
Pseudosinella pseudopetterseni, Jordana, Rafael & Baquero, Enrique, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.176557 |
|
DOI |
https://doi.org/10.5281/zenodo.5629158 |
|
persistent identifier |
https://treatment.plazi.org/id/03E7879B-FFA7-2B2D-61AE-FE46FE3AE5DC |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudosinella pseudopetterseni |
| status |
sp. nov. |
Pseudosinella pseudopetterseni sp. nov.
Figs 1–6 View FIGURES 1 – 6 , Tables 1 View TABLE 1 and 2
Type material. Mieza, Salamanca ( Spain). Holotype: female, slide 509N Bonet Collection ( MNCN 115coll2003(3)55), C. Bolivar Legit, 7.i.1931. Paratypes: 1 specimen in the same slide as the holotype, in the Museo Nacional de Ciencias Naturales-CSIC, Madrid ( Spain).
Description. Maximum length 1.55 mm (same length in the two specimens). Without pigment (possibly faded along time and we do not know its pattern). Without eyes. Ratio antenna/cephalic diagonal 1.57 and 1.72 respectively. Antennal segments I/II/III/IV ratios 1/3.75/3.5-3.75/6.5-7.5. Sensorial setae s of the sensory organ of antennal segment III short, slightly curved and rod like. Apical region of the antennal segments II and III with a pseudopore in an internal-ventral position, far from the setae line. Apical vesicle absent from antennal segment IV.
Formula of the labial base ( Fig. 3 View FIGURES 1 – 6 ): M1M2REL1L2, all setae ciliated, R as a ciliated mesochaeta, more than 1/ 2 in length of the neighbor ciliate macrochaetae M2.
Formula of the dorsal macrochaetae: R001/00/0201+2. Abdominal tergite II chaetotaxy: paBQ1q2 ( Fig. 1 View FIGURES 1 – 6 ), seta a as broad ciliated microchaeta. R complex with only R0 and R1 ( Fig. 2 View FIGURES 1 – 6 ). Accessory seta s in the anterior trichobothrial complex of abdominal tergite IV present ( Fig. 4 View FIGURES 1 – 6 ). P seta on abdominal tergite IV anterior to the trichobothrium, as a ciliated mesochaeta ( Fig. 4 View FIGURES 1 – 6 ), similar to P. g a m a e Gisin, 1967. Medial seta M1 at the level of the trichobothrium T1. Pseudopore near M 1 in the holotype, and at the same distance to M1 and M 2 in the paratype.
Unguis ( Fig. 5 View FIGURES 1 – 6 ) with dental plate occupying 50% of the basal internal edge; basal teeth of different size, medial tooth well developed, approximately 75% from base of total unguis. Unguiculus appendage acuminate with a fully developed external tooth. Dorsal tibiotarsal tenent hair clavate. Legs without scales. Retinaculum with 4 + 4 teeth and one ciliated seta. One internal and two external setae related to two distal pseudopores of manubrial plate. Mucro with distal tooth longer than the anteapical; the basal spine reaches to the tip of the distal tooth ( Fig. 6 View FIGURES 1 – 6 ).
The characters defined by Christiansen, Bellinger & Gama (1990) for Pseudosinella and the 40 characters used by Christiansen (2007) in his Delta key have been used for identification, but have been added four new characteristics ( Table 2).
Etymology. The species name is referred to the confusion with P. petterseni .
Discussion. According to the dorsal macrochaetotaxy (R001/00/0201+2), and the presence of seta s in the anterior trichobothrial complex of the abdominal tergite IV, this species belongs to the group formed by P. voylesi Christiansen, 1982 and P. leoni Christiansen, 1982 ( Table 1 View TABLE 1 ). Both species are separated by the chaetotaxy of the labium and abdominal tergite II. P. petterseni has been included in this table because the similarity with P. pseudopetterseni sp. nov., that was misidentified as P. petterseni by Bonet (1931).
Remarks. The species with a fully developed external tooth on the unguiculus and without eyes (P. p e t - terseni species group for many authors) are P. petterseni ( Germany) , P. v i o l e n t a ( Folsom, 1924) ( USA), P. rolfsi Mills, 1932 ( USA), P. halofila Bagnall, 1939 ( England) , P. espana Christiansen, 1960 ( USA), P. anderseni Gisin, 1967 ( Switzerland) , P. gamae ( Portugal) , P. biunguiculata Ellis, 1967 (Central America), P. hrabei Rusek, 1979 ( Czech Republic), P. c e r t a Christiansen & Bellinger, 1980 ( USA), P. sera Christiansen & Bellinger, 1980 ( USA), P. ashmoleorum Gama, 1988 (Açores) , P. bachae Luciáñez & Simón, 1994 ( Spain) , P. gajui Luciáñez & Simón, 1994 ( Spain) , P. espanita Christiansen & Bellinger, 1996 ( USA), P. bellingeri Wang, Christiansen & Chen, 2002 (Tibet) , P. c a o i Chen, Wang & Christiansen, 2002 ( China) and P. soriensis Simón & Signoret, 2006 ( Spain) .
Character Range anderseni . P. P ashmoleorum . bachae P 4) (. bellingeri P biunguiculata P. caoi . P. certa P espana . P espanita . P. P gamae . halofila P P hrabei .. petterseni P. P pseudopetterseni P. rolfsi P. sera . soriensis P. P violenta n
.
sp
Ch.1 1–2 1 1 1 1 1 1 1 2 2 1 - 1 1 1 1 1 1 1 Ch.2 1–2 1 1 2 1 1 1 1 1 2 1 - 1 2 1 1 1 1 1 Ch.3 1–7 4 3 4 3 4(1) 4 4 4 4 3 - 4 4 4 3 4 4 3 Ch.4 1–7 4 3 4 3 7 (1) 7 4 3 3 3 - 4 4 4 3 4 4 3 Ch.5 1–5 1 1 1 (1) 5 1 (1) 5 5 5 1 (3) 1 - 1 (1) 1 4 5 1 (1) 1 5 Ch.6 1–5 4 3 4 3 (1) 4 4 4 3 3 3 - 4 4 4 3 4 4 3 Ch.7 1–5 4 3 4 3 4 4 4 3 3 3 - 4 4 4 3 4 4 3 Ch.8 1–5 4 3 4 3 4 4 4 3 3 3 - 4 4 4 3 4 4 3 Ch.9 1–5 4 4 2 4 (1) 4 4 1 1 2 4 - 4 4 2 2(3) 1 4 3 Ch. 10 1–5 4 4 4 4 4 4 4(1) 3 4 4 - 4 4 4 4(3) 4(1) 4 4(3) Ch. 11 1–5 2 2 1 1 1 (2) 1 2 2 2 2 - 2 2 2 2 1 2 2 Ch. 12 1–5 1 4 1 1 1 2 (1) 1 3 2 (3) 4 - 1 1 4 4(3) 1 1 4(3) Ch. 13 1–5 1 1 1 1 1 2 (1) 1 1 2 1 - 1 1 1 1 1 1 1 Ch. 14 1–4 - 4 - 4 4 4 1 2 2 4 - 4 - 4 4 4 - 4 Ch.15 0– n 1 0 3 0 0 0 1 2/3 2/3 0 - 0 2 0 0 0 2 0 Ch. 16 1–3 3 3 - 3 3 3 3 - 2 3 - 3 - 3 3 3 3 3 Ch.17 0–2 0 0 2 0 0 0 0 1 2/3 0 - 0 2 0 0 0 0 0 Ch. 18 1–2 2 1 2 2 2(2) 2(1) 1 2 2 2m (1) - 2 2 2 1 1 2 1 Ch. 19 1–5 2 2(1) 2 2 1 (2) 2 2 2 2 2 - 2 2 2 1–2 2 2 2 Ch. 20 1–2 2 1 2 1 1 (1) 1 - - 1 2 - 2 2 2 1 2 2 2 Ch. 21 1–3 1 1 2 2 1 1 1 1 1 2 1 2 2 2 2 2 2 1–2 Ch. 22 1–4 2 1–2 2 3 2(1) 1 (1) 1 1 2 2 1 (1) 3 2 2 2 2 2 1–2 Ch. 23 1–2 2 1 (1) 2 2 2(1) 2 2 1 2 2 2 2(1) 2 2 2 2(1) 2 2(3) Ch. 24 1–4 4 4 4 4(1) 4 4 4 4 4 4 4 4 4 4 4 4 4 4 Ch. 25 1–3 1 (1) 1(1) 1 1 1 2 1 1(3) 1 3 3 (1) 1(1) 3 1 1 1 1 1
to be continued.
Character, place, description.
Ch.1, dorsal cephalic macrochaeta S, 1. absent/ 2. present;
Ch.2, dorsal cephalic macrochaeta T, 1. absent/ 2. Present;
Ch.3, m1 <ventral labial>, 1. smooth microchaeta/ 2. ciliated microchaeta/ 3. smooth macrochaeta/ 4. ciliated macrochaeta/ 5. smooth macrochaeta with supplementary seta/ 6. ciliated macrochaeta with supplementary seta/ 7. Absent;
Ch.4, m2 <ventral labial>, 1. smooth microchaeta/ 2. ciliated microchaeta/ 3. smooth macrochaeta/ 4. ciliated macrochaeta/ 5. smooth macrochaeta with supplementary seta/ 6. ciliated macrochaeta with supplementary seta / 7. absent;
Ch.5, r <ventral labial>, 1. smooth microchaeta/ 2. ciliated microchaeta/ 3. smooth macrochaeta/ 4. ciliated macrochaeta/ 5. absent;
Ch.6, e <ventral labial>, 1. smooth microchaeta/ 2. ciliated microchaeta/ 3. smooth macrochaeta/ 4. ciliated macrochaeta/ 5. absent;
Ch. 7, L1 <ventral labial, 1. smooth microchaeta/ 2. ciliated microchaeta/ 3. smooth macrochaeta/ 4. ciliated macrochaeta/ 5. absent;
Ch. 8, L2 <ventral labial>, 1. smooth microchaeta/ 2. ciliated microchaeta/ 3. smooth macrochaeta/ 4. ciliated macrochaeta/ 5. absent;
Ch. 9, a <second abdominal seta>, 1. smooth microchaeta/ 2. ciliated microchaeta/ 3. smooth macrochaeta/ 4. ciliated macrochaeta/ 5. absent;
Ch. 10, b <second abdominal seta>, 1. smooth microchaeta/ 2. ciliated microchaeta/ 3. smooth macrochaeta/ 4. ciliated macrochaeta/ 5. absent;
Ch. 11, p <second abdominal seta>, 1. absent/ 2. present;
Ch. 12, q1 <second abdominal seta>, 1. smooth microchaeta/ 2. ciliated microchaeta/ 3. smooth macrochaeta/ 4. ciliated
macrochaeta/ 5. absent;
Ch. 13, q2 <second abdominal seta>, 1. smooth microchaeta/ 2. ciliated microchaeta/ 3. smooth macrochaeta/ 4. ciliated macrochaeta/ 5. absent;
Ch. 14, posterior thoracic segment 2 macrochaeta, 1. acuminate/2. clavate/ 3. truncate/ 4. absent;
Ch.15, posterior thoracic segment 2 macrochaeta, number;
Ch.16, thoracic segment 3 macrochaetae, 1.acuminate/ 2. clavate/ 3. absent,
Ch.17, thoracic segment 3 macrochaetae, number;
Ch.18, anterior lateral (P) fourth abdominal dorsal macrochaetae, 1. 0/ 2. 1/ 2m. 1 mesochaetae;
Ch.19, median (M) 4th abdominal dorsal macrochaetae, 1. 1/ 2. 2/ 3. 3/ 4. 4/ 5. 0;
Ch.20, supplementary seta <4th abdominal segment>, 1. absent/ 2. present;
Ch.21, tenent hair shape, 1. acuminate/2. clavate/ 3. truncate;
Ch.22, number of teeth of inner unguis, 1. 2/ 2. 3/ 3. 4/ 4. 0;
Ch.23, ungual wing tooth, 1. absent/ 2. present;
Ch.24, unguiculus wing tooth, 1. absent/2. minute/3. weak/ 4. fully developed;
Ch. 25, unguiculus shape, 1. acuminate/ 2. clavate/ 3. basally swollen;
Ch.26, eyes per side, number;
Ch.27, inner setae manubrial plaque, 1. 1/ 2. 2/ 3. 3/ 4. 4/ 5. more than 4;
Ch. 28, outer setae manubrial plaque, number;
Ch. 29, habitat, 1. cave/2. surface/ 3. both cave and surface/ 4. under stones at low tide, in the sea;
Ch. 30, region located, 1. Europe and North Africa/ 2. North America / 3. Mexico, Central America, West Indies/ 4. South America / 5. Sub-Saharan Africa/ 6. Asia/ 7. Australia / 8. Oceania/ 9. New Zealand;
Ch. 31, apical antennal bulb, 1. absent/2. present/ 3. Unclear;
Ch.32, apical organ of third ant. segment, 1. peg or rod-like/ 2. expanded (leaf shaped)/ 3. paddle-shaped;
Ch.33, maximum length, mm;
Ch.34, distance distal unpaired ungual tooth from base/total unguis, <%>;
Ch.35, antennal/cephalic diagonal, ratio;
Ch.36, differentiated inner seta on hind tibiotarsus, 1. unclear or absent/ 2. clear, acuminate/ 3. clear, truncate or clavate;
Ch.37, cephalic seta R0, 1. absent/ 2. present;
Ch.38, cephalic seta R1, 1. absent/ 2. present;
Ch.39, cephalic seta R2, 1. absent/ 2. present;
Ch.40, cephalic seta R3, 1. absent/ 2. present;
Ch.41, cephalic seta P, 1. absent/ 2. present;
Ch.42, anterior to pseudopore thoracic segment II seta, 1. absent/ 2. present;
Ch.43, unguiculus external edge, 1. smooth/ 2. serrate;
Ch.44, apical tooth of mucro, 1. similar than second/ 2. longer than second.
The symbol "-" means absence of data.
(1): following original description. (2): Mari Mutt 1986. (3): Christiansen 1998. (4): P. gajui syn. nov. has the same characters of P. bachae except for the Ch. 21, that is "1".
Differences in bold.
The use of the presence/absence of the S and T setae on the head, the formula for the labial, second and third thoracic segments, second and fourth abdominal segments, and the presence of the accessory seta s of the anterior trichobothrial complex of fourth abdominal segment allows to separate the new species of the considered group. These characters have been selected among others because are easily observed in the specimens. The table 2, with the 44 characters frequently used for the description of species of this genus, is sufficient to identify and compare the species of this group. These characters have been obtained studying the original papers of the species descriptions ( Bagnall, 1939; Christiansen, 1960; Christiansen & Bellinger, 1980, 1996, 1998; Gama, 1988; Gama & Busmachiu, 2002; Gisin, 1967; Gisin & Gama, 1972; Luciáñez & Simón, 1994; Rusek, 1979; Wang et al., 2002; Wang et al., 2004) and completed using the program of Christiansen (2007).
P. bachae and P. gajui can be differentiated only by the shape of the tenent hair of the tibiotarsus ( Luciáñez & Simón, 1994). In our opinion this two species are the same because, in addition, P. gajui has been described using only one specimen from the same locality and date of P. bachae . For this reason we consider P. gajui a new synonym of P. bachae .
There are some problems with the characters in Table 2 for P. biunguiculata . In the original description, Ellis drew the p seta of abdominal tergite II, and presented a macrochaetae formula with two M macrochaetae on abdominal tergite IV but without P macrochaeta. Mari-Mutt (1986) compared the types with specimens from Puerto Rico and could not tell the specimens apart in that respect: the p (=a2p) seta is absent, the P macrochaeta is present, and there is only one M macrochaeta. A few specimens from Puerto Rico were different by having R2 on the head. These characters have been updated in Table 2.
The characters are only unknown for P. halophila , but its peculiar habitat (under stones uncovered at low tide, England) allows to suppose that is a valid and different species. In addition only P. ashmoleorum , P. caoi , P. espana and P. c e r t a share with P. halophila by the presence of only two teeth in the inner unguis ( Bagnall, 1939).
The reference for Spain of P. p e t t e r s e n i is erroneous because Bonet (1931) identified the two specimens used for the actual description of P. pseudopetterseni sp. nov. as P. petterseni . Posterior authors used this reference without test the specimens, found by us during the study of the Pseudosinella species in different collections from Spain Museums and particular collections for the preparation of the Entomobryomorpha volume of the “Fauna Iberica Project”.
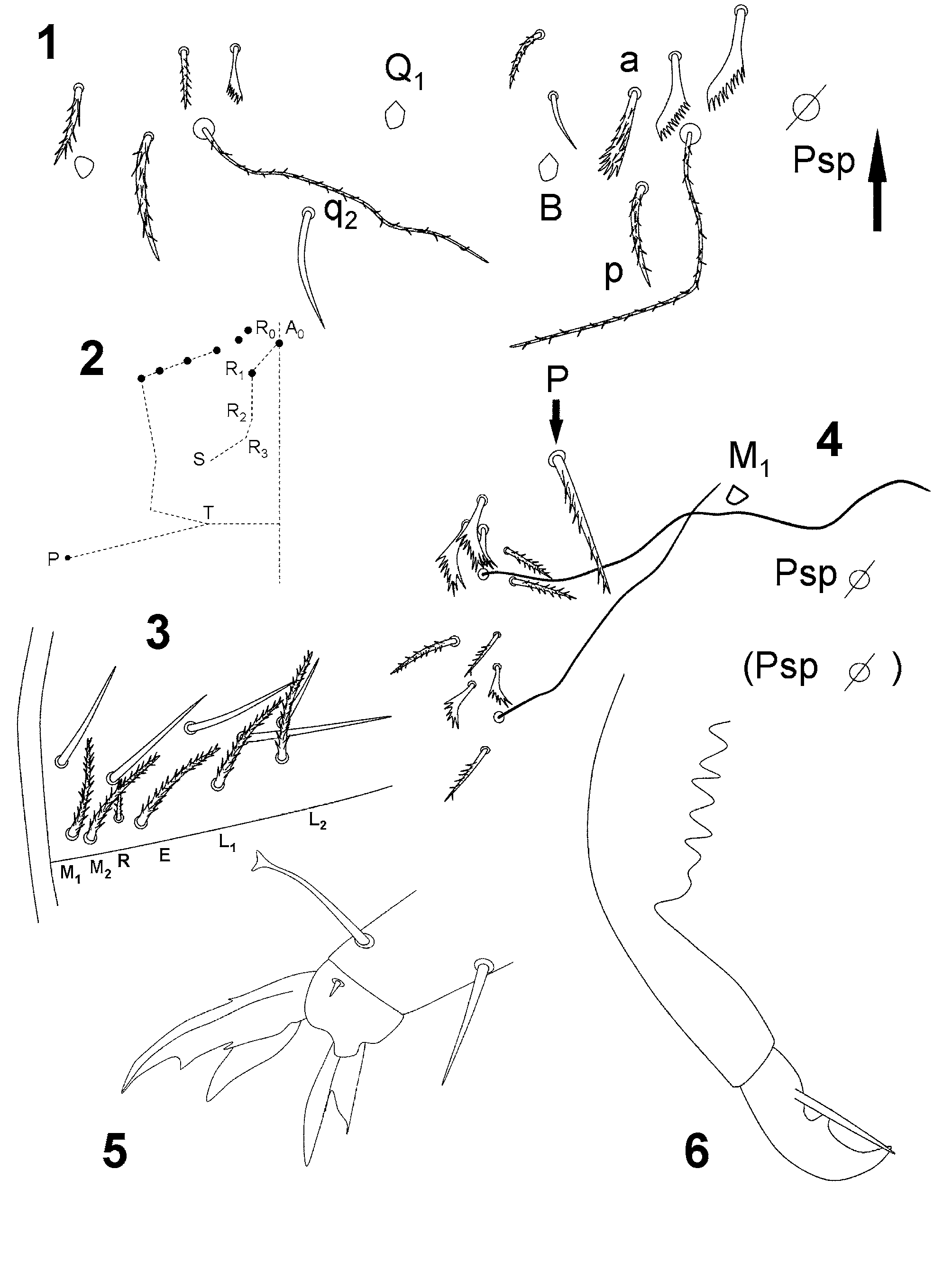
TABLE 1. Comparison of P. pseudopetterseni sp. nov. with P. petterseni and the species with the same dorsal macrochaetotaxy. Abbreviations. Tenent hair: I, pointed; Y, clavate. Distribution: A, North America; H, Holartic; E, Europe; C, caves; S, surface. Labium chaetotaxy: in capitals, seta ciliated; lower-case, seta smooth; -, absence of seta. Differences in bold.
| Species Author | Eyes | Tenent hair | Distribution | Chaetotaxy Abd. segment II | Labium |
|---|---|---|---|---|---|
| P. pseudopetterseni sp. nov. | 0 | Y | E, S | R001/00/0201+2/s p aBQ q | MM R ELL |
| P. voylesi Christiansen, 1982 | 1–2 | I | H, C | R001/00/0201+2/s -ABq q | mm r ell |
| P. l e o n i Christiansen, 1982 | 0 | Y | H, C | R001/00/0201+2/s -ABq q | mm r ell |
| P. petterseni Börner, 1901 | 0 | Y | E, A, S | R011/22/0201+2/s p ABq q | MM r ELL |
| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.