
Essigella (Sorensen, 1994)
|
publication ID |
https://doi.org/ 10.1093/isd/ixy006 |
|
persistent identifier |
https://treatment.plazi.org/id/03E787D1-FFF4-7F5F-FF23-FF177D8EB790 |
|
treatment provided by |
Felipe |
|
scientific name |
Essigella |
| status |
|
Within Essigella View in CoL
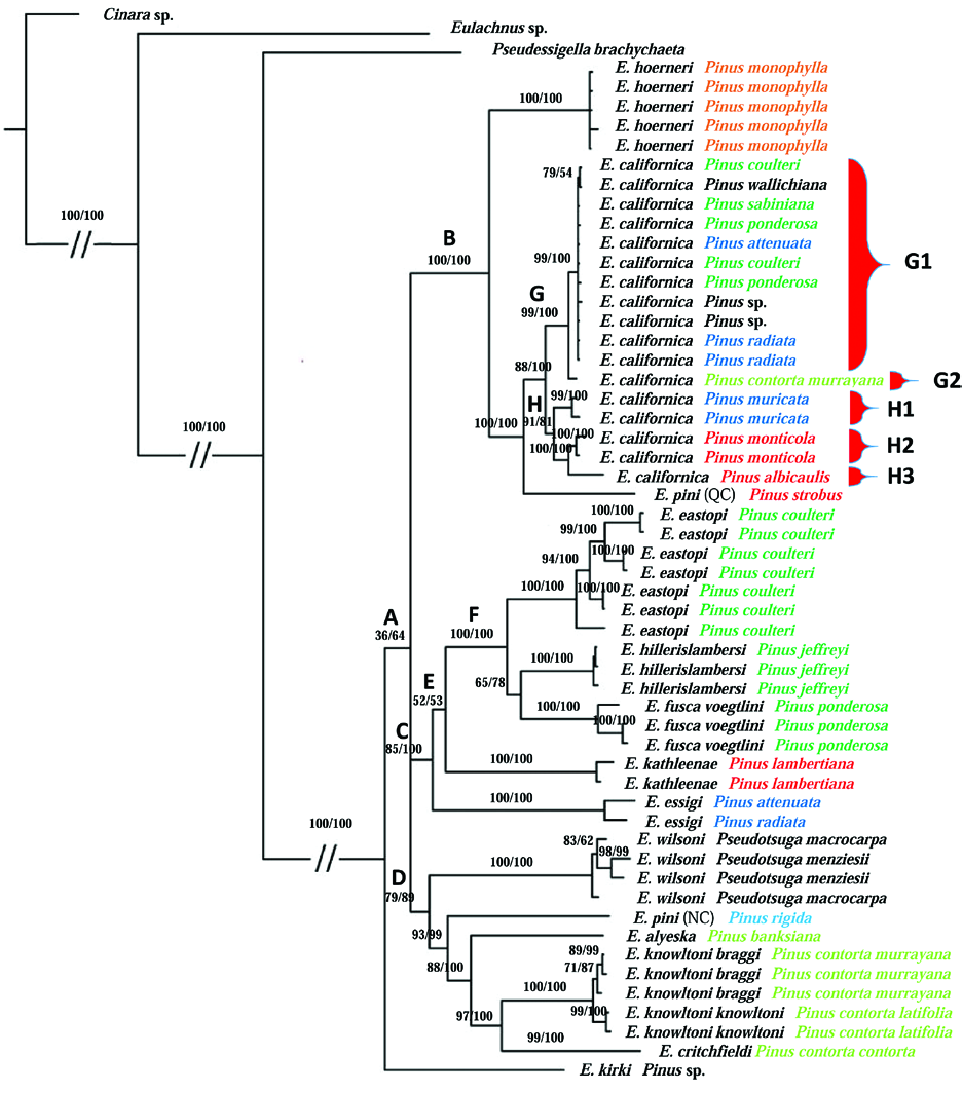
Our phylogenetic results supported neither the division of Essigella into three subgenera Archeoessigella , Essigella , and Lambersella , nor the validity of one of the two species series created by Sorensen (1994) (i.e., Series A). However, our molecular results largely corroborate Sorensen Essigella species concepts. Indeed, we recovered all species complexes and all species that Sorensen delimited with his multivariate analyses, the one significant exception being the two populations of E. pini (Clades B and D, Fig. 2 View Fig ). Several species showed clear internal cladistic structure, however, and merit discussion.
A clear division of E.californica into several clades appeared in our results. The species appeared as two lineages represented by Clades G and H ( Fig. 2 View Fig ). With a COI DNA barcode using a 2% threshold, Clades G1, G2, and H1 may represent one unique species. However, the COI barcode results were not congruent with our ABGD results which consistently separated the Clade H1 from the other E. californica populations ( Table 4). The gaps discovered by the ABGD method can vary from group to group. This flexibility increases the reliability of this method over classical static barcode methods. In aphids, as in animals in general, the genetic distance between two species is considered to be predominantly greater than 2%. However, this useful value is arbitrary and several aphid species have been found showing sequence divergence less than 2%, and even less than 1% ( Rakauskas et al. 2011). In consequence, and because of their placement in our tree ( Fig. 2 View Fig ), we conclude that populations of Clades G and H correspond to at least two distinct species. Moreover, if Clades G and H1 are to be considered two distinct species with a sequence divergence between them of 1.1–1.5%, we would have to consider that the interspecific COI threshold is lower than 2% in Essigella , and possibly near 1.1%. Thus, with a COI sequence divergence P = 2.0% between them (Supplementary Table 1), we consider populations of the Clades H2 and H3, i.e., those found on P. monticola and those on P. albicaulis , to be distinct species as well.
The populations of G1 and the population collected on P. contorta (G2) showed a sequence divergence of P = 0.9–1.2%. These weak and transitional values and the phylogenetic position of G2 do not allow us to decide on the presence of two distinct taxa; additional data are needed to evaluate their species identity. It is possible that these two different groups represent subspecies or that population G2 represents an incipient species.
Morphological comparison with the type series and ecological data suggest that Clade G corresponds to the true E. californica . Sorensen’s analyses (1983), revealed several groups within the E. californica species complex. He divided the complex into two groups, the one with populations developing on pinyon pines (i.e., on pines of the subsection Cembroides like P. monophylla ) which he discriminated as E. hoerneri , and populations developing on nonpinyon pines, which he discriminated as E. californica . Beyond this division, Sorensen (1983) mentioned several other groups within E. californica that were slightly distinct in comparison with the other populations and that could be linked to specific pine species. More specifically, he singled out populations living on Pinus flexilis and P. lambertiana . Despite these observations and following the results of his analyses, Sorensen decided that all these populations living on non-pinyon pines belong to E. californica and that observed variance between them in his analyses could be considered intraspecific variation. No populations of our discriminated MOTUs were collected on P. flexilis or P. lambertiana . Thus, we suspect that E. californica may include two additional cryptic species beyond those revealed in our study.
Species that compose the Essigella knowtoni complex, i.e., E. knowltoni and E. critchfieldi are morphologically similar. Identity of host plant and geographic data are required for species identification ( Sorensen 1994). Despite the strong proximity between these species and the E. knowltoni subspecies, our phylogenetic and molecular delimitation results support their validity. The subspecies appear in different clades and their COI barcode sequence divergence did not exceed 0.8%, whereas the divergence between E. knowltoni and E. critchfieldi was more than 4% (Supplementary Table 1).
Two genetically distinct species were revealed within E. pini in the phylogenetic results, regardless of the genetic locus examined. In our dendrogram, the Canadian population of E. pini clearly belonged to the E. californica complex, whereas the American population appeared related to species of Series B and the E. knowltoni complex ( Fig. 2 View Fig ). Those two species were also revealed as different MOTUs with all species delimitation methods and exhibited a high COI DNA barcode sequence divergence (P = 4.1%). According to these results, one of those species may correspond to E. patchae Hottes, 1957 . Indeed, before Sorensen’s revision, two species were known to occur in the Eastern part of North America. Because Sorensen’s (1994) multivariate analyses did not find any differences between them, he made E. patchae a synonym of E. pini . The issue of multiple species within E. pini will be explored in a future publication.
Species belonging to subgenus Lambersella (also corresponding to the Essigella fusca complex ( Sorensen 1994)) are difficult to distinguish, showing high morphological variability, notably in the length of dorsal metatibial setae ( Sorensen 1994). More particularly, E. eastopi is itself a highly variable species, being easily confused with E. fusca voegtlini . According to Sorensen (1994), several populations of both taxa occurring together in southern California are not distinguishable, suggesting to Sorensen that E. eastopi might be a diminutive form of E. fusca voegtlini . Indeed, during the initial stages of this study, we misidentified all but one population of E. eastopi as E. fusca voegtlini (both species occurring on P. coulteri ). We also misidentified our populations of E. fusca voegtlini as E. fusca fusca Gillette & Palmer, 1924 (both subspecies occurring on P. ponderosa ). We had initially concluded that E. eastopi and E. fusca voegtlini may represent the same species. In the light of our molecular results and following a reappraisal of our slide-mounted specimens, our populations misidentified as E. fusca fusca appear to be closer to E. hillerislambersi than to our E. fusca voegtlini . Moreover, the two subspecies of E. fusca are allopatric ( Sorensen 1994), and according to our collecting data, we collected both subspecies in relative proximity and in a region where E. fusca fusca does not occur ( Sorensen 1994). Either our identifications of Lambersella species were inaccurate, or both subspecies occur in sympatry. Because it is more likely we made wrong identifications, we concluded that we had only collected E. eastopi , E. fusca voegtlini , and E. hillerislambersi . Our mistake underlines the high morphological variability of E. eastopi .
The number of MOTUs from ABGD using COI appeared more important than those obtained using ATP6 and Gnd. However, those obtained with ATP6 appeared more coherent with groups observed with the phylogenetic analyses. This corroborates observations of Lee et al. (2014), who considered that ATP6 would be a better molecular marker than COI in the discrimination of aphid species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.