
Martensolasma catrina, Cruz-López, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4338.3.7 |
|
publication LSID |
lsid:zoobank.org:pub:5FC9941F-D455-48DD-8EDC-BD2278E571F6 |
|
DOI |
https://doi.org/10.5281/zenodo.6004762 |
|
persistent identifier |
https://treatment.plazi.org/id/03E687F2-947A-C337-A4CD-FE69FB01FE86 |
|
treatment provided by |
Plazi |
|
scientific name |
Martensolasma catrina |
| status |
sp. nov. |
Martensolasma catrina View in CoL sp. n.
Figs 1–24 View FIGURES 1 – 4 View FIGURES 5, 6 View FIGURES 7 – 9 View FIGURES 10 – 15 View FIGURES 16, 17 View FIGURES 18 – 24
Material examined. Male holotype CNAN-T1109 , Rancho Manzanal, Meztitlán, Hidalgo, Mexico, 20º40’30.814’’ N, 98º42’36.468’’W, 2,164 m, 04.ii.2017, J. Cruz, G. Contreras, D. Barrales, and L. Olguín colls. Two male and two female paratypes CNAN- T1111, same data as holotype. Other material examined. One male CNAN-SEM-Op0231 GoogleMaps , same locality data, 20.i.2016, J. Cruz, G. Contreras, J. Mendoza and L. Olguín colls. One female CNAN-SEM-Op0235 GoogleMaps , same data as holotype. One female CNAN-DNA-Op328 GoogleMaps , same data as CNAN-SEM- Op0231. GoogleMaps
Etymology. “Catrinas” are one of the most emblematic cultural symbols in Mexico. They represent pictures of elegant women’s skeletons wearing elaborate (upper class) clothing and were originally intended to criticize and protest against political and economic inequality in Mexico in latter half of the 19th century. Nowadays, they are important characters of one of the most representative holidays in Mexico called “Día de Muertos” (day of the Dead). The name is a singular noun in opposition.
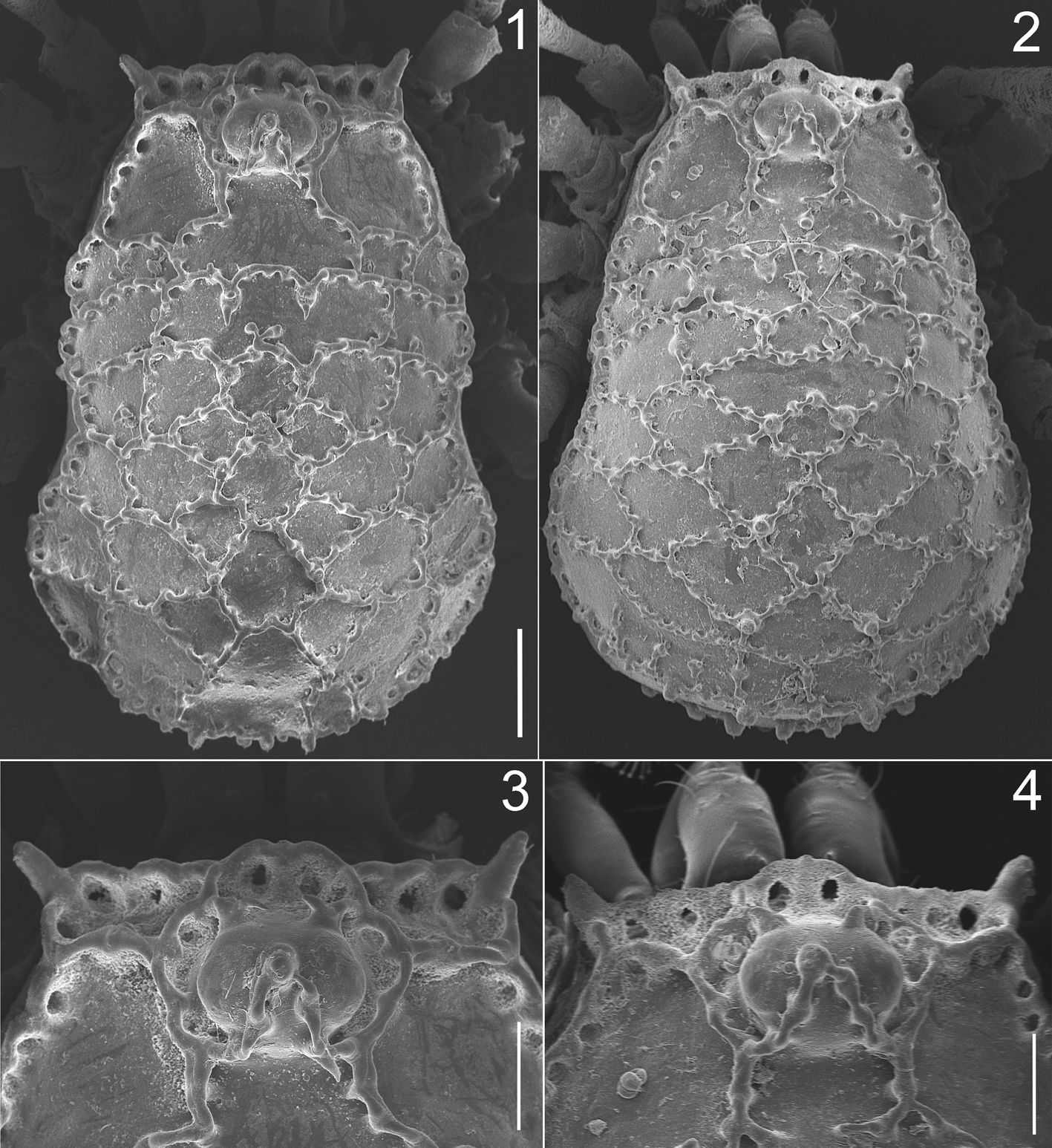
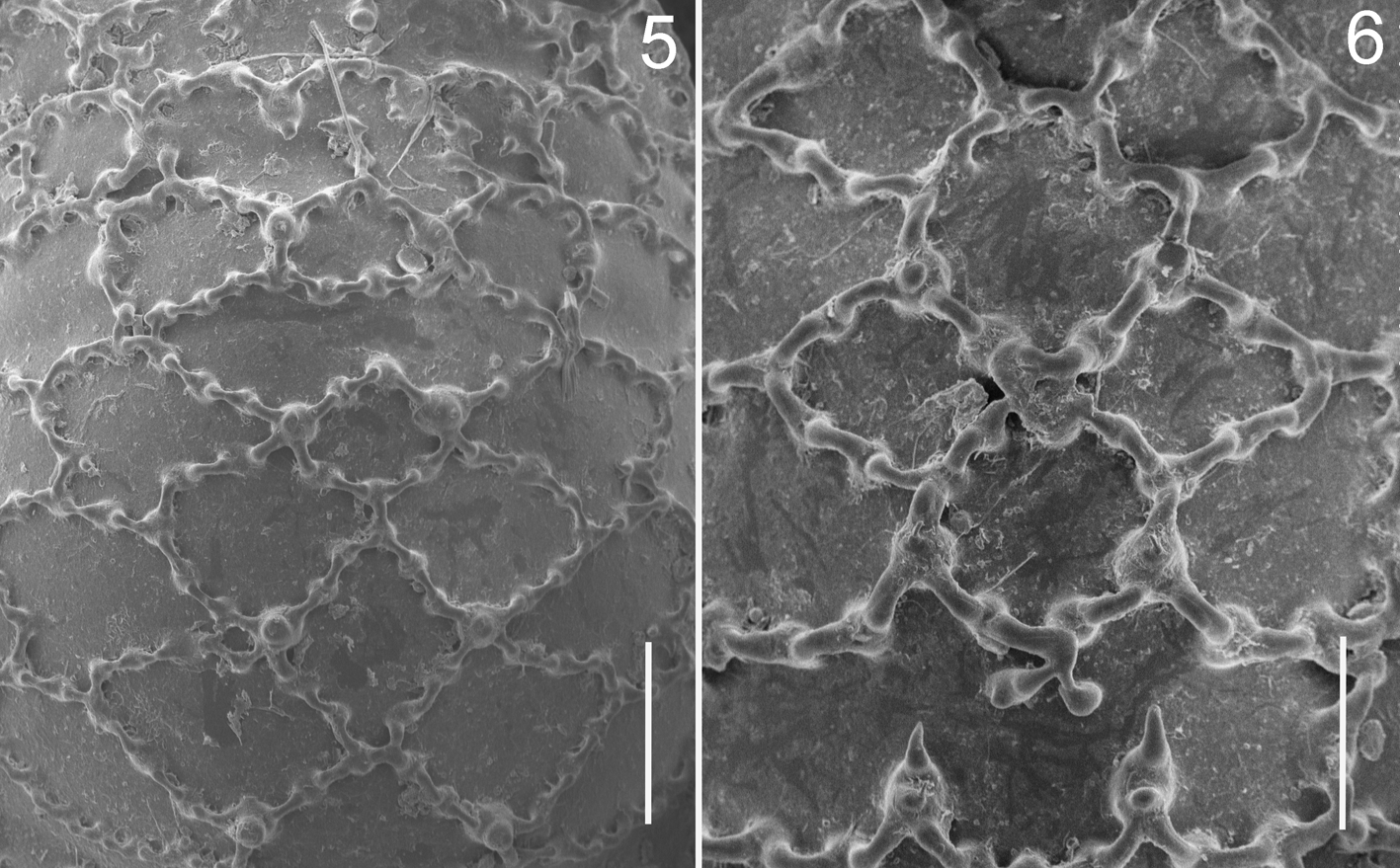
Diagnosis. Martensolasma catrina sp. n. can be differentiated easily from M. jocheni by the following combination of characters: scutal ornamentation complex—second to fifth transverse rows with five distinctly marked cells, the central three forming a forward-pointing arrowhead, cells generally closed and arranged in a rhombic pattern, whereas in M. jocheni the cells are organized in parallel rows, with most not closed ( Figs. 5, 6 View FIGURES 5, 6 ); basal segment of chelicera with small curved mesal tooth, sometimes absent ( Figs. 7–10 View FIGURES 7 – 9 View FIGURES 10 – 15 ); metatarsus II with subapical pseudoarticulation; three articles in tarsus II; penial stylus hook-shaped at tip ( Figs. 19–24 View FIGURES 18 – 24 ).
Description. Male holotYpe: Scute length = 1.80 mm. Anterolateral processes of scutum short, with rounded apices. Dorsal cells defined as follows: Two large cells lateral to eye tubercle, cell posterior to eye tubercles incomplete, continuous with the median cell of the first transverse row. First row with five cells, the two lateroexternal pairs small, with median longitudinal keel not well defined; central cell larger. Second row with five cells, latero-external cells large, rectangular; three median cells subequal in size, with longitudinal keels incomplete, three median cells arranged in a W-shaped array. Third to fifth rows with five cells, each row with latero-external pairs slightly larger than the median three; the median three cells forming an arrowhead, with central cell pentagonal, displaced anteriorly. Sixth row with five subequal cells, central cell triangular, with acute anterior apex; a small cell is between the central and the first lateral-right cells ( Figs. 1, 3 View FIGURES 1 – 4 , 5 View FIGURES 5, 6 ).
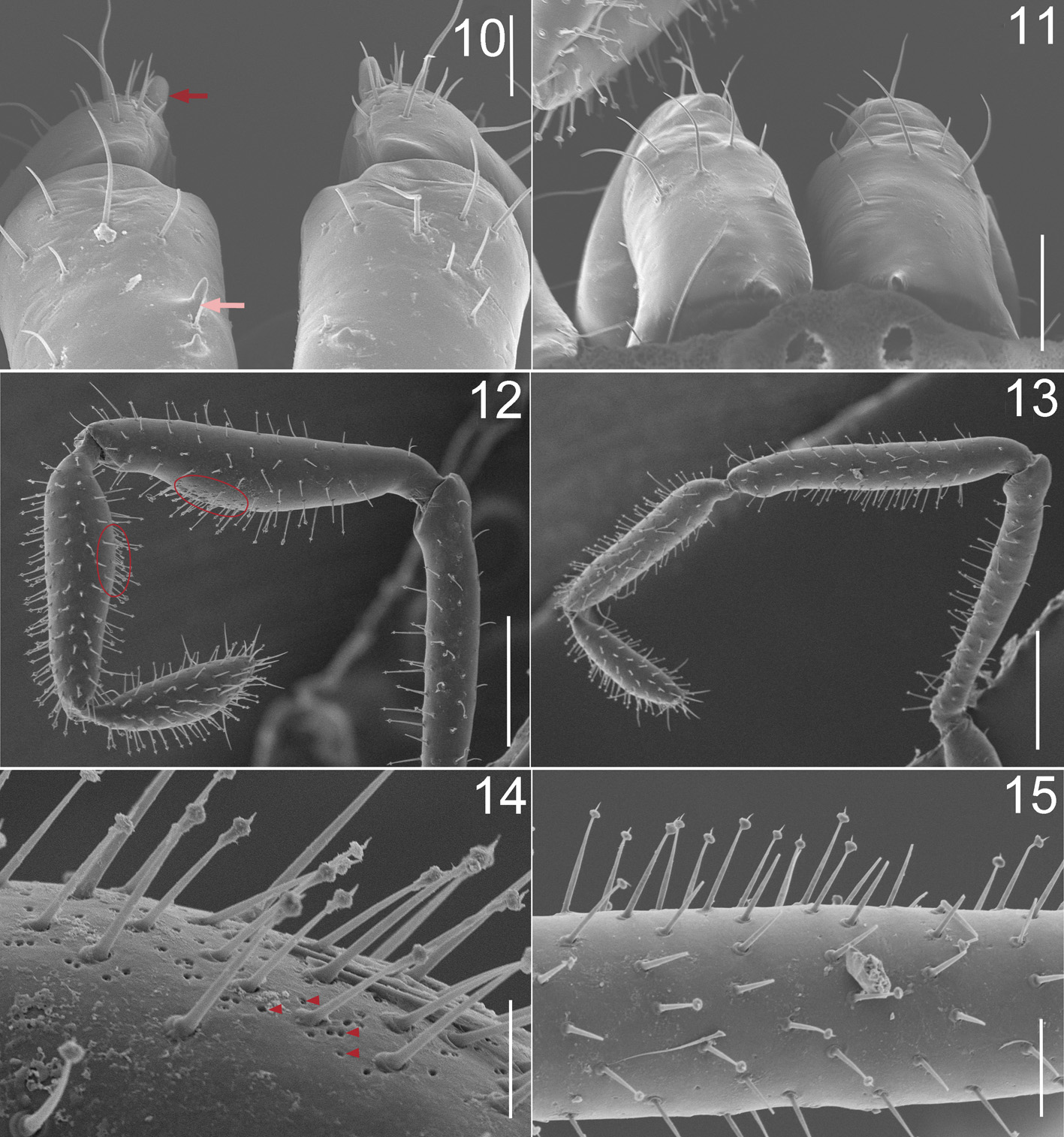
Chelicera small, with basal segment smooth, humped dorsally, with a small curved mesal tooth. Second segment with small, curved mesoproximal tooth, without evidence of pore field ( Figs. 7–10 View FIGURES 7 – 9 View FIGURES 10 – 15 ).
Pedipalpal segments thin. Femur with a few scattered glandular setae, distributed along entire segment. Patella swollen meso-apically, covered with few scattered glandular setae, with dense aggregation of small glandular pores on swollen surface. Tibia covered with many glandular setae, swollen meso-basally; small glandular pores on translucent swollen surface. Tarsus rounded apically, covered with many glandular setae and a few ordinary setae ( Figs. 12, 14 View FIGURES 10 – 15 ).
Leg formula: 2, 4, 3, 1 (measurements in table 1). Femora cylindrical in cross section; middle of femora I and III slightly swollen, femur IV with basal pseudoarticulation. Patellae globular. Tibiae shorter than femora, slightly swollen. Metatarsi long and slender, metatarsus II with subapical pseudoarticulation. Metatarsi covered densely with short trichomes and longer fine setae, more prominent in metatarsus II ( Fig. 16 View FIGURES 16, 17 ). Tarsi I and II with three articles, the third with a basal pseudoarticulation, giving appearance of four articles. Tarsi III and IV with four articles, the third and fourth with pseudoarticulations.
Penis long and thin. Stylus narrow, tip noticeably hook-shaped, apex rounded. Monomorphic setation, no more than seven small setae ( Figs. 18–24 View FIGURES 18 – 24 ).
Female paratYpe: Scute length = 1.96. Similar to the male, but with the following differences: Sulci on mesotergum slightly developed, tending toward scutum parvum; median scutal cells of second to fifth rows complex, irregular in shape; second segment of chelicera without mesoproximal tooth; pedipalpal patella and tibia not swollen, without glandular pores ( Figs. 2, 4 View FIGURES 1 – 4 , 6 View FIGURES 5, 6 , 11, 13, 15 View FIGURES 10 – 15 , 17 View FIGURES 16, 17 ).
Natural history. All specimens were found under small rocks in the boundary area between a cattle ranch and a pine forest. The specimens were found in sympatry with an undescribed species of Trilasma Goodnight & Goodnight, 1942 and the stygnopsid Chapulobunus unispinosus Goodnight & Goodnight, 1946 and undetermined species of the genera Crettaros Cruz-López & Francke, 2015 , Karos Goodnight & Goodnight, 1944 , and Potosa Goodnight & Goodnight, 1947 .
Discussion. Shear (2006) discussed the phylogenetic affinities of Martensolasma and the remaining Ortholasmatinae , and suggested that the unusual absence of the modified eye process, the dorsal ornamentation without complex cells, and the paucity or absence of pseudoarticulations on the femora and metatarsi could represent plesiomorphic characters within the subfamily. Shear (2006, 2010) considered Martensolasma to be the most divergent member of Ortholasmatinae . With the recent discovery of M. catrina described here, the morphological features of the genus are more heterogeneous, with those of the new species being shared with other members of the subfamily. According to the morphological-phylogenetic hypothesis of Shear & Gruber (1983), the dorsal ornamentation with well-marked cells and hooked stylus are synapomorphies for the clade Ortholasma + Trilasma . However, these characters are also present in M. catrina , although with the dorsal cells not as well defined as in Trilasma . Thus, M. catrina presents an intermediate condition between Martensolasma and Trilasma . This observation opens the question of whether Martensolasma is a member of the Ortholasma + Trilasma clade based on genital synapomorphies (and with a reversal in M. jocheni ), or whether the hooked stylus appeared independently in Ortholasma + Trilasma and M. catrina . However, it is possible that the stylus of the type specimen of M. jocheni depicted by Shear (2006: fig. 14) was broken and that the hooked terminal portion was lost. If so, this would indicate that the hooked stylus is ancestral for Martensolasma and that the “straight stylus” of M. jocheni is an artifact. Indeed, the specimens of this species available to me all have hooked styli.
Morphological variation among ortholasmatines is noted only in the number of pseudoarticulations on the metatarsi and number of tarsomeres of all legs. Dorsal ornamentation, shape and ornamentation of the ocular process, and the number of lateral processes are constant among species. On the other hand, male genital morphology is conserved, being very similar and apparently without variation among the genera ( Shear & Gruber 1983, 2010). The presence of straight and hooked styli and large and mixed dorsal cells in the same genus, opens the possibility that morphology has evolved in a different way than has been proposed. A future phylogeny of Ortholasmatinae might illuminate the evolutionary morphology of taxonomic traits in this group.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |