2 ).
Eyes. Eight sub-equal eyes formed in two rows; PER and AER slightly recurved.
Chelicerae. Two retromarginal teeth and two promarginal teeth, all very small, denticle-like.
Abdomen. Obovate; anterior margin of petiole concave; petiole and proximal area of dorsal sclerite with transverse wrinkles, dorsal sclerite completely covering abdomen dorsally and laterally, indistinct transverse constriction medially; ventral sclerite fully developed, covering area between ventro-lateral margins of dorsal scutum and between epigastric and inframamillary sclerites; inframamillary sclerite narrow, subrectangular ( Fig. 2 View FIGURE 2 ).
Palp. Tibia with two pointed retrolateral apophyses, vRTA projected disto-laterally, blunt, rounded in ventral view, dRTA shorter and distally oriented ( Fig. 3A, B View FIGURE 3 ). Palpal bulb with basally three loops in sperm duct ( Fig. 3C View FIGURE 3 ). E slender and pointed, screw-like, with four shallow coils, very thin and sharp apex ( Fig. 3D View FIGURE 3 ).
Female paratype. Body length 5.00; carapace length 2.46, width 1.31; carapace width index 53.25; cephalic width 0.90, cephalic width index 68.7; sternum length 1.09, width 0.74, sternum index 67.88; abdomen length 2.34, width 1.58, abdominal index 67.52; dorsal sclerite length 1.23, width 1.13; epigastric sclerite length 0.67, width 1.00; inframamillary sclerite length 0.13; width 0.29. AER 0.50; AME–AME 0.14; AME–ALE 0.05; PER 0.72; PME–PME 0.20; PME–PLE 0.13.
Lateral carapace border slightly convex at height of coxae II, III and IV ( Fig. 4A View FIGURE 4 ); abdomen broader than in male, particularly anterior half, dorsal sclerite orbicular, 53% of abdomen length, slight constriction posterior to dorsal sclerite, constriction more pronounced than in male, ventral sclerite absent, white setae on distal half of abdomen arranged in two loosely defined bands ( Figs 6B View FIGURE 6 , 7A View FIGURE 7 ); coxae II–IV translucent white; remaining somatic characters as in male.
Chelicerae. Two retromarginal teeth and two promarginal teeth.
Epigyne. Epigynal plate forming part of the epigastric sclerite. COs and CDs situated anterior to ST, COs directed laterally, subapical to CDs; ST breast-shaped, posteriorly wider than anteriorly. Inconspicuous separation between ST I and ST II; FDs dorsal, situated at postero-lateral margin of ST I ( Fig. 5 View FIGURE 5 ).
Variation. Females were larger (BL 4.73–5.29 mm) than males (BL 3.71–3.98 mm). Sexual dimorphism in coxae color (coxae II–III translucent whitish in male, II–IV in female). The density of the light feathery setae on the carapace varied between the specimens. Considering that the setae may have been abraded in some specimens under live conditions (e.g., see Fig. 6B View FIGURE 6 ), more samples are needed to assess the variability of this character. The extent of the constriction in the female abdomen varied according to the nutritional status; a female with a narrow abdomen and distinct constriction was also observed. In males, the shape of the abdomen is determined by the strongly sclerotized dorsal sclerite, which results in a narrower abdomen than in females. Except for the variability of the carapace setae, abdominal constriction and sexual dimorphism, there was no visible intraspecific variation in adult spiders.
Geographical and ecological distribution. This species is only known from the type locality in Santa Cruz de la Colina, Urubo, Santa Cruz department, Bolivia. Specimens were collected on savanna grass plants and the ground between those plants along an edge of Chiquitano forest ( Fig. 6A View FIGURE 6 ). The grass plants were about between 0.5–1.2 m high and formed a relatively continuous layer, concealing the sandy soil. The spiders became visible only after moving the grass plants slowly aside.
On the ground, the recently redescribed Castianeira spinipalpis Mello-Leitão, 1945 (see: Pett & Perger 2022) and Mazax akephaloi Perger & Pett, 2022 were observed. Another Bolivian species of Grismadox , G. mazaxoides , was found about 300 meters away from the forest edge in more open grassland in close co-occurrence with its putative ant models, Camponotus cf. melanoticus Emery, 1894 or C. cf. punctulatus Mayr, 1868 . Despite high sampling effort in several Bolivian forest ecoregions ( Perger & Perger 2017; Perger & Rubio 2020a, b, 2021), including the adjacent fragment of Chiquitano forest (unpubl. data), G. elsneri sp. nov. was not observed in forest habitats.
Ant mimicry. Two carpenter ant species, Camponotus cf. crassus Mayr, 1862 (mean BL 4.89 mm in minor workers, n=11) ( Fig. 7C View FIGURE 7 ) and C. cf. blandus (F. Smith, 1858) (mean BL 5.1 mm in minor workers, n=15) ( Fig. 7D View FIGURE 7 ), and Pseudomyrmex termitarius (F. Smith, 1855) were observed foraging together with specimens of G. elsneri sp. nov. on the grass plants. Apart from having a similar BL, minor workers of the two carpenter ants and G. elsneri sp. nov. (mean BL adults 4.43 mm, max. BL 5.3 mm) shared an obovate abdomen, a weakly shiny, blackish integument with anthracite-greyish tinge, and white and brassy/goldish setae on the abdomen ( Fig. 7B, D View FIGURE 7 ). The reddish parts of leg I and II of the spiders resembled the antennae and front legs of the ants. However, the brassy/goldish pubescence on the abdomen of C. cf. blandus was denser than in G. elsneri sp. nov. Pseudomyrmex termitarius is unlikely a model for G. elsneri sp. nov., as this ant species has a distinctly more elongated body with a reddish thorax and pointed abdomen. The remaining co-occurring ants were considerably smaller or larger than G. elsneri sp. nov. The larger ants ( Camponotus leydigi Forel, 1886 , Ectatomma permagnum Forel, 1908 and Odontomachus sp. ) had a more elongated body and different body color.
After approaching individuals of G. elsneri sp. nov., they ran at a comparably slow speed, with short stops and frequent changes in direction, even towards the potential threat, which agreed with the behavior observed in their potential ant models. Other co-occurring spiders escaped with fast sprints (e.g., lycosid spiders) or jumps (salticid spiders) and/or tried to hide. Individuals of G. elsneri sp. nov. proceeded with the same locomotory behavior after being placed in a vial to photograph their live habitus.




| R |
Departamento de Geologia, Universidad de Chile |
| CBF |
Coleccion Boliviana de Fauna |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Castianeirinae |
|
Genus |