
Jubella enucleata Jullien, 1882
|
publication ID |
https://doi.org/ 10.5281/zenodo.208139 |
|
DOI |
https://doi.org/10.5281/zenodo.5694195 |
|
persistent identifier |
https://treatment.plazi.org/id/03E7878D-5B70-FF87-03B0-7348FC76F814 |
|
treatment provided by |
Plazi |
|
scientific name |
Jubella enucleata Jullien, 1882 |
| status |
|
Jubella enucleata Jullien, 1882 View in CoL
( Figs 37–49 View FIGURES 37 – 41 View FIGURES 42 – 45 View FIGURES 46 – 49 ; Table 7 View TABLE 7 )
Jubella enucleata Jullien, 1882: 519 View in CoL , pl. 16, figs 55, 56; Calvet 1907: 377; Prenant & Bobin 1966: 457 fig. 150 (V, VI); d’Hondt 1973: 372, pl. 2, figs 9, 10, 12.
Material examined. Lectotype (designated here): MNHN 2634, Jubella enucleata Jullien, Travailleur, Dr. 42, 44°01'20'' N, 9°25'00'' W, 896 m, 16/8/1881. Paralectotypes (designated here), MNHN 879; MNHN 880; MNHN 2495; MNHN 4025; all from same locality as holotype. Other material examined: MNHN 6914, Thalassa , 476, 44º11.2’ N, 8º40.1’ W, 620 m; MNHN 6925, Thalassa , 503, 44º00.7’ N, 7º06.9’ W, 490 m; MNHN 6973 (part), Thalassa , 512, 44º01.6’ N, 7º01.9’ W, 510–530 m; MNHN 6981, Thalassa , X311, 44º05.8’ N, 4º55.6’ W, 504–515 m; MNHN 6987, Thalassa X325, 44º06.8’ N, 4º45.5’ W, 660–680 m; MNHN 6992, Thalassa , X352, 44º06.5’ N, 4º45’ W, 545–558 m; MNHN 6994, Thalassa , X325, 44º06.8’ N, 4º45.5’ W, 660–800 m; MNHN 7004, Thalassa , X344, 44º06.6’ N, 6º41.0’ W, 530–560 m; MNHN 7011, Thalassa , X351, 44º07.0’ N, 4º44.0’ W, 600–710 m; MNHN 7015, Thalassa , X353, 44º06.8’ N, 06º45.1’ W, 635–655 m; MNHN 7016, Thalassa , X353, 44º06.8’ N, 04º45.1’ W, 635–655 m; MNHN 7026, Thalassa , X362, 44º06.5’– 44º06.8’ N, 4º50.9’– 4º50.8’ W, 600– 585 m; MNHN 7030, Thalassa , X369, 44º05.7’– 44º06.2’ N, 4º42.8’– 4º43.0’ W, 505–580 m; MNHN 7037, Thalassa , X378, 44º07.8’N, 4º44.2’– 04º45.8’ W, 620–680 m; MNHN 7047, Thalassa , W366, 43º36.8’ N, 3º35.5’ W, 250– 380 m; MNHN 7056, Thalassa , W383, 44º04.3’ N, 04º 46.0º W, 524–670 m; MNHN 7059, Thalassa , W391, 44º05.0’ N, 04º33.5’ W, 730–805 m; MNHN 7065, Thalassa , W393, 44º06.09’ N, 4º44.1’ W, 510–970 m; MNHN 7079 (part), Thalassa , W405, 43º56.5’ N, 5º44.0’ W; MNHN 7136, Thalassa , W444, 44º10.1–5’ N, 8º38.4–5’ W; MNHN 7155, Thalassa , W392, 44º06.9’ N, 4º49.3’ W, 600–1130 m; MNHN 7174 (part), Thalassa , W446, 44º12.8’ N, 08º40.0’ W, 620–650 m; MNHN 7204, Thalassa , Y 415, 40º34.3’ N, 9º22.6’ W, 450 m; MNHN 7238, Thalassa , Y 434, 44º12.0’ N, 8º40.8’ W, 620 m; MNHN 7266, Thalassa , U807, 44º11.0’ N, 8º40.2’ W, 450–500 m; MNHN 7293, Thalassa , U833, 43º21.2’ N, 9º30.7’ W, 550–565 m; MNHN 7300, Thalassa , U844, 44º12.1’ N, 8º42.1’ W, 645–760 m; MNHN 7308, Thalassa, 1968 , 44º13.2’ N, 8º 39.5 W, 980–990 m; MNHN 7326, Thalassa , U844, 44º12.1’ N, 8º23.2’ W, 645–760 m; MNHN 7328, Thalassa , U852, 44º12.0’ N, 8º34.0’W, 615–645 m; MNHN 7341, Thalassa , U854, 49º11.0’ N, 8º22.3’ W, 410–640 m; MNHN 7346, Thalassa , U855, 44º11.0’ N, 8º23.2’ W, 520–555 m; MNHN 7373, Thalassa , X313, 44º02.2–8’ N, 4º59.2’ W, 525–580 m; MNHN 7386, Thalassa , X349, 44º06.9’– 44º07.1’ N, 4º43.8’– 4º44.0’ W, 570–625 m; MNHN 7392, Thalassa , X359, 44º07.2’ N, 4º49.4–6’ W, 605–630 m; MNHN 8323, Thalassa X362, 44º06.5’ N, 4º50.9’ W, 585–600 m; MNHN 8375, Thalassa , X315, 43º58’ N, 4º55.7’ W, 950–1000 m; MNHN 8450, Thalassa , X312, 44º04.8’ N, 4º55’ W, 490– 478 m; MNHN 8453, Thalassa , Y 394, 41º18.6’ N, 9º13.8’ W, 410 m; MNHN 8455, Thalassa , X350, 44º04.2’ N, 4º46’ W, 503–507; MNHN 8477, Thalassa , X342, 44º05.7’ N, 4º36.2’ W, 700 m.
Description. Colony erect, jointed, branching, anchored to substratum by basal tuft of rhizoids surrounding ancestrula. Branches unilaminar, formed by three (occasionally four) alternating series of autozooids; their frontal surfaces in three planes nearly perpendicular, fourth plane corresponding to basal walls of two autozooids. Internodes commonly 2–5 mm long, 0.38 mm wide, with 9–24 autozooids, longest in basal part of colony, shorter towards tips.
Autozooids elongate, subrectangular, wider in distal section; distal edge convex, proximally truncate. Gymnocyst reduced to narrow peripheral rim, raised distally as arched peak; rest of the zooidal frontal wall cryptocystal, imperforate. Junction between lateral and frontal surfaces forms two thick raised walls, forming raised rim most pronounced distally, absent proximally. Entire cryptocyst covered by elongate tubercles, conferring wrinkled aspect. Frontal surface deeply immersed in distal part. Opesia subquadrangular, occupying distal third; proximal border concave, distal border convex. Operculum in distal third of opesia. Spines absent. Communication via large uniporous septula. Basal side of branch flat, covered by tubercles; divided longitudinally by thin raised gymnocystal line at zooidal boundary; oblique distal boundary lines angled approximately 45º from branch axis to avicularia. Colony branching dichotomous. In branches with three series of autozooids, each lateral autozooid giving rise to one new branch, being first of lateral series of autozooids. In branches with four series, lateral autozooid follows same pattern; opposing branch formed by autozooid of central opposing series; this zooid highly modified, with wide conical base, originating central series in new branch. Branches initially biserial, unjointed, soon developing cuticularized yellowish joints. Lateral branching sporadic; new lateral branch proximally jointed, arising from the opesia of a lateral autozooid in maternal branch.
Rhizoids growing from orifices located in the middle line of abfrontal face of branch, in proximal area of colony, surrounded by gymnocystal rim of three autozoids. Rhizoids descending abfrontal surface of branch (occasionally also frontal surface), forming thick bundles that are joined in basal tuft surrounding ancestrula, anchoring colony to substratum. Occasionally, these calcified rhizoids giving rise to new branches of autozooids, with long, slender, cylindrical base and pair of distal autozooids.
Each autozooid with triangular avicularium across its distal peak; rostrum triangular, hooked, alternately directed outwards along series; mandible triangular, downcurved. Vibracula and other heterozooids absent.
Modified brooding zooids, of similar length to that of autozooids but wider, scattered along branches. These zooids replacing normal autozooid in longitudinal series but without distal avicularium; membranous area oval, extremely large, occupying three quarters of the zoecial length; operculum semicircular, larger than the autozoidal one. Lateral walls thin and warty, more raised than the autozooidal walls. Distal wall arched, projecting, produced by succeeding autozooid. These modified zooids containing a yellowish substance.
Ancestrula similar to succeeding autozooids, but covered with more marked nodules; its proximal end separated by a chitinous joint from a long tubular extension, joining the substratum.
Discussion. Jubella enucleata was originally described from two live fragments collected at 2018 m from Galicia Bank and three live fragments collected from 896 m, off the northwest of the Iberian Peninsula ( Jullien 1882; Calvet 1907). Five samples are currently conserved at the MNHN, from which we have selected a lectotype and paralectotypes. The species was afterwards reported by d’Hondt (1973, 1974) and by Hayward (1979) from material collected by the Thalassa in north and northwestern Iberian waters at 250–1130 m depth.
SD, Standard deviation; N, number of measurements.
Compared to the original material, Jullien’s (1882) description is quite incomplete, with some easily seen characters overlooked while others are misinterpreted. For instance, rhizoids are obvious in the original material but are not mentioned; neither is a likely ancestrula, seen in one of the colonies.
Jullien mentioned the dichotomous branching, partly amplified by d’Hondt (1973). In the event, J. enucleata has three different ways to produce new branches: firstly, jointed dichotomous branching, varying according to whether the branch has three or four zooidal series ( Figs 46, 47 View FIGURES 46 – 49 ); secondly, lateral branching ( Fig. 48 View FIGURES 46 – 49 ), similar to that in Cellaria salicorniodes Lamouroux, 1816 (see Hayward & Ryland 1998; Hayward & McKinney 2002); thirdly, budding of new branches from calcified rhizoids ( Fig. 49 View FIGURES 46 – 49 ). It seems likely that this diversity of branching methods would allow the colony, which appears very fragile, to quickly recover lost branches and to create denser colonies.
Jullien (1882) interpreted the depressed central area of the autozooid to be a cryptocyst, surrounded distally and laterally by elevated lateral walls. Actually, only the basal part of the autozooidal lateral walls, which is seen forming a thin peripheral rim on the abfrontal face of the branch as well as on the frontal, is gymnocystal. At the distal end of the autozooid this gymnocystal rim raises to form the distal peak over the orifice. The rest of the zooidal surface is a warty cryptocyst decorated by thick granules of calcification. The cryptocyst has frontolateral folds as in many cellariids. The surface of the avicularium is gymnocystal.
Jullien (1882) described the presence of zooids with large elliptical chambers distributed among the branches, considering them to correspond to fertile zooids. Puzzlingly, d’Hondt (1973), after claiming he had not seen ovicells or the elliptical cavities described by Jullien, describes them in detail, and calls them ovicells. As these modified zooids contain a yellowish substance, which could correspond to the remains of one or more embryos, these zooids are likely to be gonozooids. Although these zooids are fairly common in the original material, it seems that Jullien (1882) noticed only those that had lost their delicate frontal membranes. These would therefore be the wide cavities he described.
The phylogenetic position of J. enucleata has been uncertain since its description. It was attributed to its own family by Reverter-Gil & Fernández-Pulpeiro (2001), but without diagnosis or remarks. Traditionally, the genus Jubella has been associated with different taxa placed nowadays in the superfamily Buguloidea ( Jullien 1882; d’Hondt 1973, 1974; Prenant & Bobin 1966; Gordon 2010). The species included in this superfamily exhibit certain characters that are also present in Jubella , such as erect colonies, often jointed, attached by rhizoids, and the presence of avicularia. Individually, none of these characters is diagnostic, being found in a range of cheilostome taxa. On the other hand, the buguloidean family Epistomiidae lacks ovicells, having large brooding zooids (gonozooids) instead. The branching pattern in Jubella is reminiscent of that in some species of the Candidae (Buguloidea) for instance, the relative position of the articulation. Furthermore, in Candidae a type of lateral branching similar to that in Jubella can also appear (in some Notoplites ; Harmer 1926). However, J. enucleata lacks an axial zooid (usually called ‘E’) in basal view, and lateral branching is common to other taxa. Moreover, although it is usually the case that in the superfamily Buguloidea the zooids are usually lightly calcified and the cryptocyst is much reduced or even lacking, well-calcified colonies are present in many Candidae , and some have a well-developed cryptocyst. Given that the gymnocyst is vestigial in Jubella , and that the frontally ridged cryptocyst resembles that in species of superfamily Cellarioidea, we consider that Jubella cannot be included in the Buguloidea.
Species of Cellarioidea have erect, articulated colonies, joined to the substratum by rhizoids, with zooids provided with a depressed, well-developed cryptocyst and lacking articulated oral spines. Furthermore, the form of lateral branching in J. enucleata is similar to that in Cellaria salicorniodes . On the other hand, there are numerous differences. In Cellarioidea the ovicells are endotoichal whereas Jubella has no ovicells but enlarged brooding zooids. Rhizoids in Cellaria or Euginoma are chitinous extensions from by the external cuticle of specialised zooids, while in Jubella they grow from a dorsal pore of the branch, are calcified, and can bud new branches. Also, in Cellarioidea the opesia has been described as typically restricted to the orifice only ( Gordon 1984) or barely larger than the operculum ( Hayward & Ryland 1998). In the apparently related family Membranicellariidae the operculum is clearly smaller, as in Jubella . The Cellarioidea is typically devoid of gymnocyst while in Jubella it is present in the avicularia as well as in a peripheral rim and the distal peak of the autozooids. In conclusion, we consider that the Jubellidae cannot be included in the Cellarioidea.
Finally, another superfamily in which the cryptocyst is well developed is Microporoidea. Taxa in this superfamily exhibit a wide range of characters. A majority of species are encrusting, the amount of gymnocyst varies from moderately developed to absent, and the opesia is often reduced and coincident with the operculum. There are often cryptocystal opesiules. Families such as Calescharidae , Onychocellidae , Lunulariidae , and Otionellidae depart somewhat from the average among Microporoidea, making the superfamily hard to define. Some species of Microporoidea have enlarged ovicells, and gonozooids are found in Chlidoniidae , but none resemble those in Jubella .
Therefore, the precise phylogenetic position of the Jubellidae is elusive on morphological grounds but a case could be made to have it in its own superfamily in the suborder Flustrina.


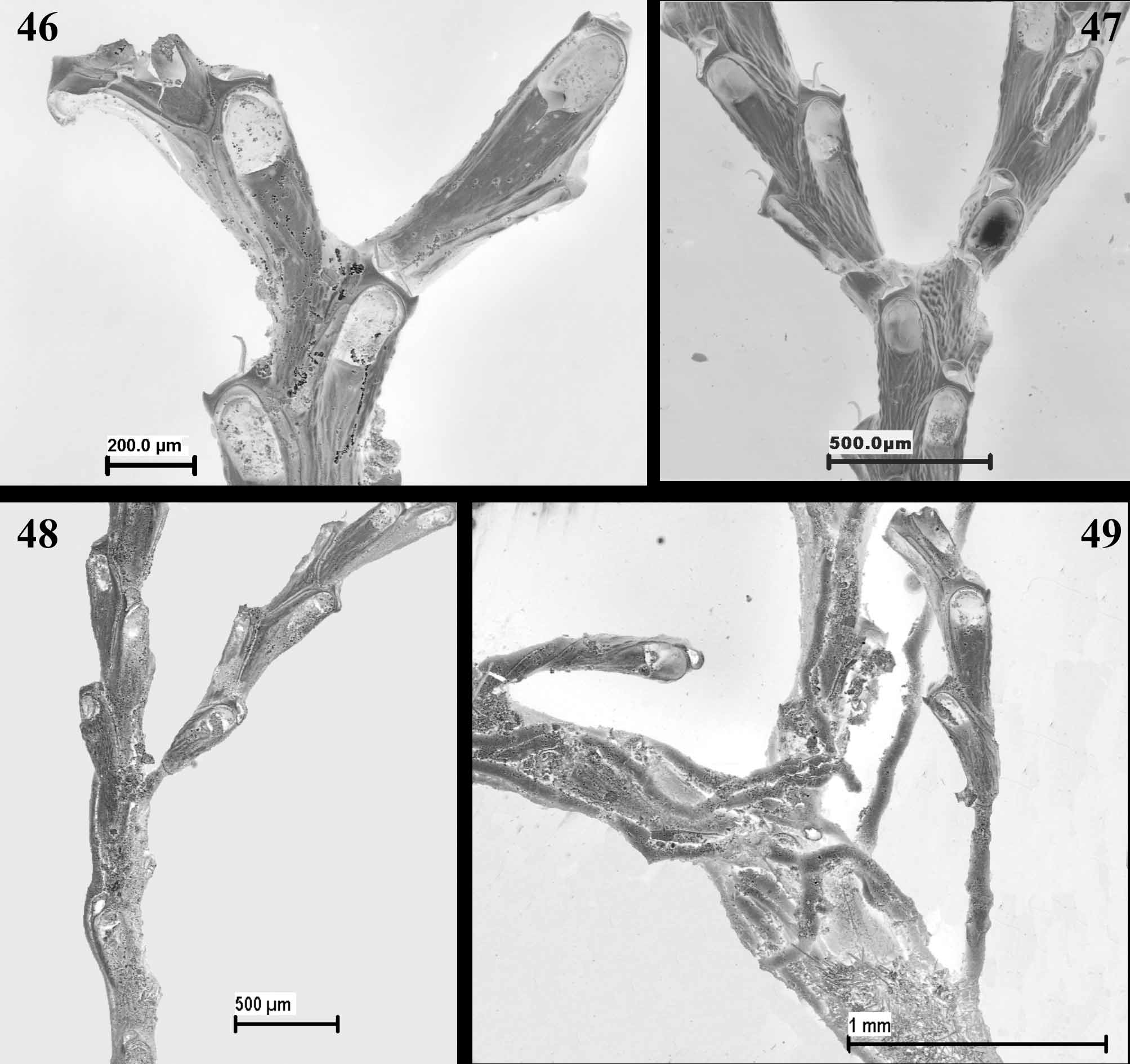
TABLE 7. Measurements (in mm) of Jubella enucleata Jullien, 1882 (MNHN 2634).
| Mean | SD | Minimum | Maximum | N | |
|---|---|---|---|---|---|
| Autozooid length | 0.641 | 0.0571 | 0.557 | 0.777 | 14 |
| Autozooid width | 0.180 | 0.0195 | 0.153 | 0.215 | 14 |
| Opesia length | 0.187 | 0.0170 | 0.158 | 0.216 | 14 |
| Opesia width | 0.131 | 0.0147 | 0.103 | 0.155 | 14 |
| Avicularia lenght | 0.166 | 0.0157 | 0.153 | 0.194 | 6 |
| Avicularia width | 0.097 | 0.0107 | 0.082 | 0.112 | 6 |
| Gonozooid lenght | 0.693 | 0.0095 | 0.686 | 0.704 | 3 |
| Gonozooid width | 0.302 | 0.0104 | 0.290 | 0.310 | 3 |
| Gonozooid opesia length | 0.529 | 0.0061 | 0.522 | 0.534 | 3 |
| Gonozooid opesia width | 0.248 | 0.0086 | 0.240 | 0.257 | 3 |
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Jubella enucleata Jullien, 1882
| Souto, Javier, Reverter-Gil, Oscar & Fernández-Pulpeiro, Eugenio 2011 |
Jubella enucleata
| Prenant 1966: 457 |
| Calvet 1907: 377 |
| Jullien 1882: 519 |