
Krabisuchus siamogallicus, Martin & Lauprasert, 2010
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2009.00582.x |
|
persistent identifier |
https://treatment.plazi.org/id/03E68793-FFE5-AC73-AFB6-F8FCFE3CFB6F |
|
treatment provided by |
Valdenar |
|
scientific name |
Krabisuchus siamogallicus |
| status |
sp. nov. |
KRABISUCHUS SIAMOGALLICUS SP. NOV. ( FIGS 2–9 View Figure 2 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 View Figure 9 )
Holotype: Kr-C-012, a complete skull with associated mandibular rami.
Etymology: ‘Siam’ in reference to the ancient name of Thailand and ‘Gallicus’ in reference to the ancient name of France. The species name ‘ siamogallicus ’ refers to the Thai–Franco collaboration, whose members contributed to the collection and study of the fossil crocodiles from Krabi.
Locality and horizon: Lignite beds of the Late Eocene (Priabonian) ( Benammi et al., 2001) in Muang and Nuea Khlong districts, Krabi Province, southern Thailand.
Diagnosis: Krabisuchus siamogallicus can be united to basal Alligatorinae in presenting: downturned retroarticular process, posterior teeth bulbous, lacrimal longer than prefrontal. Krabisuchus siamogallicus also differs from all other alligatorines in having a long dorsal premaxillary process, a medially shifted quadrate foramen aëreum, and vertical ridges on the conical teeth.
Description
Preservation of the specimens: The fossils are preserved in lignite. The bones are heavily crushed along one axis and deformation more or less affects the original shape of bones. Most often, skulls are dorsoventrally flattened but may also be crushed laterally as in Kr-C-006. This skull however preserves some original volume especially on its right side. Mandibles are often associated with skulls as they are crushed underneath them. Fossils seem to preserve some degree of connection but given that crushing provokes fusion of various elements with the lignite and bone, the associations are difficult to examine with clarity. It is, therefore, common to find packs of postcranial elements fused together, where bone boundaries are sometime difficult to discern from the matrix.
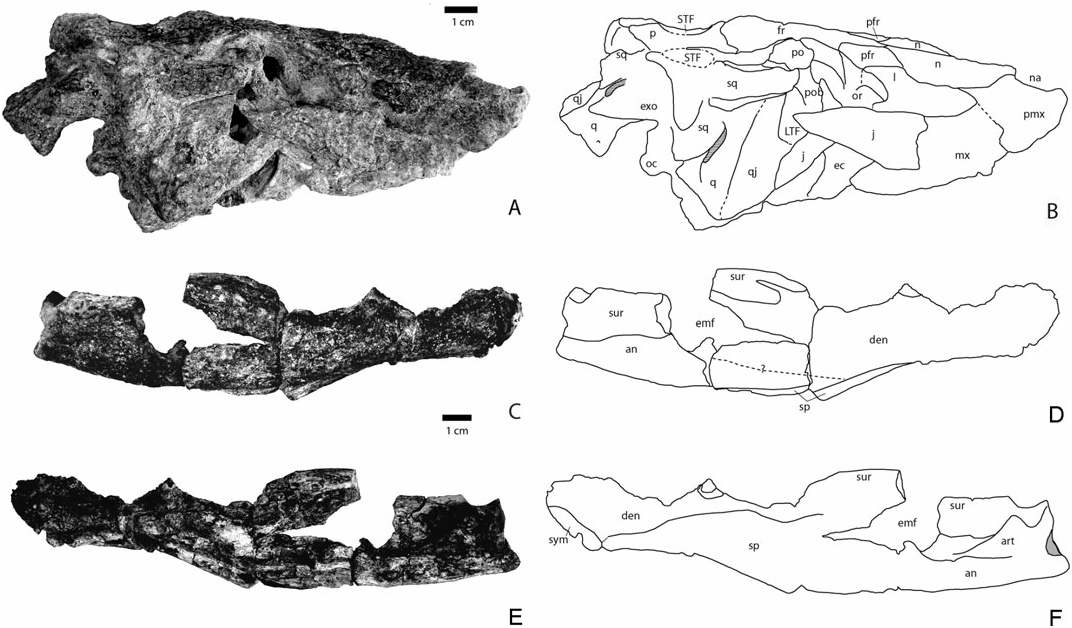
General description: Small alligatorid probably not exceeding 2 m in total length (see Tables 1 and 2 for cranial and mandibular measurements). The altirostral skull is similar to that of Arambourgia gaudryi , Procaimanoidea kayi , and Hassiacosuchus haupti . The rostrum is short, being about as long as the posterior region including the orbits. The external nares face almost anteriorly if not anterodorsally. The nasals are strongly built and draw a strong relief on top of the rostrum. The orbits are quite large but the skull table is not extensive. The mandibles are robust with a strong symphysis and well-delineated tooth waves. The posterior level of the mandibles is elevated. A notch is present between the premaxilla and maxilla. Posterior maxillary and dentary teeth are low crowned and somewhat bulbous. The bulbous condition is not as extreme as in Hassiacosuchus haupti from Messel but resembles more that of Navajosuchus mooki . Two complete mandibular rami (Kr- C-007 and 008) and the complete skull (Kr-C-006) belong to the same specimen. The external mandibular fenestra is longer than high.
Premaxilla: This bone was observed in only three specimens (Kr-C-006, 010, and 012). It is longer than large and makes up the entire lateral margin of the naris. In Kr-C-006, the right premaxilla is well preserved, whereas the left is broken. This bone is slightly prominent along the naris as shown in Kr-C-006 and 010. The external naris is anteroposteriorly elongate. Kr-C-006, which preserves the least deformed premaxilla, shows that the posterior rim of the external naris is more elevated than the anterior rim. Such a dip is not easy to observe in the other two dorsoventrally flattened specimens but in Kr-C-012 the premaxillae slope anteriorly. The external naris faces anterodorsally as in Procaimanoidea and Arambourgia gaudryi . Despite an unclear sutural pattern combined with deformation, the posterior premaxillary process is long and extends at least to the level of the third maxillary alveolus. The suture for reception of the premaxilla on the maxilla shows that the premaxilla extends posteromedially. Only Kr-C-010 provides information about tooth count. Five teeth are visible on the right premaxilla. Their alveoli cannot be observed as the teeth occlude labially over the dentary. The ventral surface of the premaxilla is visible only from the left side of Kr-C-006. The margin of the incisive foramen indicates that it was small and reached neither the anterior tooth row nor the premaxilla–maxilla suture.
Maxilla: The best preserved maxillae are also found in Kr-C-006, 010, 012, and 021. A notch is present at the premaxillary–maxillary junction for the reception of the largest mandibular tooth. However, in Kr-C-010 and 012, where mandibulae are in occlusion, the largest fourth dentary tooth is not visible laterally and might be occluding in a pit at this level. On the ventral surface (visible in Kr-C-006 and 021), occlusal pits are positioned lingually to the tooth row. The tooth row is not elevated, being approximately at the level of the palatal region. The maxilla makes up the anterior portion of the lateral suborbital fenestra. The ectopterygoid does not touch the posterior maxillary tooth row but it is not excluded by a wide extension of the maxilla. Instead, the maxilla is thinner at the contact with the ectopterygoid. In Kr-C-006, the suture with the jugal is obscured. The jugal overlaps this region. Seven alveoli are discernable in the right maxilla of Kr-C-006 but a full count is not possible because its anterior most part is missing. The best count is obtained on Kr-C-021, a right maxilla with 14 alveoli. The fourth alveolus is the largest. The tooth row is deeply festooned with a constriction behind the fifth tooth. In all specimens, the maxilla is slightly inflated dorsally at the level of the fourth maxillary tooth.
Nasals: The nasals are preserved in Kr-C-006, 010, and 012. They remain wide for most of their length, being one-third of the maximum rostrum width. They become slightly thinner posteriorly. Their contact with the maxilla is unclear, but at best the maxilla may contact the nasal in its anteriormost region. Thus the lacrimal almost or totally prevents the maxilla from contacting the nasal. Nasal and lacrimal remain in contact for at least two-thirds of the nasal length. The nasal contacts the prefrontal at its posterolateral corner. The nasals make up the top dorsal surface of the rostrum. They enter the external naris dorsally through an anteriorly tapering process. They make up all of the posterior margin of the narial opening.
Prefrontal: The prefrontal forms the anteromedial corner of the orbits. The frontal prevents the prefrontals from meeting medially as the frontal projects far anteriorly. The bone is anteroposteriorly elongate and tapers at both ends. Anteriorly, it inserts between the lacrimal and nasal. The prefrontal pillars cannot be observed.
Lacrimal: The lacrimal has a triangular shape and is best preserved in Kr-C-006, 010, and 012. The bone is very elongate and is about twice as long as the prefrontal. Its posterior region contributes to the anteriormost rim of the orbit, and the preorbital ridge is absent. Laterally, the lacrimal broadly contacts the maxilla. Its suture with the jugal is unclear.
Frontal: The anteriormost frontal process projects between the nasals. The frontal contribution to the medial orbital margin reaches half of the orbital
·
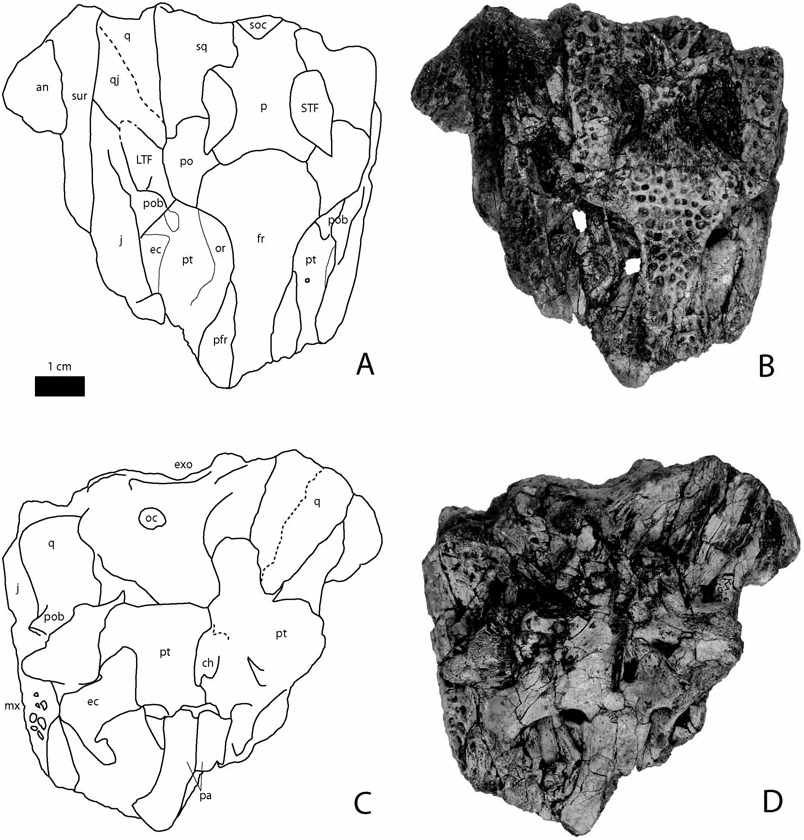
Figure 3. Images and line drawings of the holotype of Krabisuchus siamogallicus from the Late Eocene (Priabonian) of Muang and Nuea Khlong districts, Krabi Province, Thailand (Kr-C-012) in dorsal (A, B) and ventral (C, D) views. Abbreviations: an, angular; ar, articular; boc, basioccipital; den, dentary; ef, median Eustachian foramen; emf, external mandibular fenestra; exo, exoccipital; fr, frontal; j, jugal; l, lacrimal; LTF, lower temporal fenestra; mx, maxilla; n, nasal; na, naris; oc, occipital condyle; or, orbit; p, parietal; pa, palatine; pfr, prefrontal; pmx, premaxilla; po, postorbital; pob, postorbital bar; pt, pterygoid; q, quadrate; qj, quadratojugal; ra, retroarticular process; san, surangular; soc, supraoccipital; sp, splenial; sq, squamosal; STF, supratemporal fenestra; sur, surangular; sym, symphysis.
length. The frontoparietal suture is concavoconvex and is positioned at the anteriormost level of the supratemporal fenestrae. The most lateral portion of this suture enters the anteriormost portion of the fenestra. The frontal does not enter the supratemporal fenestrae as the parietal and postorbital meet along the anterior part of these openings. The dorsal surface of the frontal is continuous with the orbital margin. Although most specimens show a flat dorsal surface of the frontal that resulted from dorsoventral compression, Kr-C-006 shows that there is a continuous difference of height between the skull table and the anterior tip of the rostrum. From a lateral point of view, the frontal is continuous and slightly convex as in Arambourgia gaudryi and Procaimanoidea , reminiscent of an altirostral condition.
Postorbital: The postorbital is best represented in Kr-C-065 and makes up the posterior margin of the orbit. It extends further posteriorly than laterally. Its lateral margin flushes against the orbital frontal margin so that the anterior contour of the skull table is continuous with the orbits as in basal globidontans. The postorbital makes up all of the anterior and anterolateral margin of the almond-shaped supratemporal fenestrae. The left side of Kr-C-006 shows a suture for the jugal on the postorbital bar. The jugal seems to have reached the dorsalmost level of the bar.
Jugal: The anterior extension of the jugal contacts the lacrimal, participating in the anterolateral margin of the orbit. The jugal shows a slightly elevated rim along its dorsal margin, which disappears at the level of the postorbital bar. The postorbital bar is inset from the dorsal margin of the jugal. Kr-C-006 shows that the jugal becomes narrow and eventually turns into a rod, which makes the ventral margin of the infratemporal fenestra. The jugal meets the quadratojugal anteriorly at the posterior angle of the infratemporal fenestra. Its lateralmost extension does not project on the quadratojugal.
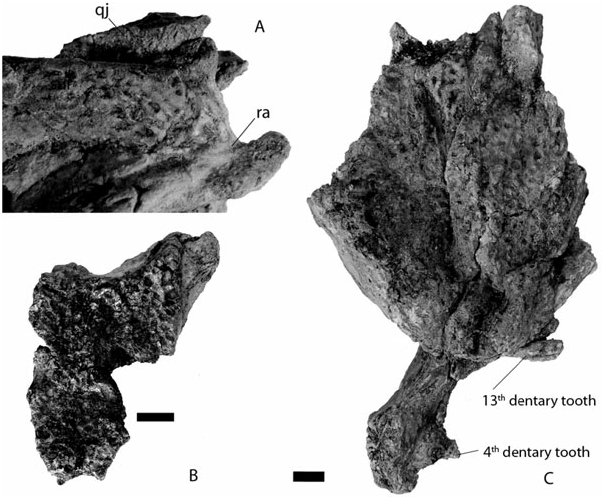
Quadratojugal: This bone can be seen well in Kr-C- 006 and 065. It is dorsally smooth and ornamented near the jugal suture, and fills the posterior angle and the dorsal margin of the infratemporal fenestra. It seems to prevent the quadrate from reaching this fenestra. The quadratojugal appears all along the lateral region of the quadrate.
Quadrate: This bone is preserved in Kr-C-006, 021, and 065. The lateral hemicondyle is the largest whereas the medial one is smaller and dorsoventrally expanded. The quadrate bears a dorsal boss for the reception of the paraoccipital process. The foramen aërum is visible close to the medial margin on the left quadrate. Because of deformation, it is difficult to assess the extension of the quadrate branches. However, Kr-C-015, 065, and the right side of 006 seem to show that the quadrates did not project far posteriorly as observed for most Alligatorinae . The ventral side of the quadrate is not visible.
Squamosal: The squamosal makes up most of the lateral and the posterior margin of the supratemporal fenestra. It is anteroposteriorly elongate and represents two-thirds of the length of the skull table. Its lateral margins are almost parallel, contrary to Arambourgia gaudryi or Procaimanoidea where the squamosal flushes medially toward the posterior level of the skull table. Here, the squamosal prongs project posteriorly but not extensively. The posterior margin of the squamosal overhangs the exoccipital.
Parietal: The parietal is slightly constricted between the supratemporal fenestrae but remains wide. Its width between the fenestrae is the same as the interorbital width. It contacts the squamosal along a straight suture near the posteromedial corner of the supratemporal fenestrae. The bone is slightly depressed at this level and appears to be flat between the openings. An elevated rim underlines the medial contour of these fenestrae. Here, the parietal does not overhang these fenestrae.
Supraoccipital: The bone clearly appears on the dorsal surface of the skull table, as in Navajosuchus mooki but unlike in Arambourgia gaudryi or Procaimanoidea kayi . The posterior skull table margin is uneven with a marked notch at the level of the squamosals. Medial to this notch, parietal and supraoccipital project posteriorly. In occipital view, the supraoccipital is thin and does not expand ventrally, as seen in the small specimen Kr-C-065. The supraoccipital is triangular.
Pterygoid: The pterygoid is best seen in Kr-C-065. The paired bone appears wider than long. The posterior margins of its wings are almost perpendicular to the anteroposterior direction. The wings are as wide as long. The posteromedial portion of the pterygoids seems to project slightly posteriorly but this is unclear. The pterygoid makes up the posteromedial corner of the suborbital fenestra. The suture with the palatine occurs anterior to the posterior limit of the fenestra. The suture with the ectopterygoid is straight. The choanae are fully enclosed in the pterygoids. They are very elongated anteroposteriorly and, although positioned closer to the posterior margin of the pterygoids, they represent half of the anteroposterior length of the pterygoids.
Ectopterygoid: The ectopterygoid makes up most of the posterior rim and half of the lateral margin of the suborbital fenestra. Sutural contact with the pterygoid indicates that the ectopterygoid does not reach the posteriormost level of the pterygoid wing but rather overlaps this bone for two-thirds of its length. The lateral edge of the suborbital fenestra is made up of the medially bowed ectopterygoid. The ectopterygoid participation on the postorbital bar is unknown but its posterior extension on the jugal bar is prominent, at least for most of the infratemporal length, as seen on the right side in Kr-C-006 where the ectopterygoid is disarticulated.
Palatines: The palatines (Kr-C-065) are short and slightly curved on the suborbital margin. They participate in all the medial and in the anterior half of the suborbital fenestra. They send a wide anterior process beyond the level of the suborbital fenestrae. The suborbital fenestra is smaller than the orbits and almost as long as wide. Its posterior margin is smooth and does not present any notches.
Dentary: The bone is represented by the following specimens: Kr-C-007 and 008, which are barely deformed and several others that are flattened: Kr-C- 011, 012, 017, 018, 022, 028, and 032. Each ramus comprises 18 alveoli (count obtained from Kr-C-032). The fourth mandibular tooth is the largest and occludes in the pit between the premaxilla and maxilla. The dentary is then characterized by a deep constriction in the tooth row, deeper than that of Navajosuchus mooki , until the 12th alveolus. The fourth maxillary tooth occludes labially in that area. All alveoli in this area are minute and about the same diameter. Their diameter suddenly increases from the 13th alveolus. The tooth row is now dorsally positioned at about the same level as the first four alveoli. Kr-C-012 and 032 show very large alveoli/teeth in this area (13 and 14, respectively). A bulbous dentition is present in Kr-C-028, but is characteristic of a more posterior position. In medial view, the symphysis is obliquely orientated and attains the level of the fifth dentary tooth. The tooth row is punctuated lingually with small foraminae. The area dorsal to the symphysis is long and slopes gently posteriorly. From a dorsal perspective, the dentary is arched laterally at the level of the 13th tooth. The anterior region is wide,
giving a general robust aspect to the mandibular branches.
Splenial: The splenial is not completely preserved but its sutural contact with the dentary in Kr-C-007 and 032 permits us to state that it contacts the posterior margin of the symphysis. Contrary to Navajosuchus mooki , it does not extend dorsal to the symphysis but rather contacts all the posteroventral margin of the symphysis. The splenial remains dorsoventrally thin until the level of the constricted region of the tooth row; it then expands greatly from the level of the 13th tooth to eventually reach the dorsalmost level of the tooth row near the posterior crushing teeth. Kr-C-022 shows that the splenial lamina is thickened mediolaterally, but not as much as in Acynodon . The posterior organization of the splenial is best seen in Kr-C-017. Its posteriormost process participates in the anterior portion of the foramen intermandibularis caudalis.
Angular: The angular participates in the ventral portion of the external mandibular fenestra. On the lateral side, the angular–dentary suture begins at the anteriormost corner of the fenestra as seen in Kr-C- 012 and 007. On the medial side, the angular prevents the splenial from participating in the ventral limit of the foramen intermandibularis caudalis by the presence of an anterior process (Kr-C-017). The angular also makes up the posterior and dorsal margin of this craniocaudally elongate foramen. The posterior part of the angular is almost horizontal. The ventral insertion for musculus pterygoideus posterior is well marked and smooth.
Surangular: The angular–surangular suture is laterally horizontal and connects to the posteriormost corner of the external mandibular fenestra. The external mandibular fenestra is oblique with its posterior corner located dorsally relative to the anterior corner. The dorsal margin of the surangular is thin and very elevated compared to the jaw joint. It gives a tall aspect to the posterior part of the mandible. Anteriorly, this portion enlarges progressively for insertion of the M. adductor mandibulae externus superficialis and medius. The anteriormost portion is forked with the medial extension longer and more massive than the lateral extension.
Articular: The articular flushes along the surangular surface. The foramen is lingually positioned on the articular medial surface. The anteriormost process of the articular does not meet the angular–surangular suture. The retroarticular process has a light dorsal curvature and mostly projects posteriorly contrary to most modern eusuchians. The retroarticular process is positioned below the level of the glenoid fossa. This condition is similar for Navajosuchus mooki , Hassiacosuchus haupti , and Arambourgia gaudryi .
Dentition: The premaxillary dentition is very fragmentary and can be described from Kr-C-010. Here, the teeth are broken but their conical shape is preserved. The first two and the fifth teeth are very small. The third and fourth are about the same size but twice as big as the other teeth. Anterior maxillary and dentary teeth are conical but their crown is not very long. Instead, it is more robust than in Alligator mississipiensis . All the crowns bear carinae. Many small vertical ridges ornament the labial and lingual surface of these teeth. The fourth maxillary tooth is the largest and is followed by minute teeth. Kr-C-066 preserves two crushing posterior teeth, which are low crowned with rounded surfaces. One of these teeth is particularly longer than wide ( Fig. 7E View Figure 7 ). The mandibular teeth follow the same conical pattern with the fourth tooth being the largest, and then the 13th and 14th teeth being as big as the fourth ( Fig. 7C View Figure 7 ). Kr-C- 028 preserves two erupting teeth with the same crushing morphology as in the maxilla, the last being longer than wide. Their position is impossible to establish.
Postcranial elements: Postcranial elements represented here are for the most part osteoderms, which often create conglomerates around all the rest of the skeleton. However, some vertebrae and single elements such as the phalanx, ilium, scapula, and femur can be described.
Several disarticulated postcranial elements (Kr-C- 020) were found associated with a poorly preserved skull (Kr-C-019) and therefore may belong to the same individual. The sculpturing on osteoderms consists of regularly spaced small and rounded pits covering their entire dorsal surface. Two types of osteoderms can be recognized. The first type is square and bears a median and prominent keel that progressively elevates in the anterior region and extends beyond the posterior margin ( Fig. 8A, B View Figure 8 ). This morphotype has a moderately concave ventral surface and may pertain to the dorsal parasagittal series. In contrast, the second type of osteoderms bear a much more prominent median keel ( Fig. 8C, D View Figure 8 ). The extent of this keel is comparable to the other dorsal osteoderms in that it extends beyond the posterior margin as well. However, their ventral surface is deeply concave and differences in keel position on the dorsal surface can be observed. Whereas in some specimens the keel lies in a median position ( Fig. 8D View Figure 8 ), in others the keel is displaced medially ( Fig. 8C View Figure 8 ). These osteoderms are reminiscent of the cervical shield because no suture between the osteoderms could be detected.
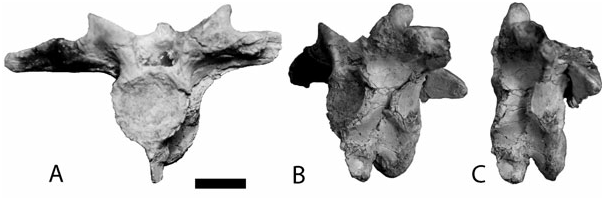
All the vertebrae are procoelous. Cervical vertebrae (Kr-C-020, 21, and 043) bear a hypapophysis near the anterior end and present elliptical parapophyses in cross-section ( Fig. 9 View Figure 9 ).
Kr-C-038 represents a left ilium ( Fig. 10 View Figure 10 ). Its posterior process is well developed and bears a notch on its dorsal margin. The dorsal part has a rough edge. The supra-acetabular crest is protruding. The anterior margin of the bone is not even but is marked by an anterior process.
Kr-C-025 is a left scapula lacking its distal end ( Fig. 10 View Figure 10 ). The deltoid crest on the posterolateral margin is sharp and thin. The anterior part connecting with the coracoid is massive, whereas it is thin and flat posteriorly.
Two femora are preserved. Kr-C-021 lacks the distal end; Kr-C-034 is complete (80 mm long). The ulna-olecranon is flat and medially expanded.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |