
Aaptos Gray, 1867
|
publication ID |
https://doi.org/10.5281/zenodo.183878 |
|
DOI |
https://doi.org/10.5281/zenodo.6235805 |
|
persistent identifier |
https://treatment.plazi.org/id/03E887DC-5E76-FFA7-5EC9-16C8FBF1A919 |
|
treatment provided by |
Plazi |
|
scientific name |
Aaptos Gray, 1867 |
| status |
|
Genus Aaptos Gray, 1867
Diagnosis (sensu van Soest 2002):
Sponges are lobate, spherical, subspherical or globular often consisting of confluent globular or lobate units. Surface smooth or tuberculatepapillate, usually rough to the touch. Some species show a distinct colour change when taken out of the water. In cross section, the outer region is often fibrous and may be considered as a cortex which grades into the choanosome. Skeleton strictly radial, with tracts and single spicules issuing from the centre of the lobe or globular body. At the surface the tracts fan out and form a dense palisade consisting of smaller spicules intermingled between the ends of the larger spicules. Spicules are strongyloxeas, in three overlapping size categories; the intermediate and smaller spicules are occasionally oxeas, styles or tylostyles.
Type species: Ancorina adriatica Gray, 1867 (by original designation) = Aaptos aaptos (Schmidt, 1864) .
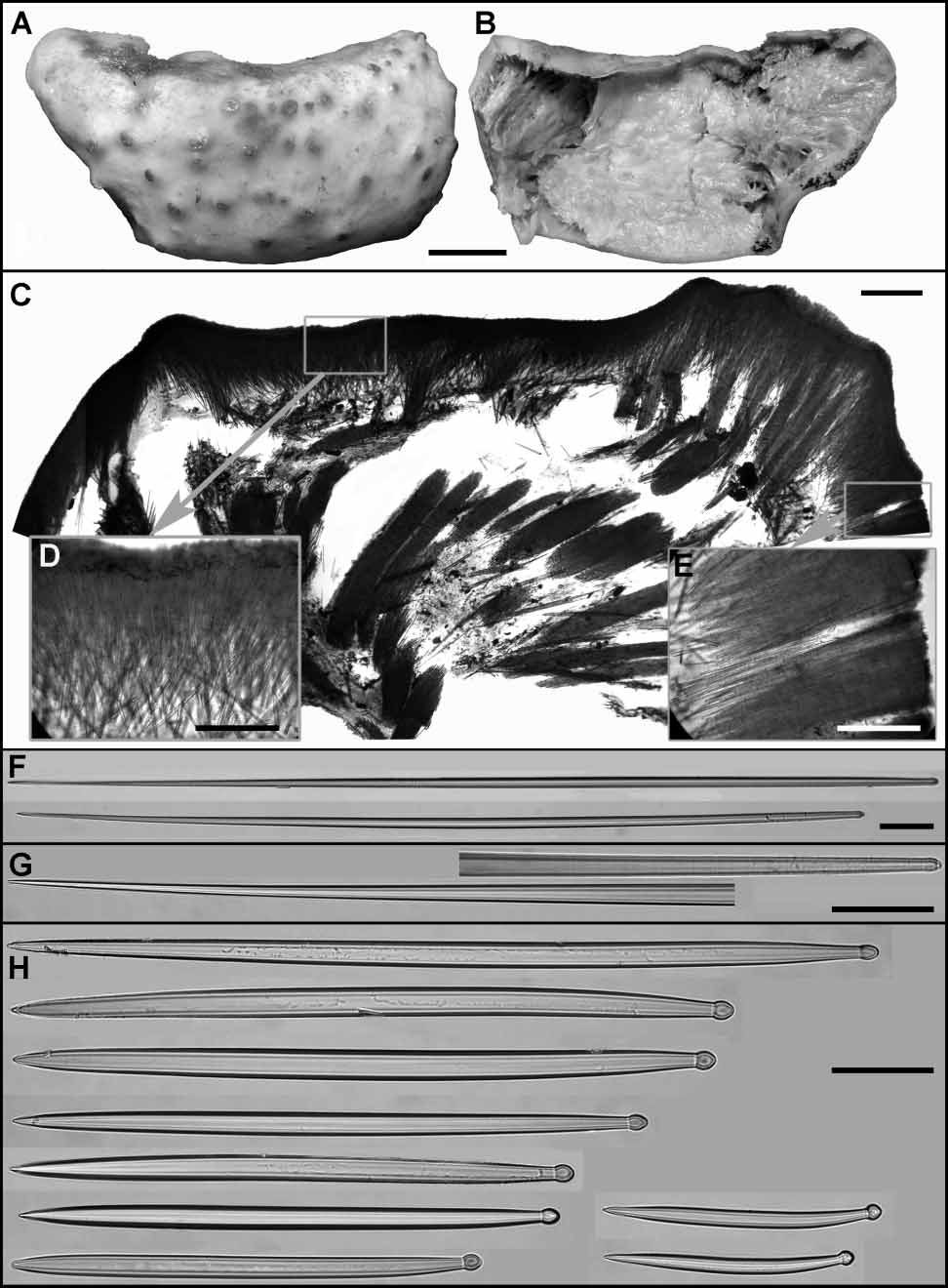
Aaptos robustus sp. nov. ( Fig. 14 View FIGURE 14 )
Material examined
Holotype: SMF 10579: PS61/1419.
Description
External morphology. The holotype is a cushionshaped sponge originally attached to a stone but detached from it on the deck (figs. 14A, B). The dimensions are approximately 55x26 x 7 mm. Surface is smooth, slightly rough to the touch, with numerous gentle elevations at the tops of which small oscula are opened. Surface colour was light brown in life and has become pale yellow in alcohol, the elevations have remained darker. The cortex is pale yellow to white, very hard and firm in consistency, and cannot be detached from the choanosome. The latter is also hard but crumbly, its colour pale yellow. Numerous aquiferous cavities are located under the cortex and in the internal choanosome.
Skeleton. The choanosomal skeleton is constituted by the radial tracts of principal strongyloxeas running from sponge base to the surface and measuring 150–500 µm in thickness (fig. 14C). Cortical skeleton made of a layer of small and intermediary tylostyles which are mainly arranged perpendicularly to the surface but also sometimes obliquely to it (fig. 14D). The cortex measuring 1–1.2 mm in thickness is perforated by rare, small ostia (fig. 14E).
Spicules. Three size categories of spicules are well distinguished. Thirty spicules of each category were measured.
Principal strongyloxeas are mainly straight, rarely slightly curved (figs. 14F, G) and measure: length 13781647 1780 µm, proximal diameter 1011.312 µm, central maximal diameter 1416.321 µm. Small and intermediary tylostyles are always stout and fusiform, with welldeveloped, terminally located, spherical or oval tyles (fig. 14H). Intermediary tylostyles are mainly straight and measure: length 470618830 µm, diameter of the tyle 1315.518 µm, diameter of the shaft underneath the tyle 910.812 µm, maximal diameter of the shaft 2023.328 µm. Small tylostyles are often slightly curved. Their dimensions are: length 250259 300 µm, diameter of the tyle 1414.616 µm, diameter of the shaft underneath the tyle 99.911 µm, maximal diameter of the shaft 1414.916 µm.
Type locality
SE Atlantic: to the East of the South Sandwich Islands, ca. 2300 m.
Etymology
The name refers to the hard consistency of the cortex (Latin robustus =hard, robust).
Remarks
No species of Aaptos has previously been registered in the Antarctic and deeper than 1000 m either, although Sarà et al. (1992) included A. unispiculus (Carter, 1880) in the Antarctic faunistic complex. In the meantime, the taxonomy and geography of this species are rather confused. Hymeraphia unispiculum described by Carter (1880) from the Gulf of Manaar, tropical Indian Ocean, was a thinly encrusting, hispid sponge with long rhabdostyles echinating the surface. De Laubenfels (1936) proposed transferring H. unispiculum to Aaptos without examination of any material and in spite of evident disparities. Later de Laubenfels (1954) recorded A. unispiculus from the Marshall Islands, tropical North Pacific, probably from shallow waters. His specimen really shared some features with Aaptos spp., at least it possessed a surface palisade of styles, but any resemblance to Carter’s description could hardly be observed. Following the actions of de Laubenfels, Desqueyroux (1972) and Desqueyroux & Moyano (1987) reported A. unispiculus as deep as 200– 400 m, from the SE Pacific, off Juan Fernandez Islands and in the Magellan area of the Chilean Coast, respectively. Subsequently the latter record was taken into account by Sarà et al. (1992). The SE Pacific sponges were thickly encrusting, hard and hispid. Their confused choanosomal skeleton was made of styles and microxeas, and the distinct dense cortex contained the styles oriented either tangentially or perpendicularly to the surface. Thus, the placement of H. unispiculum in Aaptos is hardly justified. Currently, the species in question is accepted as Eurypon unispiculum , both Eurypon and Hymeraphia belong to the family Raspailiidae Hentschel, 1923 . Subsequently, the sponges recorded as A. unispiculus from the tropical and subantarctic Pacific, belong in fact to some other species, which, probably but not for certain, may belong to Aaptos . There is at least no doubt that these species share very few features with A. robustus sp. nov. described herein.
Meanwhile, seven valid species of Aaptos are currently known from the temperate shallow waters of the Southern Hemisphere – four New Zealand species, A. confertus , A. globosum , A. rosacea and A. tentum , all described by KellyBorges and Bergquist (1994); one South Australian species, A. horrida (Carter, 1886) ; and two South African species, A. nuda (Kirkpatrick, 1903) and A. alphiensis Samaai & Gibbons, 2005 . The latter six species share a globular or (sub)spherical body and a strictly radial choanosomal skeleton composed of the main spicule tracts which issue from the body center. Conversely, A. robustus sp. nov. described herein is characterized by a cushionshaped body and a choanosomal skeleton constituted by the tracts starting from sponge base. Our new species probably has the growth pattern similar to that of A. confertus . The body of the latter is said to result from the confluence of several strictly globular or spherical units, and subsequently its skeleton is considered as a compilation of several strictly radial subskeletons ( KellyBorges & Bergquist 1994). We suppose that A. robustus sp. nov. demonstrates even more complete confluence of the units than A. confertus , so that these units can hardly be recognized.
Concerning the skeletal arrangement and spicule composition, our new species shares the main choanosomal tracts made of principal strongyloxeas, and the cortex constituted by a single palisade of smaller spicules with A. confertus , A. globosum , A. rosacea , A. tentum and A. horrida , whereas A. nuda lacks any cortical or even ectosomal skeleton, and A. alphiensis is distinguished by the principal spicules being subtylostyles and by the underpalisade layer of densely packed, tangentially oriented spicules. The cortical palisade of Aaptos spp. is often composed of two categories of spicules ( KellyBorges & Bergquist 1994). In A. robustus sp. nov. and A. confertus these categories are distinguished only by size and presented by fusiform, straight tylostyles in the former species and by slender, curved and/or bent styles in the latter species. In A. globosum , A. rosacea and A. tentum these are small, fusiform tylostyles and larger, slender subtylostyles or styles, and in A. alphiensis they include small tylostyles and larger styles.
Thus, the principal spicules and two size classes of the cortical spicules constitute altogether three spicule categories of A. robustus sp. nov., and no other spicules were observed in this species. Conversely, most other Aaptos spp. possess a fourth category of spicules being intermediary in size between the longer spicules of the palisade and the principal megascleres, and embedded in the fanning main tracts protruding the cortex. In A. confertus , A. rosacea and A. alphiensis , such an intermediary category is constituted by styles or subtylostyles, and these are also abundantly scattered in the choanosome between the main tracts. In A. tentum and A. globosum the intermediary spicules do not differ from the principal strongyloxeas in shape and are rarely found (in the former species) or absent (in the latter species) in the choanosome.
| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |