
Acrocanthosaurus atokensis Stovall & Langston, 1950
|
publication ID |
https://doi.org/10.5281/zenodo.4664675 |
|
persistent identifier |
https://treatment.plazi.org/id/03E6E25D-E129-E12C-FCAF-D2DE1F02FA00 |
|
treatment provided by |
Felipe |
|
scientific name |
Acrocanthosaurus atokensis Stovall & Langston, 1950 |
| status |
|
Acrocanthosaurus atokensis Stovall & Langston, 1950
MATERIAL. — North Carolina State Museum of Natural Sciences, NCSM 14345, nearly complete skull and most of postcranium.
LOCALITY AND AGE. — NCSM 14345 was recovered from the Antlers Formation (Trinity Group, Lower Cretaceous) of McCurtain County, Oklahoma.
D E F G
FIG. 3. — Acrocanthosaurus atokensis . Reconstruction of skull; A, dorsal view; B, left lateral view; C, occipital condyle; D-G, denticles of maxillary teeth. Scale bars: A, B, 10 cm; C, 2 cm.
REVISED DIAGNOSIS. — Large theropod with elongate neural spines that are more than 2.5 times corresponding presacral, sacral and proximal caudal lengths of the centra. Lacrimal contacts postorbital; supraoccipital expands on either side of the midline to protrude as a double boss behind the nuchal crest; pleurocoelous fossae and foramina pronounced on all presacral and sacral vertebrae; cervical neural spines have triangular anterior processes that insert into depressions beneath overhanging processes on preceding neural spines; accessory transverse processes on mid-caudal vertebrae.
DESCRIPTION
NCSM 14345 was found lying on its side, partially disarticulated. There has been some crush-
ing and distortion of the skull, mostly towards the back ( Fig. 2 View FIG ). For example, the left postorbital was broken in at least two places, and had pulled away from its suture with the frontal, the ventral part rotating medially. Minor crushing and distortion are also evident in the posterior half of the jaws.
The skull ( Figs 2 View FIG ; 3) is almost 129 cm long (premaxilla to quadrate) with a preorbital length of 85 cm. The height of the skull in front of the orbit is 47 cm. Femur length ( Table 1) is a relatively stable standard of measurement in theropods, and can be used to assess proportional differences of the skull. The skull/femur ratio is 1.17, which compares well with Ceratosaurus Marsh, 1884 (1.00, Gilmore 1920), sinraptorids (0.92-1.17, Currie & Zhao 1993) and tyrannosaurids (0.97-1.17). The skull of Allosaurus is apparently relatively short ( 0.76 in USNM 4734 and 1.03 in AMNH 5753).
In addition to the five major openings on each side of the skull (external naris, antorbital fenestra, orbit, upper and lateral temporal fenestrae), there are two accessory openings in the maxilla ( Figs 2 View FIG ; 3). The external naris is relatively small and elongate, with a maximum length of 16 cm (anteroventral to posterodorsal axis). The naris is bound entirely by the premaxilla and nasal, and the maxilla is excluded from the opening. This is the plesiomorphic state found in primitive theropods like Herrerasaurus Reig, 1963 (Sereno & Novas 1992), in contrast with Allosaurus Marsh, 1877 (Madsen 1976) , Monolophosaurus Zhao & Currie, 1993 and some Cretaceous theropods (Currie 1985). The triangular antorbital fenestra is enormous, covering almost half ( 42 cm) the preorbital skull length and two thirds (63%, 29 cm) of the height. The jugal forms part of the boundary in Acrocanthosaurus . In some reconstructions of Allosaurus (Madsen 1976) , the jugal is excluded from the margin of the fenestra, although it does reach the antorbital fenestra in at least some specimens (Bakker, pers. comm. 1993). The keyhole shaped orbit is high ( 34 cm) but relatively short anteroposteriorly ( 12 cm). The outline of the lateral temporal opening is pear-shaped and is bordered above by a short intertemporal bar.
Anterior to the orbit, the side of the skull is a relatively flat, vertical face, clearly set off from the narrow skull roof as in most carnosaurs. A sharply defined ridge separates the dorsal and lateral surfaces of the nasal and lacrimal bones. The ridge, which overhangs the face by as much as 1.5 cm at the anteriormost nasal-lacrimal contact, is relatively straight in profile and is only slightly rugose as in Allosaurus (Madsen 1976) .There are no nasal, lacrimal, frontal or postorbital horns as there are in Allosaurus , Ceratosaurus , Carnotaurus Bonaparte, Novas & Coria, 1990 , Monolo- phosaurus and most tyrannosaurs, and in this character Acrocanthosaurus is similar to Tyrannosaurus . The paired premaxillae form the sharp, narrow tip of the skull (Fig. 3A). Each is pierced by more than half a dozen foramina (Fig. 3B) for branches of the medial ethmoidal nerve and subnarial artery. The maxillary body is higher than long as in Ceratosaurus , Yangchuanosaurus Dong, Zhou & Zhang, 1983 and Torvosaurus Galton & Jensen, 1979 , whereas it is longer than high in Allosaurus (Britt 1991) and Sinraptor Currie & Zhao, 1993 . In contrast with Allosaurus , the anterior margin slopes posterodorsally. There is a sloping, posteriorly concave (in lateral aspect) contact with the maxilla, which tapers into a posterodorsal (subnarial) process to contact the subnarial process of the nasal. As in Allosaurus , the subnarial foramen passes between the main bodies of the premaxilla and maxilla.
The maxilla is more than 82 cm long. It is excluded from the border of the external naris by the premaxilla and nasal, but forms most of the anterior margin of the antorbital opening. A maxillary fenestra ( 7 cm by 3.5 cm) pierces the maxilla between the external naris and the antorbital opening. As in most theropods, the smaller promaxillary fenestra ( 3.5 cm high) is tucked under the rim that forms the anteroventral corner of the antorbital fossa. The right maxilla has 15 teeth (counting one empty position), which is close to the counts for Sinraptor (15, Currie & Zhao 1993) and Yangchuanosaurus (14-15, Dong et al. 1983). As in Allosaurus , Sinraptor , Yangchuanosaurus and most other large theropods, the lateral surface of the maxilla is rugose only along its anterior edge and immediately above the tooth row. It is definitely not rugose in the way that the maxillae of Abelisaurus , Carcharodontosaurus , Carnotaurus and Giganotosaurus are. Most of the lateral surface of the maxilla is smooth and somewhat indented where it defines the extent of the antorbital fossa. The maxillae of Carcharodontosaurus (Stromer 1931; Sereno et al. 1996), Carnotaurus (Bonaparte et al. 1990) , Ceratosaurus (Gilmore 1920) , Giganotosaurus (MUCPv-CH-1), Indosuchus Chatterjee, 1978 and Torvosaurus (Britt 1991) are quite different in that most of the lateral surfaces are rugose, showing that the antorbital fossae are not as well developed.
The ventral margin of the promaxillary fenestra in the anteroventral apex of the antorbital fossa is at about the same level as the floors of the naris and the antorbital fenestra. This fenestra is found in most theropods; sometimes (e.g. Ceratosaurus ) it is not visible in lateral aspect, and sometimes it is a relatively large opening (Witmer 1997). The larger maxillary fenestra is not consistently expressed in theropods, but tends to be present in advanced forms, including Afrovenator (Sereno et al. 1994) , Allosaurus (Witmer 1997) , tyrannosaurids and most small theropods. It is absent in Abelisaurus Bonaparte & Novas 1985 , Carnotaurus (Bonaparte et al. 1990) , Ceratosaurus (Gilmore 1920) , Carcharodontosaurus Stromer, 1931 (Sereno et al. 1996 identify a maxillary fenestra, but it is in the position normally occupied by the promaxillary fenestra), Giganotosaurus (MUCPv-CH-1), Indosuchus Chatterjee 1978 , Majungatholus Sues & Taquet, 1979 (Sampson et al. 1998) and Torvosaurus (Britt 1991) , and is expressed as a variable complex of openings in sinraptorids (Currie & Zhao 1993; Witmer 1997). The posterodorsal process of the maxilla passes between the nasal bone and the antorbital fenestra, and bifurcates distally to embrace the anteroventral process of the lacrimal.
The nasal is 85 cm in length, more than four times the length of the frontal. The dorsal surface is flat and relatively narrow for most of its length, and is clearly delimited by a sharp inflection from the nasal contribution to the antorbital fossa as it is in Allosaurus , Sinraptor and Yangchuanosaurus . This is different in abelisaurids (Bonaparte & Novas 1985; Bonaparte et al. 1990), Monolophosaurus and tyrannosaurids, where at least half of the nasal has a convex and highly rugose dorsal surface. Like most theropods,the median internasal suture is not fused. The nasal is split anteriorly where the posterior tip of the premaxilla overlaps a shallow trough for a length of 11 cm. As in Allosaurus , Ceratosaurus , Giganotosaurus , Monolophosaurus , Sinraptor and Yangchuanosaurus , the nasal forms the anterodorsal rim of the antorbital fossa, and even extends slightly onto the medial wall. It appears as if pneumatic openings may have invaded the nasal along the margin of the antorbital fossa in the way they do in Allosaurus (Madsen 1976; Gilmore 1920), Giganotosaurus (R. Coria, pers. comm. 1996), Monolophosaurus (Zhao & Currie 1993) and Sinraptor (Currie & Zhao 1993) . As in most other large theropods, there is a groove in the ventrolateral margin of the nasal for the maxillary articulation that would have permitted some lateral-medial rotation of the maxilla. Above the posterior end of the suture with the maxilla, the nasal overlaps the anterior tip of the lacrimal. Each nasal meets a prefrontal posterolaterally, and overlaps an anteroposteriorly elongate shelf of the frontal (Fig. 3A).
The lacrimal is 35 cm long and 37 cm high. The dorsolateral margin forms a low ridge at the front of the orbit comparable with Carcharodontosaurus , Giganotosaurus (R. Coria, pers. comm. 1996), Sinraptor (Currie & Zhao 1993) and Yangchuanosaurus (Dong et al. 1978) . Allosaurus , Ceratosaurus (Gilmore 1920) , the smaller genera of tyrannosaurids, and many other theropods have developed “horns” in this region.The lacrimal has a limited dorsal exposure. Anteriorly, there are interdigitating sutures with the maxilla and nasal. Its lateral surface is sculptured posterodorsally for close association with the skin. The lacrimal sends a narrow posterior extension along the ventrolateral margin of the prefrontal and seems to contact the postorbital beneath a thin lateral lappet of the prefrontal (Fig. 3B). In lateral aspect ( Figs 2A View FIG ; 3B), the posterior margin of the ventral (preorbital) process of the lacrimal has a double concavity separated by a short process (for attachment of the Ligamentum suborbitale) that marks the anteroventral extent of the eyeball itself (diameter was about 11 cm). Although less obvious in the holotype (Stovall & Langston 1950), the process is similar to those of Abelisaurus (Bonaparte & Novas 1985) , Majungatholus (Sampson et al. 1998) , Monolophosaurus , Sinraptor and Yangchuanosaurus . The posterior margin of the preorbital bar of other large theropods tends to be uniformly concave in lateral view. Below this low process of the lacrimal in Acrocanthosaurus , the preorbital bar narrows to 64 mm in lateral aspect. The posteroventral end of the lacrimal expands somewhat for its contact with the jugal. Much of the anterolateral surface of the preorbital bar is shallowly excavated for the antorbital fossa, which extends ventrally onto the jugal. Ventrally, the lacrimal is overlapped by the jugal, and does not contact the maxilla. In contrast, the lacrimal may contact the maxilla externally in Allosaurus and Ceratosaurus (Gilmore 1920) .
As in other large theropods, there is a lateral, pneumatic opening into a vacuity in the main posterodorsal body of the lacrimal. The opening has a diameter of 2.7 cm in this specimen. It would have housed a pneumatic diverticulum of the nasal cavity (Osmolska 1985; Witmer 1997). The presence of two large openings in the lacrimal of the holotype (Stovall & Langston 1950) shows there is some variation of this feature, which is not surprising considering how variable pneumatic features are (Currie & Zhao 1993).
Posteromedial to the lacrimal, there is a relatively large, triangular prefrontal ( 11 cm long, 6 cm broad) as in Allosaurus , Sinraptor and other carnosaurs. It extends posteriorly to contact the postorbital and helps to roof over the orbit. A thin lappet appears to have extended over the lacrimal to reach the orbital margin.
The dorsal surface of the frontal is flat, as it is in Allosaurus and Sinraptor (Currie & Zhao 1993) . The surface of the bone is smooth as in the majority of large theropods, but in contrast with the ornamented surface in Abelisaurus (Bonaparte & Novas 1985) and Majungatholus (Sampson et al. 1998) . As in the holotype, the interfrontal suture is more or less completely fused. Like the frontals of the holotype, there is a slightly raised rugose bump along the midline near the frontoparietal suture. A shallow trough, emphasized anteriorly by a raised ridge, leads into a foramen at the junction of the frontal, prefrontal and postorbital (Fig. 3B), although the latter two bones contact each other to exclude the frontal from the margin of the orbital rim. To an extent, this is a size specific characteristic that is shared by the largest theropods, including Carcharodontosaurus , Giganotosaurus , Tarbosaurus and Tyrannosaurus .
The frontal forms the anterior rim of the supratemporal fossa, which it floors for a distance of almost 5 cm. At the dorsal limit of the upper temporal fenestra, the fused frontoparietal suture is strengthened by additional bone deposition that protrudes laterally into the fenestra as a sharply defined ridge. This has also been noted in Giganotosaurus (Coria & Currie in prep.) and Sinraptor (Currie & Zhao 1993) . The frontoparietal suture can be followed laterally to the point where both bones contact the postorbital.
The parietal has limited dorsal exposure along the midline (about 3 cm) and compares well with the parietals of carcharodontosaurids and sinraptorids. However, the dorsal surface is almost perpendicular to the occiput (Stovall & Langston 1950: pl. II-2), which itself is almost vertical. This angle is distinctly obtuse in carcharodontosaurids and sinraptorids. Correlated with this, each posterolateral wing of the parietal forms an almost vertical posterior wall for the supratemporal fossa, whereas it strongly slopes in carcharodontosaurids and sinraptorids. The posterolateral wing extended along the top of the paroccipital process, tapering to a point that ends only 5 cm from the distal end of the process. There is no sharp sagittal crest along the midline of the parietal and, in this sense, Acrocanthosaurus is as primitive as all theropods except tyrannosaurids and troodontids. The nuchal crest is low and does not extend more than a few millimetres above the supraoccipital. Again, this situation is very different in tyrannosaurids and troodontids. The orbital margin of the postorbital extends forward to contact the lacrimal and prefrontal, and has thickened into a prominent horizontal orbital boss as in abelisaurids, carcharodontosaurids and the largest tyrannosaurids. These are size-dependent characteristics that have uncertain taxonomic significance. The rugose rim of the postorbital is 4 cm thick dorsoventrally. The intertemporal ramus of the postorbital is relatively short and is oriented almost as much ventrally as it is posteriorly (Fig. 3B). The postorbital bar of the postor- bital is relatively wide in lateral view, reaching a maximum anteroposterior length of 9.7 cm behind the orbit and has an anterior process beneath the orbit ( Fig. 2A View FIG ). This process is also evident in the holotype (Stovall & Langston 1950), and similar processes are found in Carcharodontosaurus , Giganotosaurus , Gorgosaurus Lambe, 1917 , Tarbosaurus Maleev, 1974 and Tyrannosaurus . Presence of the process is in part size-related, and therefore has limited utility for assessing taxonomic relationships.
The jugal is similar to those of most other large theropods in that it separates the maxilla and lacrimal to take part in the rim of the antorbital fenestra. The long, sloping anteroventral edge sits in a shallow trough on the posterodorsal margin of the maxilla. The lateral surface is depressed anteriorly where it contributes to the antorbital fossa. This region has been damaged somewhat,but there were almost certainly pneumatic openings into the body of the jugal as in other specimens of Acrocanthosaurus (Stovall & Langston 1950; Harris 1998a), some specimens of Allosaurus (USNM 4734, UUVP 1403, UUVP 3894, UUVP 3981), Monolophosaurus , Sinraptor and tyrannosaurids.The postorbital process of the jugal (NCSM 14345, OMNH 10146) is a huge triangular plate of bone in lateral view, and has a long sloping contact with the postorbital bone that only reaches two thirds of the way up the postorbital bar. The lateral surface of this part of the jugal is shallowly concave as in Daspletosaurus Russell, 1970 , Tarbosaurus and Tyrannosaurus . Similar to A l l o s a u r u s, C e r a t o s a u r u s, S i n r a p t o r and Deinonychus Ostrom, 1969 , the jugal forms the ventroposterior margin of the orbit. Like all theropods except Sinraptor dongi , the quadratojugal process of the jugal splits posteriorly into two. The uppermost prong of the quadratojugal process is much shorter than the ventral prong as in all large theropods except abelisaurids (Bonaparte et al. 1990; Sampson et al. 1998), Ceratosaurus and torvosaurs (Bakker et al. 1992).
The holotype of Acrocanthosaurus (Stovall & Langston 1950) has an inferior process on the squamosal that expands moderately ventrally for its contact with the quadratojugal. The two bones then invade the lateral temporal fenestra to a greater extent than is seen in Allosaurus and sinraptorids, but not as much as in tyrannosaurids. The inferior process of the squamosal is indented by a deep quadrate cotylus, and the head of the quadrate would have been exposed laterally. Posteroventrally, the squamosal extended along the dorsal surface of the paroccipital process, covering up the ventrolateral process of the parietal and wrapping around onto the anterodorsal surface of the distal end of the paroccipital process. The squamosal was inclined in such a manner that it would have roofed over the posterior temporal musculature.
A conspicuous midline ridge on the supraoccipital is comparable with those of carcharodontosaurids, sinraptorids and Allosaurus (Madsen 1976) . Dorsally the supraoccipital expands on either side of the midline until it protrudes in a double boss 5 cm behind the nuchal crest. The dorsal surfaces of these knobs are rugose, and are only a few millimetres below the highest point on the parietal. Stovall & Langston (1950) reported that the supraoccipital was excluded from the margin of the foramen magnum by the exoccipitals, although fusion makes it difficult to be sure that this is the case. The lateral margins of the supraoccipital are nearly vertical, and are more similar to those of Allosaurus (Madsen 1976) than to Sinraptor .
The paroccipital process is relatively wide but shallow, in contrast with the deeper tyrannosaurid condition. Distally the process ends in a vertical sheet of bone 2.5 cm thick and 11 cm high. The paroccipital process turns downwards distally (Stovall & Langston 1950) as in Allosaurus and Sinraptor , and does not project as strongly backwards (Fig. 3A) as it does in Carcharodontosaurus (Sereno et al. 1996) , Giganotosaurus (MUCPv-CH-1) and Sinraptor (Currie & Zhao 1993) . In the holotype, at least two branches of the twelfth cranial nerve emerge through a depressed area between the occipital condyle and the paroccipital process.
The quadrate is a relatively tall bone ( 31 cm) as in almost all theropods except Allosaurus and tyrannosaurids, and is only shallowly concave posteriorly. The condyles of the jaw articulation are at least 11 cm across. There is a quadrate foramen with a diameter of 28 mm. Except for a narrow slit, it is completely surrounded by the quadrate as in Allosaurus (Madsen 1976) and Sinraptor (Currie & Zhao 1993) . The gap is closed by the quadratojugal, which also curves onto the occiput where it overlaps the quadrate somewhat along the dorsolateral margin of the foramen.
The occipital condyle (Fig. 3C) is nearly circular in appearance, which contrasts with the flattened oval appearance of the occipital condyles of Giganotosaurus , Piveteausaurus Taquet & Welles, 1977 and Tyrannosaurus . The shape of the occipital condyle is characteristic of theropods that carry their skulls at the end of an extended neck. In spite of the large size of the specimen, the sutures between the exoccipital and basioccipital are still visible in the occipital condyle.
Palatal elements are generally obscured by the other bones of the skull. The palatine and ectopterygoid of Acrocanthosaurus have been described for SMU 74646 (Harris 1998a).
There are four premaxillary, 15 maxillary and an unknown number of dentary tooth positions. The third premaxillary tooth is the largest on the left side, attaining a dorsoventral length of 85 mm and a basal diameter of 27 mm. It is 72% the length of the largest maxillary tooth. Allosaurus and Neovenator Hutt, Martill & Barker, 1996 have five premaxillary teeth, Ceratosaurus and Torvosaurus (BYU 4882) have three, and most other theropods, including abelisaurids, Eustreptospondylus Walker, 1964 , Herrerasaurus , Marshosaurus Madsen, 1976 , Monolophosaurus , Proceratosaurus Woodward 1910 , Sinraptor , tyrannosaurs and Yangchuanosaurus , have four. Because the premaxillae meet at an acute angle, the premaxillary teeth are not D-shaped in section as they are in tyrannosaurids (Currie et al. 1990). In fact, even in the first premaxillary tooth of Acrocanthosaurus , the anterior carina is positioned on the midline of the tooth at the tip.However, it curves onto the anteromedial surface closer to the root as in allosaurids and dromaeosaurids, but in contrast with Torvosaurus (Britt 1991) . Ceratosaurus has no anterior carina on premaxillary teeth (Bakker, pers. comm. 1993). There are 15 denticles per 5 mm on the anterior carina, and 13 on the posterior. The longest crown (fourth from the front) in the left maxilla is 118 mm high, with a FABL (fore-aft base length) of 41 mm. This is comparable in length with the longest tooth of a similar sized tyrannosaurid (GI 107/2, Tarbosaurus , has a skull length of 122 cm and a maximum maxillary tooth length of 115 mm), with the exception of Tyrannosaurus which seems to have had more elongate teeth (AMNH 5027 has a skull length of 136 cm and a maxillary tooth length of 142 mm). Although the maxillary teeth are narrower and more blade-like than those of tyrannosaurids, they are not as laterally compressed as the teeth of carcharodontosaurids (Sereno et al. 1996). For example, the ratio of length to width of a cross-section of the base of an Acrocanthosaurus tooth described by Harris (1998a) is 0.63, compared with 0.43 in a selection of carcharodontosaurid teeth (casts in Tyrrell Museum of privately owned specimens from Morocco). Furthermore, the teeth of Acrocanthosaurus do not have the wrinkles in the enamel that are characteristic of carcharodontosaurid teeth (Sereno et al. 1996; Harris 1998a). In spite of the large size of the teeth in NCSM 14345, the denticles are minute (the second premaxillary tooth has 12.5 per 5 mm on the posterior carina; the fourth maxillary tooth has 17.5 per 5 mm on the anterior carina, and from 12.5 to 17.5 on the posterior edge). This is even less than the denticle size of Carcharodontosaurus , where there are 10 serrations per 5 mm in a tooth with a FABL of 36 mm (Russell 1996). Harris (1998a) pointed out that the denticles extend across the tips of Acrocanthosaurus teeth, and suggested this is unusual. However, it is the same situation in a diverse assemblage of theropods that includes carcharodontosaurids, velociraptorine dromaeosaurids, and tyrannosaurids, so this feature has limited taxonomic utility.
Lower jaw
The lower jaw ( 132.5 cm long) is well-preserved on both sides, although crushing has damaged some of the thinner bones, especially along the intramandibular articulation and around the margins of the external mandibular fenestra. The external mandibular fenestra is preserved on both sides, although it is difficult to precisely define the margins. It appears to have been about 20 cm long, which is significantly larger than that of Allosaurus (Madsen 1976) . It is positioned at the junction of the dentary, angular and surangular. The jaw is shallow anteriorly, and deep posteriorly. The depth has been emphasized somewhat by collapse of the adductor fossa.
As in almost all theropods (Bakker et al. 1988), including Herrerasaurus (Sereno & Novas 1992) , there is an intramandibular joint. A posteriorly projecting lateral process of the dentary near the dorsal margin, referred to by Currie & Zhao (1993) as the intramandibular process of the dentary, fits into a slot in the labial side of the surangular. An anteriorly projecting tongue, the intramandibular process of the surangular, extends ventrolateral to the intramandibular process of the dentary to form a combination sliding and pivoting joint. Ventrally, the anterior end of the angular had a sliding joint along the top of a shelf formed by the dentary and the splenial.
The dentary is relatively long and low, with generally concave upper and lower margins in lateral view. It is unusual in having a distinctive ventral process at the bottom of the symphysis, which gives the front of the jaw ( 10 cm high) an angular, squared off appearance. This is even more conspicuous in Giganotosaurus , but is also present to a lesser degree in large tyrannosaurids, suggesting that it may be a size-dependent character. The lateral surface, especially at the front of the bone, is pierced by a dorsal row of relatively large mental foramina for innervation and vascularization of the skin and lips. The smooth surface of the bone contrasts with the ornamented surface in Abelisaurus (Bonaparte & Novas 1985) . The front of the jaw is relatively shallow and is only 96 mm deep at mid-length. For comparison, the same measurement in AMNH 5881, a specimen of Tyrannosaurus rex with a slightly shorter jaw, is 170 mm. Posteriorly, the dentary is deep ( 19 cm at the front of the intramandibular articulation, and almost 10 cm deeper at the back). Its posteri- or margin slopes posteroventrally from the lock- ing intramandibular joint. Two notches in the back of the dentary on the lower, left side are probably the result of postdepositional damage, but are fortuitous in that they expose part of the sliding joint with the angular.
The splenial presumably wrapped around the ventromedial margin of the angular, thereby forming part of the sliding intramandibular joint. However, it does not wrap around far enough to be exposed in lateral view as in Herrerasaurus (Sereno & Novas 1992) , Ceratosaurus (USNM4735),dromaeosaurids (Currie 1995) and other primitive theropods.
The anterior surangular foramen of the surangular is continuous anteriorly with a groove as in Allosaurus , Monolophosaurus , Sinraptor and other theropods. The lower margin of the groove is formed anteriorly by a finger-like process that extends laterally over the labial surface of the dentary. A similar finger-like process of the dentary contacts the medial and dorsal margins of the groove. There is an extensive squamose suture for the angular, and posterolaterally the surangular covers part of the prearticular. Stovall & Langston (1950) suggested that the posterior surangular foramen of OMNH 10146 was relatively large, as it is in tyrannosaurs. However, it is relatively small (diameter of 12 mm) in NCSM 14345, where this region is better preserved. It is found anterolateral to the glenoid beneath a powerful lateral ridge that overhangs the lateral surface of the jaw by about 5cm.This ridge is more powerfully developed than in any other known large theropod other than Giganotosaurus (MUCPv-CH-1), although it is also pronounced in abelisaurids (MACN- CH 894). Harris (1998a) noted the presence of a conspicuous knob on the surangular shelf close to the jaw articulation. As in other theropods, the surangular of Acrocanthosaurus forms part of the lateral mandibular glenoid (Harris 1998a).
The angular is a shallowly curved plate of bone strengthened by a thick ventral margin. An extensive part of the bone is overlapped laterally by the dentary, but the contact is smooth, and sliding movement was possible. Posteriorly, the angular laterally overlaps the surangular. The ventral edge of the angular is thickest where it forms the ventral margin of the jaw behind the intramandibular joint. Unlike Allosaurus but like most other theropods, the posterior end of the angular terminates anterior to the posterior surangular foramen. The dorsal edge of the supradentary can be seen behind the last mandibular tooth extending pos- teriorly across the intramandibular joint. It cannot be determined whether it was fused to the coronoid or not, although it was presumably capable of restricted sliding movement against the surangular laterally and the prearticular medially as in Monolophosaurus (Zhao & Currie 1993) . The posteroventral margin of the prearticular is exposed in lateral view where it extends to the back of the jaw.
The medial glenoid and most of the interglenoid ridge for the jaw articulation are found on the articular. Harris (1998a) reported that the interglenoid ridge of SMU 74646 is relatively low compared with that of Allosaurus . Overall, the articular looks relatively short in lateral view because of the posteromedial orientation of the retroarticular process, but is in fact much larger than it is in Allosaurus (Gilmore 1920; Madsen 1976) and tyrannosaurids (Lambe 1917; Osborn 1912; Molnar 1991).
Axial skeleton
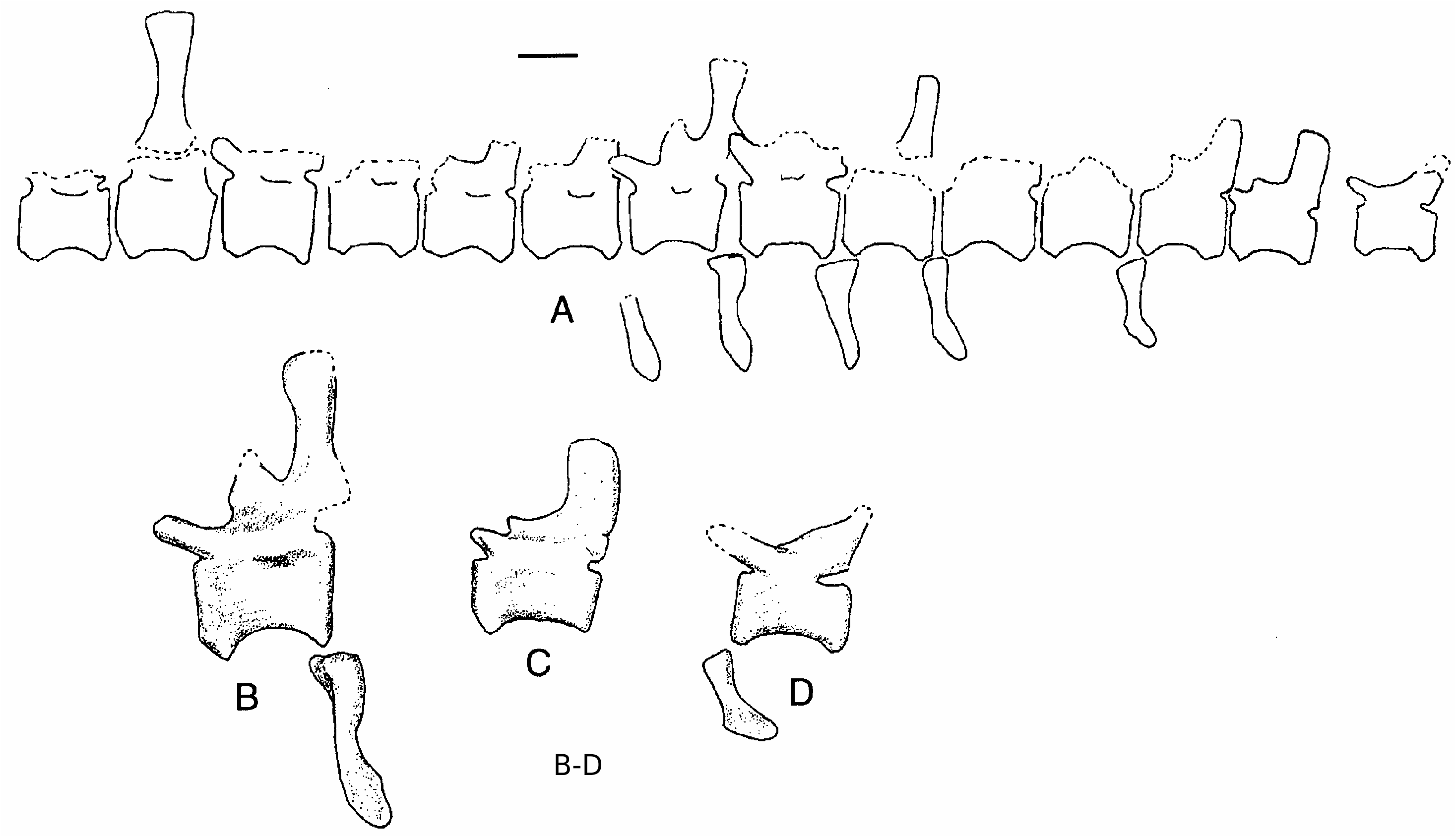
Almost two dozen partial vertebrae were recovered from the skeleton, but, with the exception of three caudals, none are complete ( Fig.4 View FIG ). From what can be seen of the vertebrae and a number of isolated neural spines, vertebrae of NCSM 14345 did not differ in any substantial way from those found with the holotype (Stovall & Langston 1950) and SMU 74646 (Harris 1998a). A fragment of a posterior cervical or anterior dorsal centrum is pierced by two pneumatopores as in OMNH 10146 and SMU 74646. The internal pneumatization is of the complex type (Britt 1993), in contrast with the simple pneumatization found in Allosaurus and Sinraptor . This has also been noted in SMU 74646 (Harris 1998a). The neural spines in the middle and distal parts of the tail are restricted to the posterior region of the vertebrae. As in the caudal vertebrae described by Stovall & Langston (1950) and Harris (1998a), there is a “cranial process” above the anterior insertion region for the interspinous ligaments ( Fig. 4B, C View FIG ). Mid-caudal centra are about 16 cm long, 10 cm wide and 11.5 cm high, whereas the most distal one preserved is 12 by 7.5 by 7.5 cm. The distal caudal has a moderately elongate prezygopophysis ( Fig. 4D View FIG ).
Six chevrons ( Fig. 4 View FIG ) from the mid-caudals are directly comparable in overall shape with those of Allosaurus (Madsen 1976) , and in spite of elongation of the neural spines, do not appear to be any more elongate than in the Jurassic theropod.
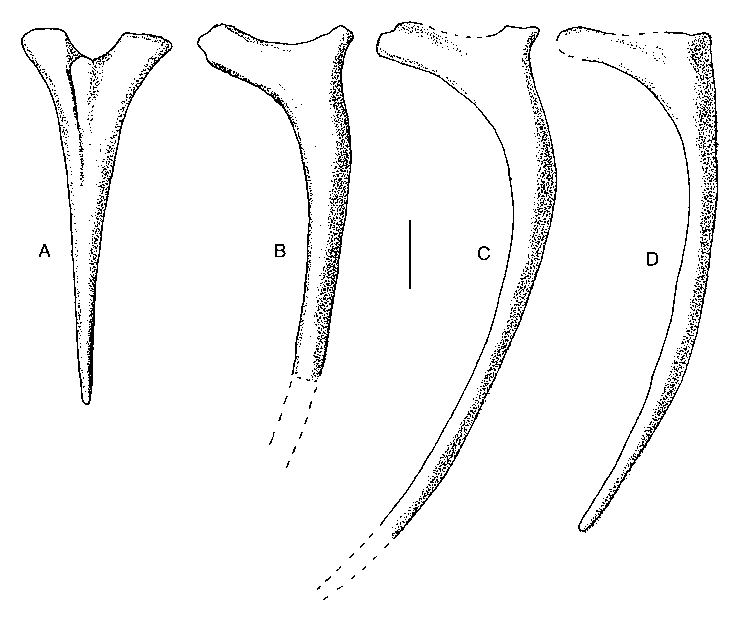
One elongate cervical rib ( Fig. 5A View FIG ) has a prominent knob projecting anterolaterally from between the tuberculum and capitulum. Most of the ribs from the dorsal region are represented in the collection, but fewer than half a dozen are complete. They are directly comparable with those of Allosaurus (Madsen 1976) , and show that Acrocanthosaurus had a rather high, narrow body. There were also many fragmentary gastralia recovered, but there is nothing to suggest that the gastral basket was different in any substantial way from other specimens of Acrocanthosaurus (Harris 1998a) .
Appendicular skeleton
All elements of the pectoral girdle ( Fig. 6 View FIG ) and front limb ( Fig. 7 View FIG ) are represented except for some of the unguals. The arm is relatively short. The sum of the lengths of the humerus, ulna (excluding the olecranon process), ulnare, metacarpal II and digit II is 105 cm, which is shorter than either the scapulocoracoid ( 118 cm) or the femur ( 128 cm). In contrast, Allosaurus has a relatively longer arm. That of MOR 693 is 94 cm long, which is 30% longer than the scapulocoracoid. The arm of Acrocanthosaurus is 81% the length of the femur, whereas the arm of Allosaurus is 26% longer than the femur in MOR 693 and 12% longer in USNM 4734. The differences are less evident in the humerus than they are in the forearm and hand, where specimens of Allosaurus (MOR 693, TATE 11, USNM 4734) that are 20% smaller than Acrocanthosaurus have radii, ulnae and metacarpals that are absolutely longer than the same elements in Acrocanthosaurus . This shows that the differences in arm length between the taxa cannot be attributed to allometric scaling. Tyrannosaurids, on the other hand, have even shorter arms than Acrocanthosaurus . The humerus is about 30% the length of the femur in both Acrocanthosaurus and Tyrannosaurus , but the forearm is relatively short- er in the latter. The front limb of Tarbosaurus (GI 107/2) is 65 cm in length, which is 93% the length of the scapulocoracoid and 60% the length of the femur, whereas that of one Tyrannosaurus rex (FMNH PR2081) is 89 cm long, which is less than 70% of either scapulocoracoid or femur length.
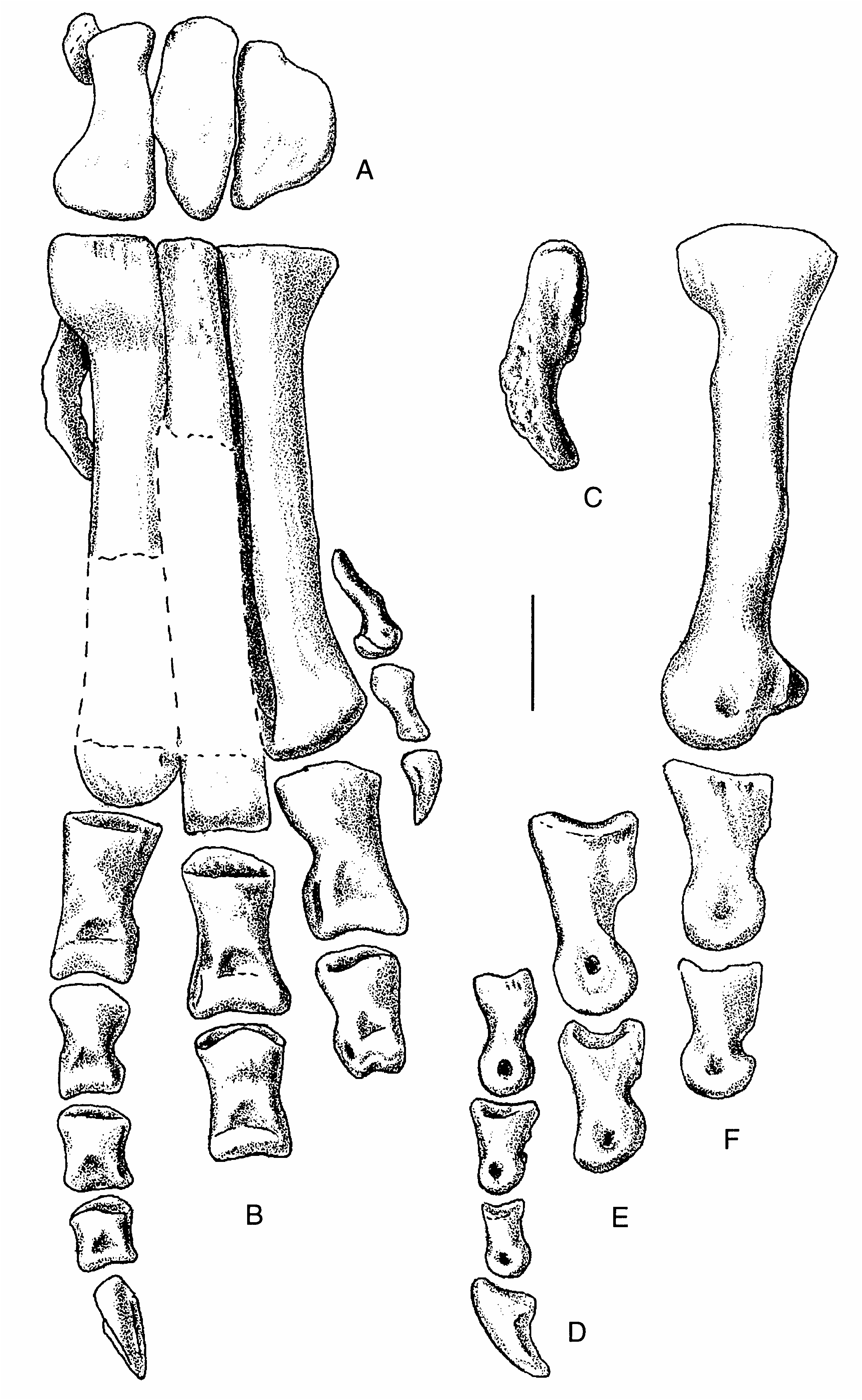
The scapula is a long, slender, gently curved element ( Fig. 6 View FIG ), 97 cm ( 103 cm measured on the outside curve) long. It is only 9.5 cm wide at its narrowest point when seen in lateral aspect, and the distal end is only moderately expanded. The acromial process is pronounced and is sharply offset from the anterodorsal margin of the scapular blade as in Allosaurus (Madsen 1976) , Sinraptor (Currie & Zhao 1993) and tyrannosaurids (Maleev 1974). The scapular blades of Ceratosaurus (Madsen 1976) , Carnotaurus (Bonaparte et al. 1990) , Edmarka Bakker et al., 1992 and Torvosaurus (Bakker et al. 1992) , and Megalosaurus Buckland, 1824 (Walker 1964) are more robust, and the anterior margin grades smoothly into the acromion process.
The outline of the coracoid ( Fig. 6 View FIG ) is also closer to that of Allosaurus than it is to Ceratosaurus (Madsen 1976) or megalosaurs (Bakker et al. 1992). The coracoid foramen is a small but conspicuous opening close to the centre of the bone. The coracoid extends posteroventrally well beyond its contribution to the glenoid as in advanced theropods plus abelisaurids.
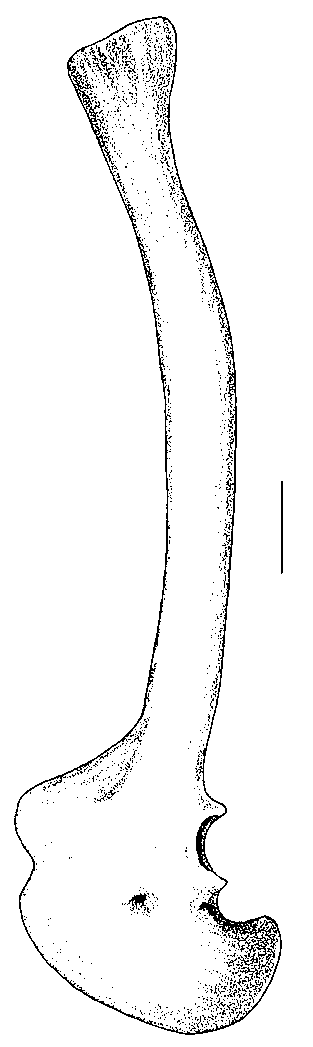
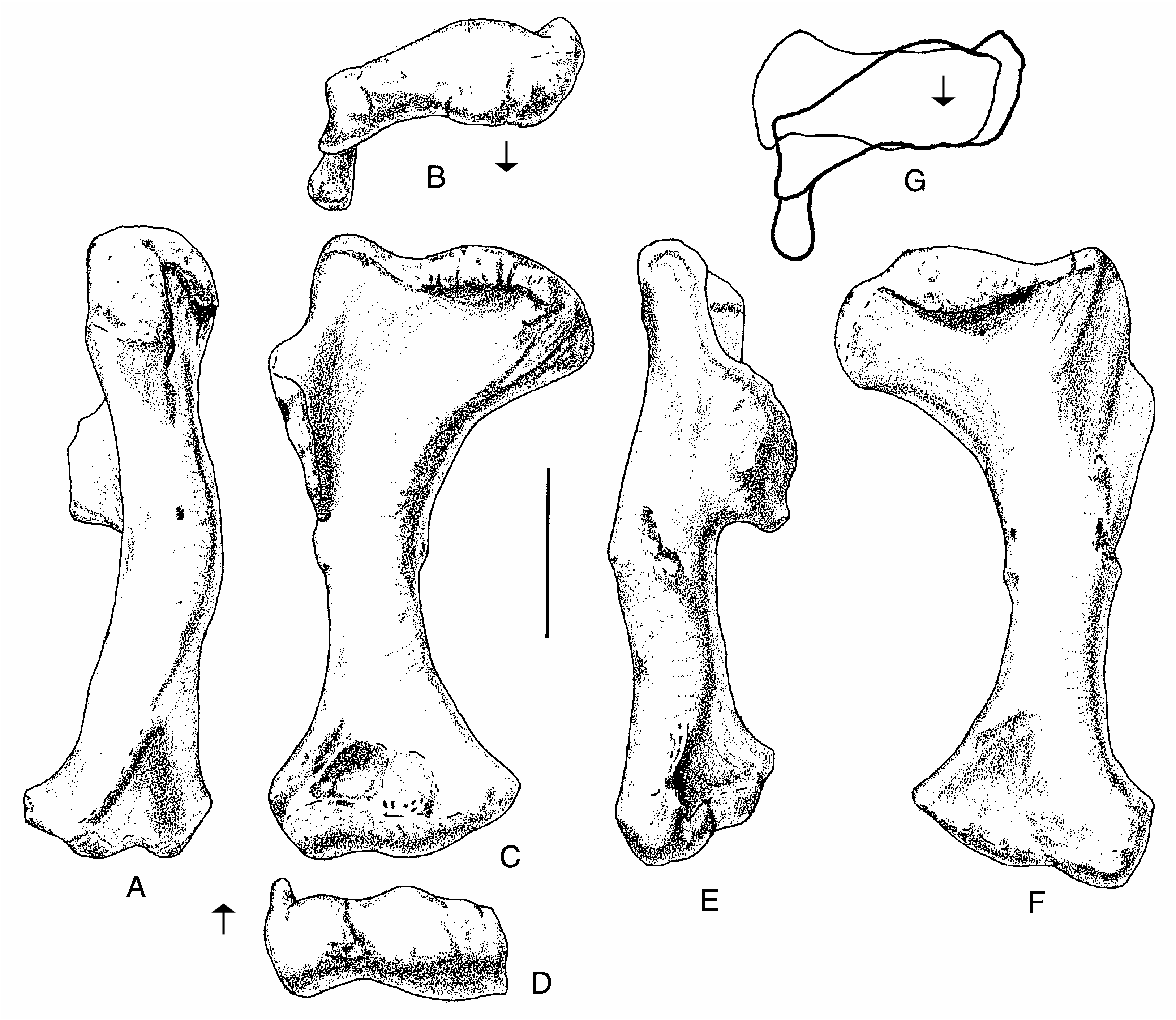
The humerus ( Fig. 8 View FIG ) is a rather massive element 37 cm long. The proximal end is expanded to such a degree that its maximum width is half the length of the bone. The deltopectoral crest is large ( 9.5 cm) and well-developed, protruding 6.5 cm from the shaft ( Fig. 8E View FIG ). Unlike Allosaurus , which has a deltopectoral crest that tapers steadily away from the shaft, that of Acrocanthosaurus is quadrilateral in outline. There is a well-developed scar for the humeroradialis muscle ( Fig. 8E View FIG ). The transverse shaft diameter is only about a third the width of the proximal end, and slightly less than half of the distal expansion. The distal end is rotated outward from the plane of the proximal end ( Fig. 8B View FIG ), and the deltopectoral crest is oriented at right angles to the distal end of the bone ( Fig. 8G View FIG ) as in Allosaurus . There is a deep pit on the anteroventral side proximal to the radial condyle and the ulnar groove (Fig. 3C). The entepicondyle is a small ( 1.5 cm) but pronounced tubercle ( Fig. 8D View FIG ) separated from the radial condyle by a groove of finished bone, although finished bone continues from the distal end of the humerus up along the crest of the tubercle. In most characters, the humerus of Acrocanthosaurus is like that of Allosaurus , and most of the differences seem to be allometric changes related to the larger size of the Cretaceous form. However, the shaft is not as twisted along its length as it is in Allosaurus .
The ulna is a robust bone with a total length of 25.5 cm, almost 15% of which is composed of the olecranon process ( Fig. 9 View FIG ). The proximal concavity that contacts the proximal end of the radius has neither articular facets nor strong ligamentous attachments. However, there is a welldefined distal articulation for the radius, which consists of a convex surface of unfinished bone elevated almost a centimetre from the general contour of the ulna by a pedestal of bone ( Fig. 9C View FIG , lower right). Amongst described theropods, the ulna most closely resembles that of Allosaurus (Madsen 1976) , although it is relatively shorter (69% the length of the humerus, compared with 85% the length in USNM 4734). The radial notch of the ulna is not as deep or prominent as in Allosaurus .
The radius ( Fig. 9 View FIG ) is a curved, slender element that is difficult to measure. The straight line length is 22 cm, whereas the distance between the closest points of the articular surfaces is only 20 cm. It is therefore 55% to 60% of the length of the humerus, whereas the radius of Allosaurus is more than 70% of humerus length (Gilmore 1920). The proximal articulation is trapezoidal ( Fig. 9H View FIG ) with a maximum mediolateral width of 6 cm and an anteroposterior length of 5.2 cm. It is saddle-shaped with strong anterior and posterior ridges. The trough-like articulation continues medially onto the ulna. The edge of the proximal end is rugose where it contacts the ulna. Distal to the proximal articulation, the shaft of the radius curves anteromedially ( Fig. 9I View FIG ) as it twists around the front of the ulna, closely following the curvature of that bone. A well-defined, twisting ridge with a rugose surface ( Fig. 9I View FIG ) lies close to the anterior margin of the ulna and presumably marks the line of ligamentous attachments with its neighbor. The lower half of the bone has an almost straight shaft, although the distal end is set off at an angle to it ( Fig. 9I, K View FIG ). There is an extensive elevated area (about 4 cm wide and 2 cm high) posteromedially near the distal articulation for contact with the ulna. Its rugose surface, and that of the corresponding region of the ulna, suggests that the ligamentous attachments were strong and movement between the bones was limited. This also would have restricted rotation of the wrist. The distal articular surface is a shallow concavity that would have articulated with the radiale.
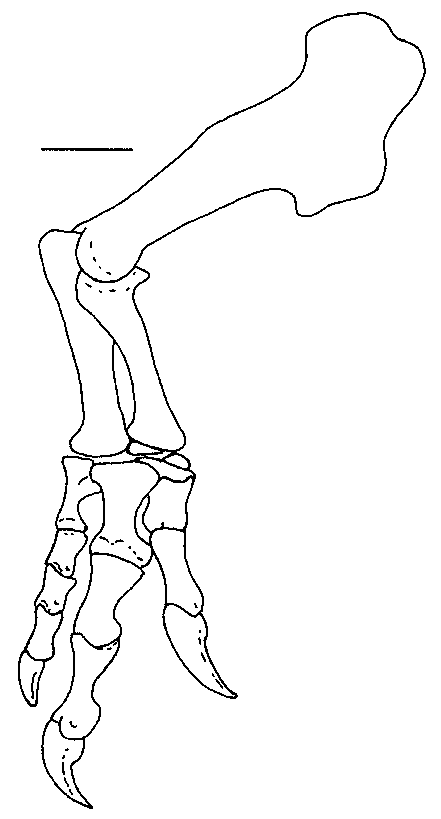
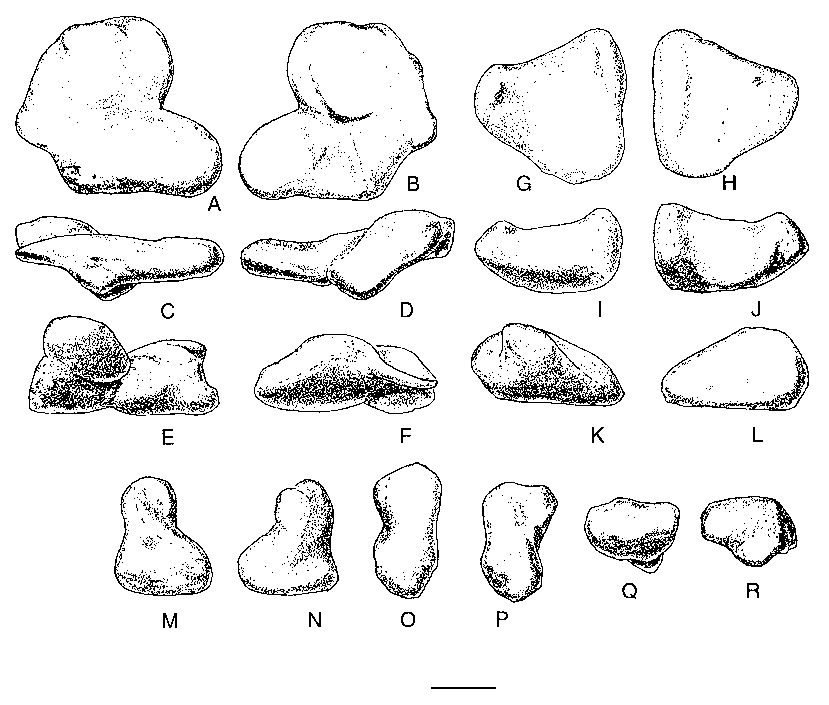
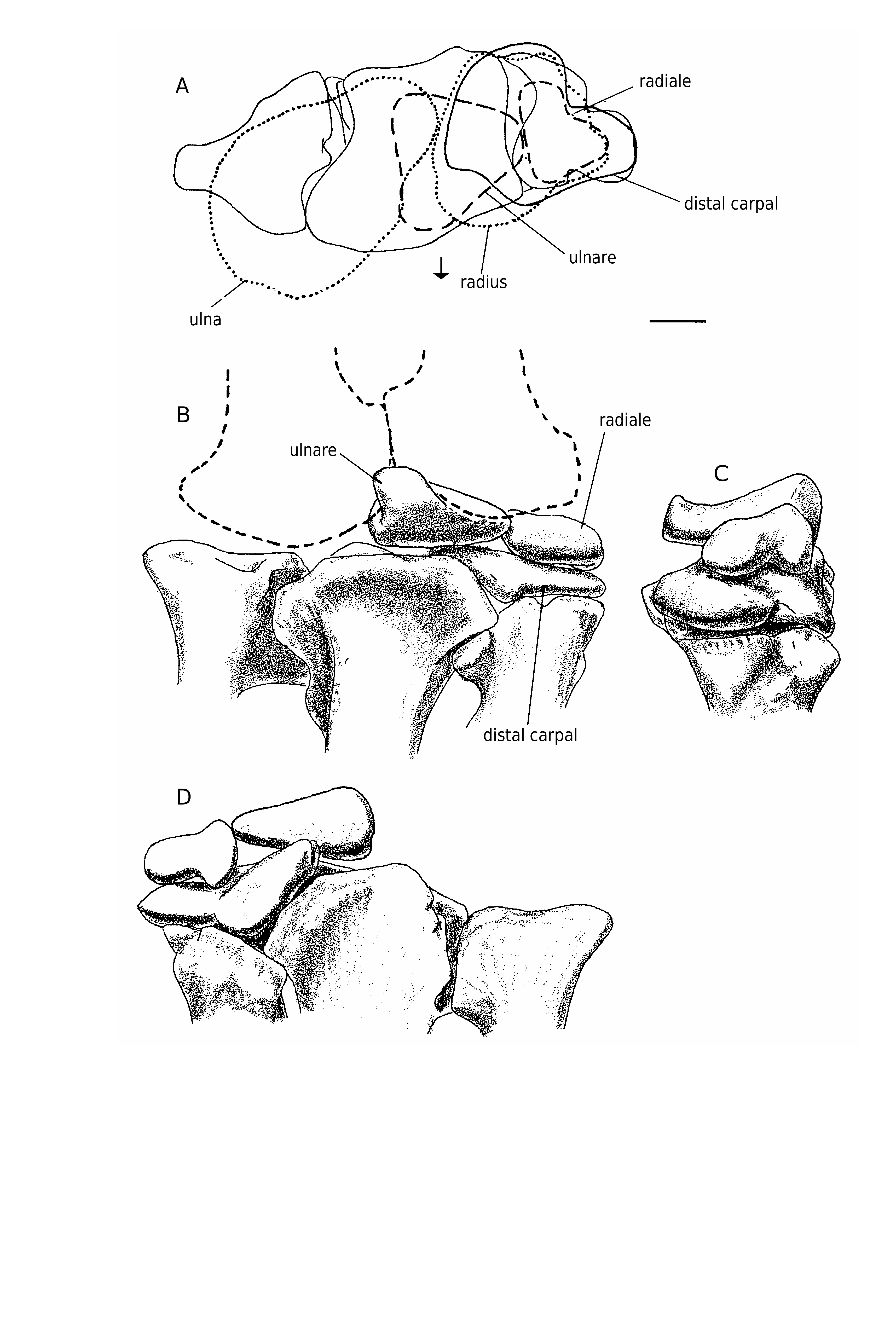
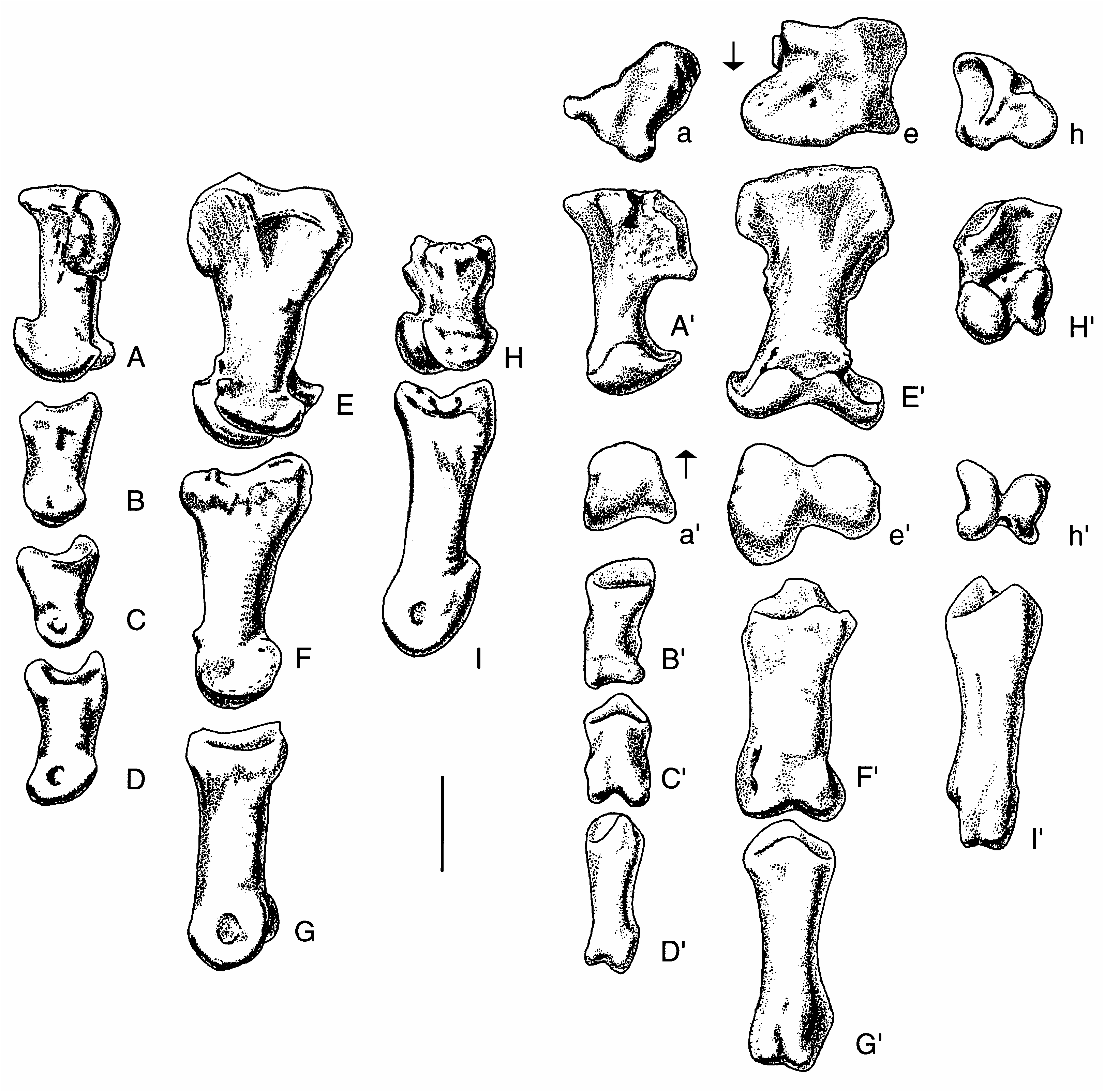
Three carpals were recovered with the specimen ( Figs 10 View FIG ; 11 View FIG ). One of these elements was preserved in articulation with the metacarpals of the right hand. Furthermore, it fits perfectly across the head of metacarpal I and extends about a quarter of the way across the head of metacarpal II. This element has been identified as the intermedium in other theropods, including Allosaurus (Gilmore 1920) , Gorgosaurus (Lambe 1917) and Tarbosaurus (Maleev 1974) , although Madsen (1976) identified it as distal carpal I (with carpal II fused into it in 14 out of 18 specimens of Allosaurus ), as did Osborn (1917) for Struthiomimus, Russell & Dong (1993) for Alxasaurus and Padian & Chiappe (1997) for maniraptorans. Colbert identified the carpal in this position in Coelophysis Cope, 1889 as distal carpal I + II. In this paper, we refer to it as distal carpal I because it occupies the primitive position (Sereno 1993) of this element above the contact between metacarpals I and II. However, we also recognize that it was probably formed by the fusion of several elements as in Allosaurus . Regardless of what name should be put on this element, it is morphologically similar to the same unit in Allosaurus , therizinosaurids and tyrannosaurids.
Distal carpal I is a complex element that is closely appressed to metacarpals I and II. This close association shows that little movement would have been possible amongst these three elements, all of which were tightly integrated. At its thickest point above the posterior junction of metacarpals I and II, it is 16.2 mm thick proximodistally. Lateromedially it measures 60.8 mm close to the extensor surface and is 52.4 mm anteroposteriorly. In comparison, one of the larger distal carpals from the Cleveland-Lloyd collection of Allosaurus elements is thicker ( 24 mm), narrower ( 50 mm at the extensor surface) and anteroposteriorly longer ( 54 mm). The anterior (extensor) surface ( Figs 10C View FIG ; 11B View FIG ) is a thin edge of finished bone (it is tall in Allosaurus, Madsen 1976 ) that aligns with the anterior margin of metacarpal I. This surface is rugose and pitted like the proximoanterior surface of the first metacarpal, suggesting there were strong ligamentous attachments between these bones. Lateral to the junction of metacarpals I and II, the distal surface ( Fig. 10B View FIG ) becomes concave, rising up to cover the proximal articular surface of the second metacarpal. Concurrent with this, the carpal thins laterally and withdraws from the extensor surface to leave part of the proximal articular surface of metacarpal II exposed anteromedially ( Fig. 11A View FIG ). The posterior (flexor) surface of the carpal is partially composed of a surface of finished bone above the junction of the two metacarpals ( Figs 10D View FIG ; 11D View FIG ). Medial to this, the carpal thins and withdraws from the flexor surface ( Fig. 11A View FIG ) to wrap around the margin of a depression in the proximoposterior surface of the first metacarpal. This is where the troughlike “pulley” articulation of the carpal passes onto the metacarpal. Medial to the depression in the metacarpal, the posterior edge of the carpal approaches the flexor surface of the metacarpal, although it remains a thin edge between convex proximal and distal articular surfaces ( Fig. 11C View FIG ). The proximal surface has a distinct trough that starts from the emarginated posteromedial region ( Fig. 10A View FIG ) and extends anterolaterally to the lateral surface ( Fig. 10F View FIG ). A less prominent troughlike depression isolates the convex anteromedial region of the proximal surface. The distal surface of the carpal ( Fig. 10B View FIG ) has a prominent ridge that follows the junction of the first and second metacarpals. The more lateral articular surface for the second metacarpal consists of shallow anterior and posterior depressions separated by a weak lateromedial ridge (emphasized anteriorly by a shallow trough). The lateral articular surface fits perfectly on a pair of low convex surfaces on the second metacarpal. The medial articulation on the distal surface is more complex. It contacted the first metacarpal in three areas. The most medial one is the largest, and is a flattened saddleshaped articulation. This is separated from the other articulations by a channel that seems to have been utilized by blood vessels or nerves. It is deep and well-defined on the distal surface of the carpal where it enters posteriorly the region of articulation with the first metacarpal from the flexor surface, but becomes less distinct as it divides into two troughs. The more medial one exits the extensor surface via a well-marked canal in the proximal surface of the first metacarpal. The other branch turns to become the shallow trough anterior to the low mediolateral ridge that divides the lateral articulation into anterior and posterior components. Overall, then, the distal articular surface has a total of five contacts with the first and second metacarpals.
The ulnare closely resembles that of Allosaurus (Madsen 1976) . In proximal and distal aspects ( Fig. 11G, H View FIG ), the bone is quadrangular, although the anterolateral side is so short that it looks trian- gular. The anterolateral face has a convex surface and an oval outline ( Fig. 10K View FIG ). The longitudinal axis of the oval is 18 mm long. The posterior side of the carpal ( Fig. 10L View FIG ) is triangular ( 41 mm wide and 23 mm high) and has an almost flat surface. The proximal surface is saddle-shaped (in section it is concave along the anteroposterior axis, and convex perpendicular to that axis) and the distal surface ( Fig. 10H View FIG ) is mostly convex.
The smallest of the carpals is the radiale ( Figs 10 View FIG ; 11 View FIG ), which is 4 cm across and 2.8 cm deep. Most surfaces are convex, and the bone had a sliding anteromedial-posterolateral contact with distal carpal I.
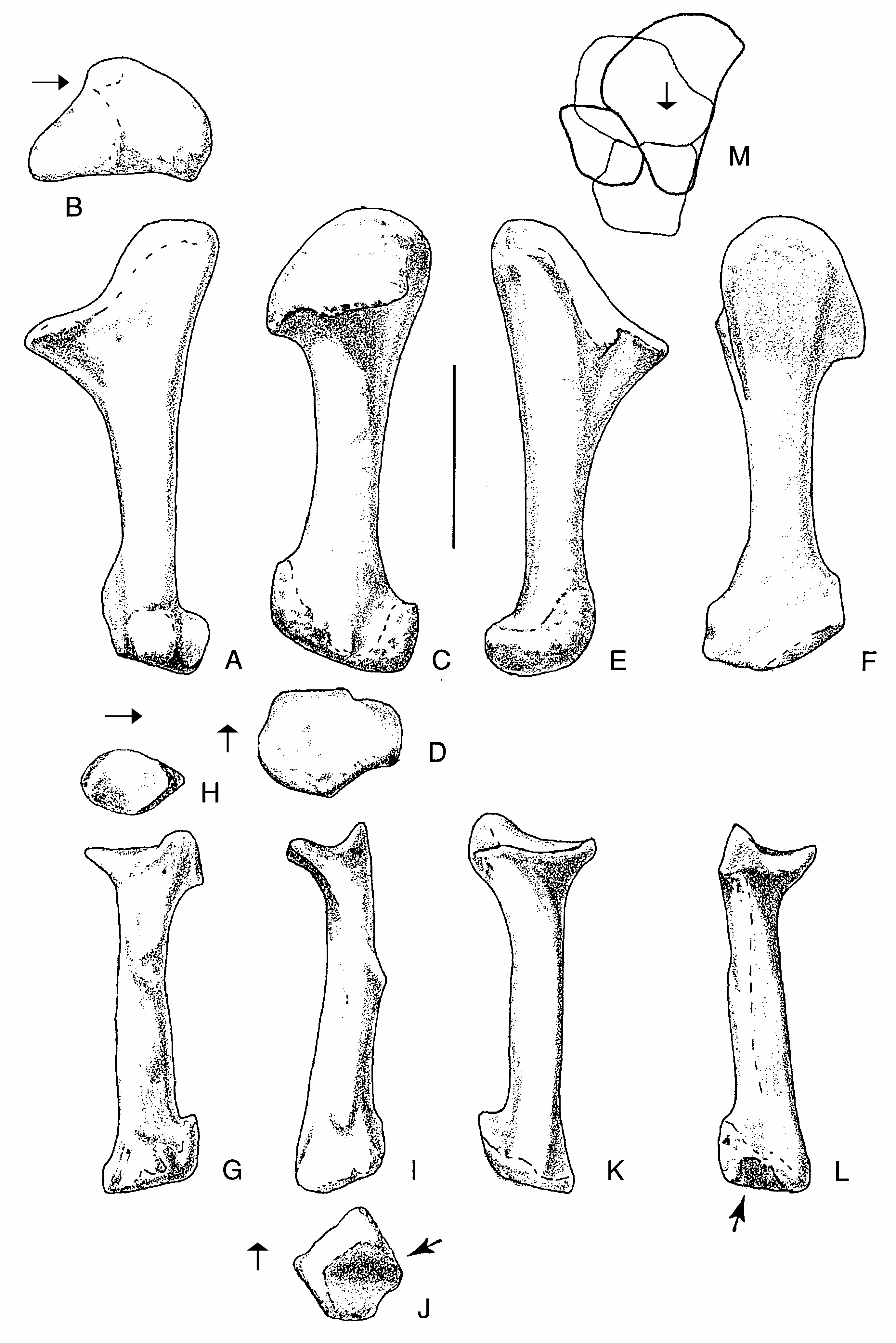
There are three powerfully built metacarpals in Acrocanthosaurus ( Fig. 12 View FIG ). With the exception of the broadly expanded proximal and distal ends, the metacarpals resemble those of Allosaurus (Madsen 1976) , Sinraptor (Currie & Zhao 1993) and other carnosaurs. Szechuanosaurus zigongensis (Gao 1993) also has broadly expanded ends on some of its metacarpals. However, there are many highly significant differences showing this animal is not closely related to Acrocanthosaurus , which suggests that the expanded metacarpal ends probably have functional significance.
The first metacarpal is 62 mm in length. Its proximal and distal heads expand to 50 mm, while the shaft is a flattened oval in section ( 36 mm transversely, 24 mm anteroposteriorly). In comparison, one of the larger first metacarpals from the Cleveland-Lloyd collection of Allosaurus elements is 68 mm long with a proximal expansion of 47, shaft diameters of 45 × 36, and a distal expansion of 53 mm. The proximal end in Acrocanthosaurus is closely applied to the lateral surface of the second metacarpal. The roughly triangular proximal articular surface ( 50 mm lateromedially, 47 mm anteroposteriorly compared with 45 × 50 mm in Allosaurus ) is complex ( Fig. 12A View FIG ). The lateral half slopes lateroventrally, especially where it articulates with the second metacarpal. This articulation consists mainly of a deep depression close to the flexor surface, but it extends anteriorly to the extensor surface as well. The posteromedial face is emarginated for a distinct, smooth-walled depression that is continuous with an emargination on the posteromedial edge of the first distal carpal. This is continuous with the “trough” of the pulley-like surface of the distal carpal. It may represent the end of a sliding articulation of the “pulley-like” wrist joint. As pointed out, a canal for a blood vessel or nerve enters the region between these two bones on the lateral side of this depression. This canal becomes more obvious on the metacarpal as it passes forward and subdivides, forming a deep trough that cuts into the proximal surface of the metacarpal on the anterior (flexor) surface where the bone is emarginated. The Y-shaped system divides the proximal surface into three articular surfaces for the first distal carpal. The deep lateral depression for the second metacarpal and the posteromedial depression for the end of the “trough” of the “pulley” are characteristic of all avetheropodans (= carnosaurs + coelurosaurs).
There are prominent tubercles with rugosities for the attachment of ligaments in the proximolateral ( Fig. 12H View FIG ”) and distomedial regions of the extensor surface ( Fig. 12 View FIG H’). The distal condyles are separated by a deep sulcus ( Fig. 12 View FIG H’). The articular surfaces extend high on the extensor surface, allowing the digit to hyperextend until its shaft is almost perpendicular to the shaft of the metacarpal. In contrast, the first phalanx could not flex much more than 30 degrees. The condyles are asymmetrical so that the first phalanx would have rotated somewhat during extension, when the tip of the claw would have turned more medially. In the degree of rotation and the asymmetry of the condyles, Acrocanthosaurus seems to be different from Allosaurus (Madsen 1976) . There is a deep ligament pit distolaterally ( Fig. 12H View FIG ”), but there is not one medial to the medial condyle ( Fig. 12H View FIG ).
The second metacarpal is 116mm long, with a maximum proximal breath of 74.5 mm, a transverse shaft diameter of 32 mm, and a distal width of 69.5. In comparison,one of the larger second metacarpals from the Cleveland-Lloyd collection of Allosaurus elements is 120 mm long with a proximal expansion of 58, a shaft width of 34 and a distal expansion of 56mm.In proximal outline( Figs11A View FIG ; 12E View FIG ), the lateral, medial and flexor edges are concave, and the extensor margin is mostly convex. The proximal articular surface extends onto the medial surface posteriorly( Fig.12E View FIG )to form a surprising small facet that articulates with the first metacarpal. On the lateral surface, there are two rugose surfaces on the proximal end for a firm, almost suture-like contact with metacarpal III ( Fig. 12E View FIG ”). There is a 3 mm wide canal between the two surfaces that undoubtedly was used by blood vessels and/or nerves ( Fig. 12E View FIG ”). The course of the canal starts in a deep notch in the anteromedial surface of the third metacarpal ( Fig. 12 View FIG A’), continues distally in the canal on metacarpal II, and exits on the flexor surface distal to the contact between metacarpals II and III, just proximal to a prominent ligament tubercle ( Fig. 12 View FIG E’’’). Shallow depressions are all that represent the colateral ligament pits on the medial and lateral condyles. These are highly developed in Allosaurus (Madsen 1976) , Sinraptor (Currie & Zhao 1993) and most other large theropods. The condyles are highly asymmetrical ( Fig. 12 View FIG E’), the lateral one having a much greater anteroposterior surface.The joint allows the first phalanx of the second digit to hyperextend as much as it flexes (about 40 degrees), and turns the digit so that the tip of the claw would have rotated medially during flexion and laterally during hyperextension. As in the first metacarpal, the disparity in distal condyle size and function is much greater than it is in Allosaurus . The third metacarpal is 89 mm long, about 53 mm across proximally, has a shaft diameter of 22 mm, and a distal width of 39 mm. The equivalent measurements in a large Allosaurus are 105 (length), 38 (proximal), 12 (shaft diameter) and 29 mm (distal width). The bone has a pronounced posteromedial expansion ( Fig. 12A View FIG ) where it contacts the second metacarpal in what must have been a virtually immobile joint. There is a suturelike contact with the adjacent metacarpal on the posteromedial expansion, bound anteriorly by a well-defined canal for a blood vessel or nerve ( Fig. 12A View FIG ). The canal divides, part of it passing distoanteriorly into the sutural contact, and the other branch transferring distomedially onto the lateral surface of the second metacarpal ( Fig. 12E View FIG ”). When articulated, the long axis of the third metacarpal is rotated posteriorly so that the centre of the distal end of the bone is about 3 cm behind the centre of metacarpal II.
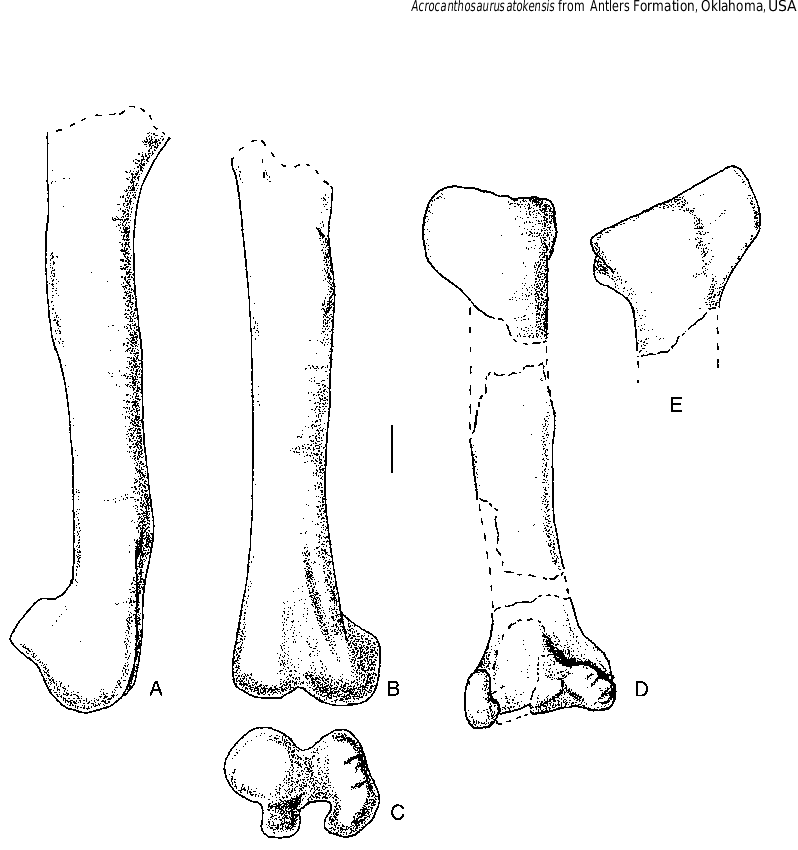
The length of the phalanges of the second digit is almost 90% the length of the humerus. The third digit is 93% as long as the first, which in turn is 73% the length of the second. These proportions are very close to what is found in Allosaurus (USNM 4734, Gilmore1920), which suggests that the stocky appearance of the hand of Acrocanthosaurus is a function of allometric changes. As in Allosaurus , the longest phalanx (excluding the unguals) is I-1, followed by II-2, II-1 and III-3. Phalanges I-1 and II-1 have ginglymoid articulations with their metacarpals, whereas III-1 has a shallowly concave, undivided articulation. Collateral ligament pits are shallow and poorly defined on the manual phalanges ( Fig. 12 View FIG ). Although the femur is incomplete ( Fig. 13 View FIG ), it was at least 110 cm long. The length of the femur can be estimated as 128 cm by comparison with previously described specimens of Acrocanthosaurus (Stovall &Langston 1950; Harris1998a).As in other carnosaurs, the relatively straight shaft of the femur is pierced by a nutrient foramen proximal to the fourth trochanter. Like Allosaurus , Sinraptor and other carnosaurs, the fourth trochanter is represented by a low but prominent ridge next to the depression for the M.caudifemoralis longus .The minimum transverse diameter of the shaft is 15 cm, which is less than the same dimension in adult specimens of Tyrannosaurus (BHI 3033, MOR 555, TMP 81.12.1). This indicates that Acrocanthosaurus was a lighter animal than Tyrannosaurus ,even though the overall length of the body was similar. The distal end of the femur has a ridge along the medial margin of the anterodorsal surface ( Fig. 13B View FIG ) similar to Allosaurus (Madsen1976) , but not as sharply defined as those of Giganotosaurus (MUCPv-CH-1) and Sinraptor (Currie & Zhao 1993) . The ridge bounds the adductor fossa medially. The fossa is continuous with the intercondylar trough of the distal end.There are well-developed distal condyles ( Fig.13C View FIG ), the lateral one associated with a distinct crista tibiofibularis. The tibia is incomplete.However, similarity in size of the distal end of the tibia and the lengths of the
➝
second metatarsals suggests that it would have been close to the same length as that of OMNH 10147 ( 96 cm). There is a well-developed cnemial crest ( Fig. 13D, E View FIG ) and a fibular crest. The tibia flares distally in anterior view ( Fig. 13D View FIG ) where it contacts the astragalus and calcaneum. The anterior surface is flat to accommodate the ascending process of the astragalus. Only the medial portion of the main body of the astragalus was recovered. Overall it appears to have been quite similar to that of Allosaurus with a relatively high, proximolaterally sloping ascending process. The calcaneum is relatively large disk of bone ( 122 mm anteroposteriorly, 66 mm mediolaterally and 99 mm high) with a distinct process that plugged into a depression in the lateral margin of the astragalus as in Allosaurus and Sinraptor .
The first, second, and fifth metatarsals are complete, but only the proximal ends of the third and fourth ones were recovered ( Fig. 14 View FIG ). The second metatarsal ( 41 cm) is 99% of the length of that of OMNH 10147, so the total length of metatarsal III can be estimated as 44 cm. Only the smallest specimens of tyrannosaurids have such short third metatarsals. For example, TMP 91.36.500 is a 5.1 m long skeleton of Gorgosaurus libratus that has a 46 cm long metatarsal III. The length the metatarsus does, however, fall within the range expected for all other large theropods. Generally the foot is closely comparable to those of Allosaurus and Sinraptor , but is more robust. In proximal view, the outlines of the metatarsals ( Fig. 14A View FIG ) are almost identical to those of Allosaurus .
Digit I of the pes, which is complete, is 15 cm long. The first two phalanges of pedal digit III are 16.0 and 11.5 cm, suggesting that the total length of the toe would have been about half a metre. The fourth toe is missing the last two phalanges. The unguals have triangular cross-sections and resemble the pedal unguals of other carnosaurs.

| NCSM |
North Carolina Museum of Natural Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.