
Doconesthes dustinchiversi, Reiswig, Henry M., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3920.4.6 |
|
publication LSID |
lsid:zoobank.org:pub:E945E455-0C4E-459C-9F61-443D800643B2 |
|
DOI |
https://doi.org/10.5281/zenodo.5691321 |
|
persistent identifier |
https://treatment.plazi.org/id/03E84366-8D77-4526-C885-EEA6C0AA326A |
|
treatment provided by |
Plazi |
|
scientific name |
Doconesthes dustinchiversi |
| status |
sp. nov. |
Doconesthes dustinchiversi View in CoL n. sp.
( Figs. 1 View FIGURE 1 & 2 View FIGURE 2 , Table 1 View TABLE 1 )
Material examined. Holotype: RBCM 014-00412-01, FV ‘Pacific Viking', set 0 18, 0 6 May 2014, Bowie Seamount, 53 E22.1 ’N, 135 E34.6 ’W, 562–1061 m.
Description. The single specimen ( Fig 2 View FIGURE 2 B–C) is a slightly flattened sac 65.4 x 50.8 x 22.8 mm with a single large osculum 19.6 x 13.7 mm opening to a deep atrium. The body wall is thick, to 8.0 mm on the flattened sides and 9.2 mm on the more abruptly curved sides. Large prostal diactins project singly (2.7 mm apart) or in small groups of 2 or 3 (0.4 mm apart within groups) obliquely upwards, 7.0 (3.1–13.3; 35) mm, from the upper half of the nearly smooth body or low-lumpy surface; prostalia project from margins of low inflations of the body surface, not from the prominences. The sponge is very firm and almost incompressible. The attachment base is missing so it remains uncertain if a basidictyonal mass was present or if the sponge attached to hard substrate with grappling spicules. The dermal surface has an irregular, transparent, thin lattice formed mostly by diactine dermalia supported on a coarser lattice of pentactine and diactine hypodermalia ( Figs 2 View FIGURE 2 D–F). The atrial surface has a regular transparent lattice formed by pinular hexactins ( Figs 2 View FIGURE 2 G–I) oriented long ray out; the lattice covers the entire surface including the apertures of the exhalant canals; hypoatrialia are absent. Body color is light tan with a pink tinge.
Megascleres are diactine prostalia, diactine dermalia (some as monactins to hexactins), hexactine atrialia (some as pentactins), pentactine hypodermalia (some as diactins), and choanosomal diactins. Diactine prostalia are straight or slightly curved, 62% of the spicule exposed and 38% embedded in the body wall; most prostalia have broken outer ends and all are covered by a final shagreen layer of silica ( Fig. 3 View FIGURE 3 A). Dermalia are predominately rough diactins ( Fig. 3 View FIGURE 3 B) with rounded tips; of 300 counted, 90.7% are diactins, 5.7% are triactins, 3% are stauractins and 1% are hexactins. Variants with only 1 ray developed but the one opposite longer than the tubercles of cross rays ( Fig. 3 View FIGURE 3 B bottom) may be classed as either diactins or monactins. Atrialia are predominately pinular hexactins ( Fig 3 View FIGURE 3 C) with the distal ray longest and bearing densely packed proclined thorns; ray tips are sharp. A few (~ 5%) are not pinulate while fewer are pentactins and rarely they are comparatively spiny. Pentactine hypodermalia are large spicules with smooth rays with rough rounded ends; they are the second thickest spicule type after only the prostalia. They range widely in length of proximal ray, categorized as being “very short” (less than half of tangential ray length, Fig 3 View FIGURE 3 D upper) in 38% of hypodermalia, “short” (one-half to the same length as its tangential ray) in 44% of them, and “long” (longer than tangential rays, Fig. 3 View FIGURE 3 D lower) in 18%. Some diactins ( Fig. 3 View FIGURE 3 E) occurring in the hypodermal position with size similar to hypodermal pentactins are considered to also be hypodermalia. Choanosomal diactins ( Fig. 3 View FIGURE 3 F) are long, thin spicules, often broken and difficult to obtain whole; they are lightly curved and generally smooth with rough parabolic tips and insignificant central swellings. They occur in crossing bundles throughout the choanosome.
Microscleres are strobiloplumicomes and oxyhexactins with variants. Strobiloplumicomes ( Fig. 3 View FIGURE 3 G) have a small spherical centrum from which extend the six primary rays, each proximally almost smooth but with a few small lumps and subapically inflated as a sphere which bears the terminal ray bases; the end of the primary ray projects beyond the inflation to end in a bare round-tipped peg 2.5–3.0 µm long. The terminal rays number about 40 per primary ray and originate in 3–5 offset whorls (strobiloid pattern) from the primary ray inflation; they project as overlapping sigmoid curves around the 6 primary axes. They end in a single terminal spine and bear round-tipped spinelets on the inner sides of the strongly curved distal halves; the proximal halves are bare. The oxy-tipped microscleres are mostly (93%) oxyhexactins ( Fig. 3 View FIGURE 3 H) with a few oxypentactins (5%, Fig. 3 View FIGURE 3 I), hemioxyhexasters (2.3%, Fig. 3 View FIGURE 3 J) and paratetractins (0.3%, not figured). The rays bear very fine recurved spines and usually end in straight pointed tips, but hooked tips ( Fig. 3 View FIGURE 3 J upper left) are not uncommon. Individual spicules also may have a mixture of both oxyoid and onychoid tips but these are very rare.
Etymology. The species name, dustinchiversi , commemorates the life of Mr. Dustin Chivers, an outstanding marine biologist who spent most of his career with the California Academy of Sciences in San Francisco, California and died in 1995. He was both my friend and colleague. He introduced me to the beauty and diversity of marine organisms during our high school days and, most important for me, saved my life in a 1962 diving accident. I owe my success and accomplishments to Dusty's presence and strength of character that day; everyday I thank him for my life.
Remarks and discussion. The new species differs from the three descriptions of the North Atlantic Doconesthes sessilis in having diactins as long prostalia lateralia. In other respects, the three descriptions of D. sessilis (here Topsent 1928 as 1, Tabachnick and Collins 2008 as 2, Tabachnick & Menshenina 2013 as 3) differ quite significantly from each other. Prostalia were entirely absent in 1 and 3 but projecting hypodermal pentactins were present in 2. Hypodermal pentactins with short proximal rays were reported in 1 but not 2 or 3. Hypoatrialia were absent in 1 and 2 but differentiated from atrialia in 3. Dermal diactin lengths do not overlap at all as given in the three reports: longest in 1 (320–390 µm), shorter in 2 (156–280 µm) and shortest in 3 (23–68 µm). For oxytipped microscleres, none of the three reports are sufficiently quantitative to allow comparisons between them or with the new Pacific specimen, e.g. in 1 and 2 they are reported as “most are hemioxyhexasters”, in 3 as “oxyhexactins, hemioxyhexasters and oxyhexasters” but without relative abundances. Some of these and other differences between specimen descriptions could be attributed to condition and size (maturity) of the specimens, but it is extremely unlikely that dermalia would differ significantly in size. Authors of the latest report (3) have been conservative in interpreting the two later specimens as belonging to D. sessilis but with limited specimens of very different size, their conclusions were reasonable. However, with addition of the nearly complete specimen of a new Northeast Pacific species, a re-evaluation of the differences between the North Atlantic specimens appears warranted.
I thank Melissa Frey, Curator of Invertebrates at RBCM, for bringing the specimen to my attention and to Archipelago Marine Research, Victoria, BC, for providing commercial fishery bycatch to RBCM. SEM work was carried out with assistance of Brent Gowan at the Department of Biology Advanced Imaging Laboratory and Elaine Humphrey, Department of Mechanical Engineering Advanced Microscopy Facility, both at University of Victoria. Funding for the SEM work was provided by the Royal British Columbia Museum.
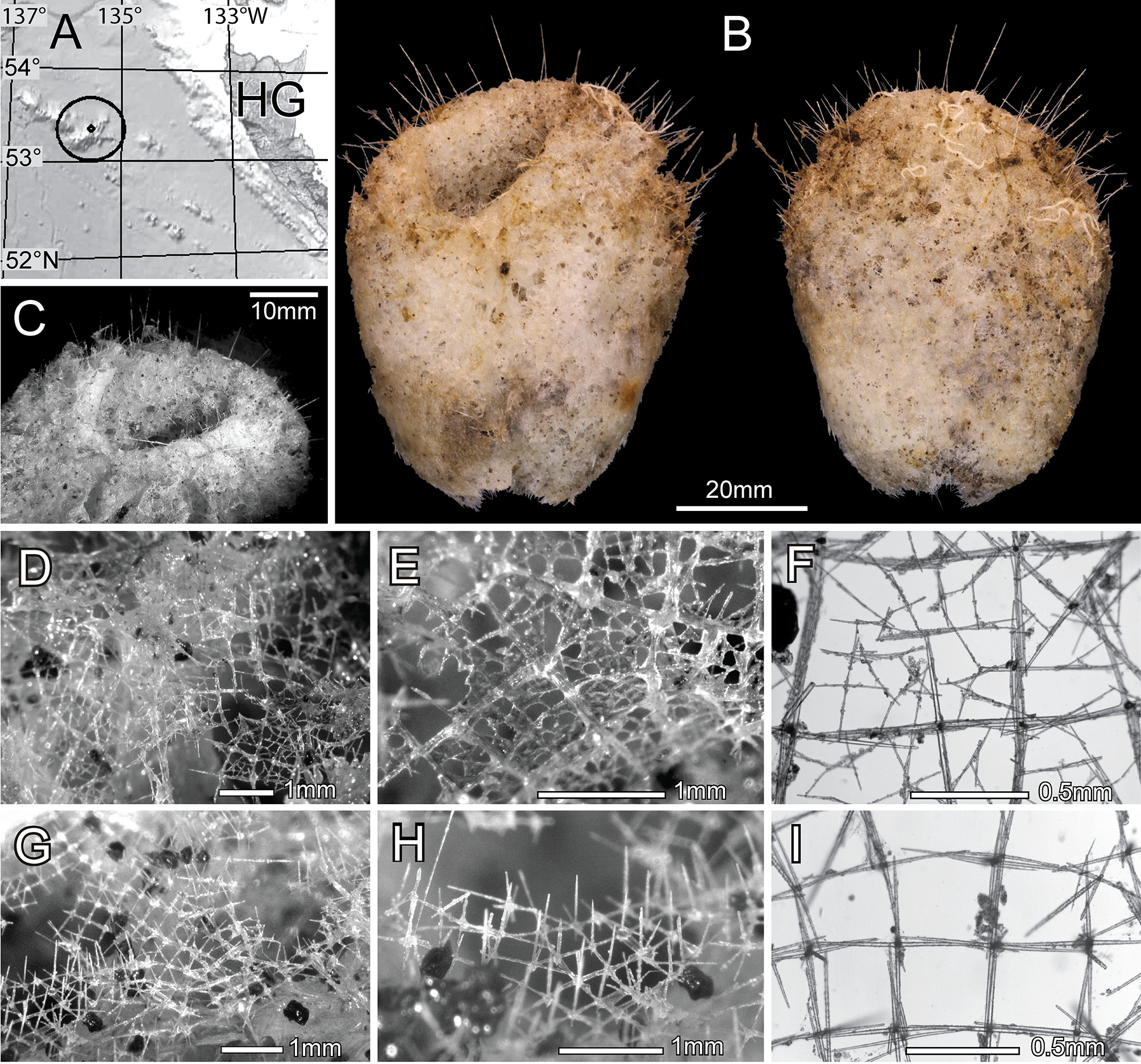

TABLE 1. Spicule dimensions of Doconesthes dustinchiversi, RBCM 014 - 00412 - 01, from British Columbia, Canada (dimensions in µm unless otherwise indicated).
| parameter | mean | st. dev. | range | number |
|---|---|---|---|---|
| Prostal diactin length (mm) | 14.67 | 5.47 | 7.59–21.31 | 6 |
| width | 112 | 21 | 85–146 | 6 |
| Dermal diactin length | 273 | 33 | 224–353 | 50 |
| width | 8.3 | 2.0 | 5.0–13.3 | 50 |
| triactin axial length | 233 | 49 | 133–360 | 50 |
| stauractin axial length | 208 | 52 | 126–353 | 50 |
| pentactin axial length | 245 | 30 | 225–266 | 2 |
| hexactin axial length | 177 | 23 | 142–208 | 6 |
| Atrial pinular hexactin | ||||
| distal ray length | 472 | 123 | 89–733 | 50 |
| distal ray width | 15.5 | 3.7 | 7.8–28.3 | 50 |
| tangential ray length | 288 | 69 | 159–431 | 50 |
| tangential ray width | 14.5 | 3.4 | 7.5–25.4 | 50 |
| proximal ray length | 144 | 44 | 82–284 | 50 |
| proximal ray width | 14.1 | 3.1 | 8.0–23.8 | 50 |
| Hypodermal pentactin | ||||
| tangential ray length | 599 | 91 | 382–782 | 50 |
| width | 27.4 | 2.9 | 21.0–33.8 | 50 |
| proximal ray length | 395 | 196 | 144–841 | 50 |
| width | 25.6 | 3.1 | 20.0–31.7 | 50 |
| Hypodermal diactin length | 1142 | 370 | 501–1601 | 18 |
| width | 16.1 | 3.4 | 10.0–21.2 | 18 |
| Choanosomal diactin length | 4066 | 1331 | 1473–7155 | 50 |
| width | 19.8 | 3.9 | 9.0–28.6 | 50 |
| Strobiloplumicome diameter | 43.5 | 4.1 | 33.1–52.6 | 63 |
| primary ray length | 9.0 | 1.2 | 5.9–14.1 | 63 |
| secondary ray length | 15.2 | 1.7 | 11.7–18.8 | 63 |
| Oxyhexactin diameter | 141 | 19 | 70–183 | 50 |
| Hemioxyhexaster diameter | 152 | 11.1 | 140–180 | 15 |
| primary ray length | 7.4 | 1.8 | 5.2–10.3 | 15 |
| secondary ray length | 69.4 | 4.6 | 62.6–80.1 | 15 |
| Acknowledgements |
| RBCM |
Royal British Columbia Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.