
Radiella antarctica, Plotkin, Alexander S. & Janussen, Dorte, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.183878 |
|
DOI |
https://doi.org/10.5281/zenodo.6235795 |
|
persistent identifier |
https://treatment.plazi.org/id/03E887DC-5E7A-FFAD-5EC9-1635FE0EAF99 |
|
treatment provided by |
Plazi |
|
scientific name |
Radiella antarctica |
| status |
sp. nov. |
Radiella antarctica sp. nov. ( Figs. 7–9 View FIGURE 7 View FIGURE 8 View FIGURE 9 , Table 4)
Material examined
Holotype: SMF 10558: PS67/1102.
Paratypes: SMF 10559 (2 specimens): PS61/1323; SMF 10560 (10 specimens), 10561 (7 specimens) and 10562 (1 specimen): PS61/1353; SMF 10563 (66 specimens) and 10564 (1 specimen): PS67/10211; SMF 10566 (62 specimens), 10567 (42 specimens) and 10568 (2 specimens): PS67/1102; 10565 (1 specimen), 10569 (1 specimen) and 10570 (1 specimen): PS67/1108.
Description
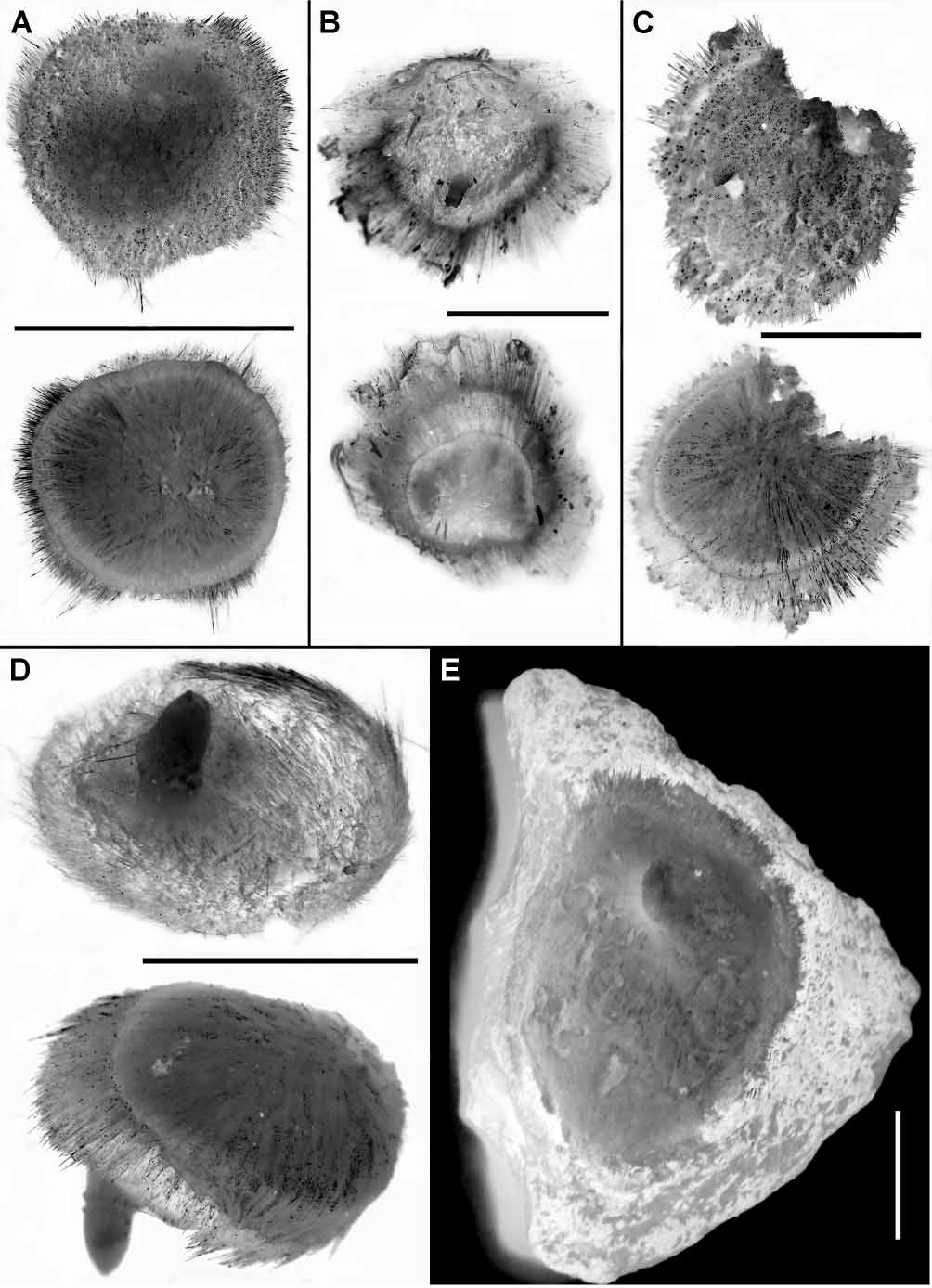
External morphology. The holotype is a lenticular, slightly irregular sponge measuring approximately 9 mm in diameter and 2 mm in thickness, without any substrate (fig. 7A). The central area of the upper surface is gently rough and has a pale colour in alcohol, with a single very low exhalant papilla of 1 mm in diameter. The periphery of the upper surface is hispid and covered with greyish sediment particles. The basal surface is nearly smooth and pale coloured. A spicule fringe of 0.7 mm in width is in the margin between the upper and basal surfaces. The cortex is hardly detachable, whitish in colour and moderately firm in consistency. The choanosome is whitish, firm but crumbly.
All paratypes share the presence of solitary exhalant papillae and the marginal fringes with the holotype. The colour of their upper surfaces is mainly determined by the hispidation pattern and the subsequent density of the soiling sediment particles. The basal surface, if present, is usually pale, and the papillae are of the same colour. The cortex is hardly detachable, whitish to grey in colour and moderately firm or slightly resilient in consistency. The choanosome is whitish to pale beige. Other external features of the paratypes vary much more. The paratype SMF 10564 is a lenticular, slightly irregular sponge without any substrate (fig. 7B). The paratype SMF 10565 is a discoid sponge attached to a very small sediment particle by the central area of its basal surface (fig. 7C). Both specimens SMF 10564 and 10565 possess uniformly hispid upper surface covered with sediments and a smooth basal surface. Another paratype SMF 10562 (fig. 7D) has a discoid upper surface with a smooth pale central area and a hispid periphery, and a conical, smooth basal surface without substrate.
Most other studied sponges are attached by the basal surface to various hard substrata including pebbles, rock pieces and volcanic concretions (fig. 7E). The shape varies from discoid to hemispherical and cushionlike. The diameter is up to 30 mm while the thickness may reach 8 mm. The upper surface is mainly smooth, velvet or gently rough though its peripheral area close to the marginal fringe may be hispid. The length of the papillae varies from 0.5 to 4 mm; the diameter is 1–2 mm.
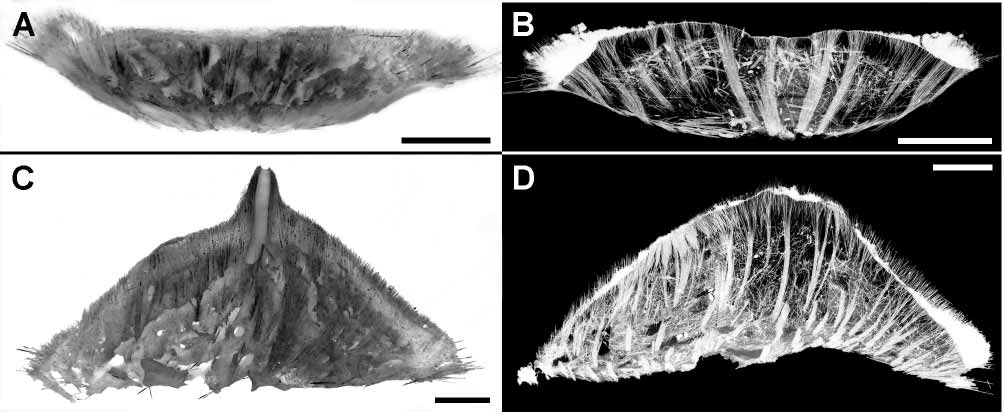
Skeleton. The main choanosomal skeleton is constituted by the tracts of principal tylostyles radially running from the central basal area (fig. 8). The thickness of the radial tracts is 110–330 µm in the holotype and 100–280 µm in other specimens. In sponges free of substrate, or with a small substrate, the radial tracts form a smooth envelope of the basal surface (figs. 8A, B). In specimens attached to larger substrata, such an enveloping is less visible (figs. 8C, D). The additional choanosomal skeleton is composed of the variously positioned principal and small tylostyles lying singly or gathered in the thin bundles. In the upper surface the main choanosomal tracts enter the cortex, diverge into bouquets and make up the surface hispidation developed mainly in the periphery. The cortical bouquets are reinforced by the palisade of small tylostyles. Ostia are located between the bouquets. The thickness of the cortex is 250–680 µm in the holotype and 210–800 µm in other specimens. The thickness of the surface hispidation is 120–630 µm in the holotype and 260–410 µm in other specimens. The marginal fringe is made of the additional extralong tylostyles which may also reinforce the peripheral hispidation. Papilla skeleton is constituted by the ascending main tracts surrounding the exhalant canal of 540 µm in diameter The skeleton of each papilla possesses ascending main tracts surrounding the exhalant canal, the latter measuring 540 µm in diameter (fig. 8C).
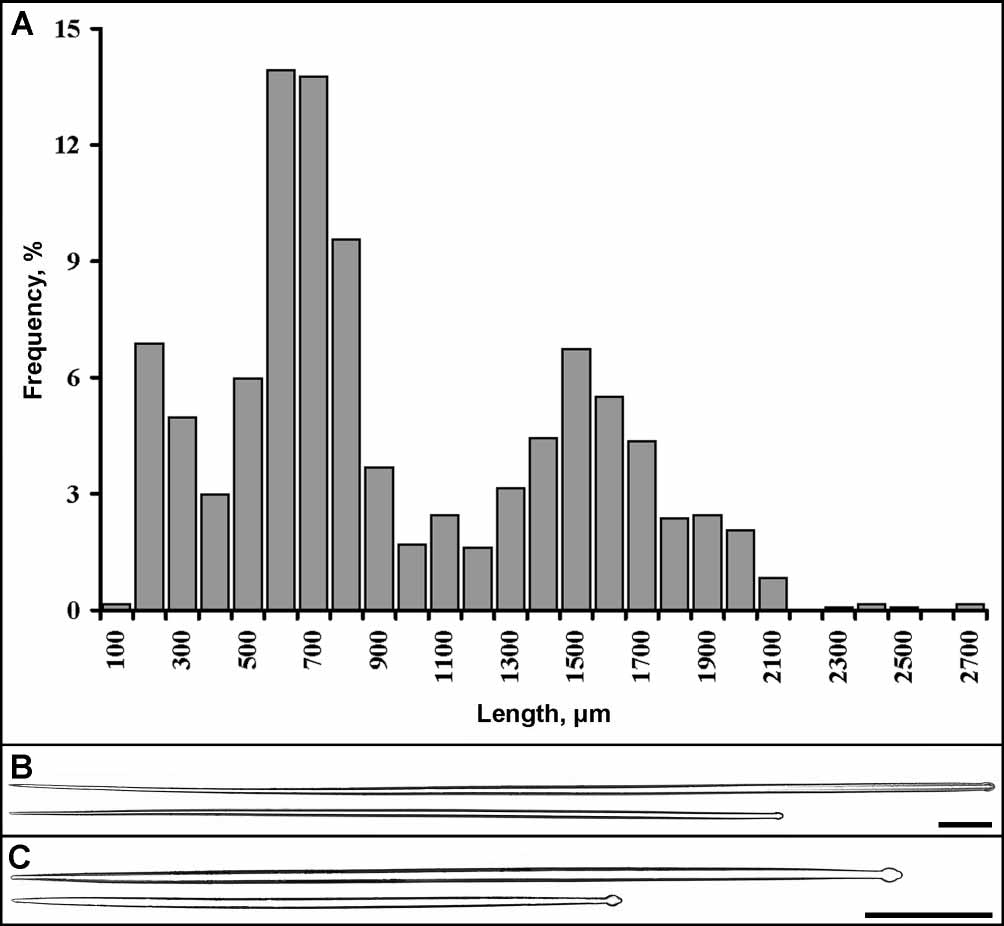
Spicules. Altogether 1307 spicules from 15 specimens were measured. Two main size categories of tylostyles are well distinguished and no intermediary class can be marked out (fig. 9A). Extralong tylostyles of the fringe undoubtedly constituted an additional size category. But the measurements of them were very few, because most of these spicules became broken on the slides. Summarizing results of spicule measurements are given in the main text below, indicating the total number of spicules measured for each category (n). Particular results of spicule measurements for each specimen (extralong tylostyles excluded) are given in table 4.
Principal tylostyles are usually straight or rarely slightly curved, slender (fig. 9B). Most of them possess terminally located, spherical tyles. More then 60% of principal spicules have welldeveloped tyles and about 35% of them have weaklydeveloped tyles. Very infrequently, principal spicules have slightly displaced, weaklydeveloped tyles or even lack any tyles. In general the principal tylostyles measure: length 957 1599 2686 µm, tyle diameter 815.027 µm, diameter of the shaft underneath the tyle 511.826 µm, maximal diameter of the shaft 817.627 µm (n= 412).
Small tylostyles are mainly straight or rarely slightly curved and most of them are slightly fusiform (fig. 9C). The majority of small spicules have terminally located, welldeveloped spherical tyles. About 11% of small tylostyles are with weaklydeveloped tyles. The slight displacement of tyle and its absence are observed very rarely. In general the small tylostyles measure: length 1256361271 µm, tyle diameter 410.021 µm, diameter of the shaft underneath the tyle 26.019 µm, maximal diameter of the shaft 311.024 µm (n= 895). It should be mentioned that in three specimens (SMF 10559 and 10564), the small spicules are significantly shorter than in other sponges—125290829 µm (n= 209).
Extralong tylostyles of the fringe are very slender as their diameters are similar to those of the principal tylostyles. Their minimal length is about 2500 µm while the maximal one exceeds 5600 µm. No remarkable differences in tyle location and development have been found between the fringe and principal tylostyles. Type locality: Antarctic: Northern Weddell Sea, ca. 2080–4800 m.
Etymology: The name arises from the type locality.
TABLE 4. Radiella antarctica sp. nov.: Results of spicule measurements. Measurements are given as in table 2.
Specimen no. Large tylostyles Small tylostyles
SMF 10558 (holotype) 13301571 1710 490676810
12.814.416 / 9.610.612 810.111.2 / 45.17.2
12.81617.6 (n= 21) 810.814.4 (n= 39) SMF 10559 (specimen 1) 10141690 2686 155275829
9.615.423 / 4.81116.2 4.110.618.9 / 2.46.211.2
8.119.927 (n= 49) 4.810.116.2 (n= 104)
SMF 10559 (specimen 2) 10641400 1710 170320660
12.814.517.6 / 6.410.412.8 5.610.217.6 / 2.45.512.8
1618.822.4 (n= 19) 49.720.8 (n= 48)
SMF 10561 (specimen 1) 13431744 2086 5578721271
10.81417.6 / 8.111.716.2 8.111.716.2 / 4.169.5
13.518.321.6 (n= 19) 8.112.120.3 (n= 41) SMF 10562 10641401 1710 460702930
8.810.813.6 / 6.49.312.8 5.68.712.8 / 2.46.19.6
12.815.317.6 (n= 60) 4.811.416 (n= 121) SMF 10563 (specimen 1) 13861523 1686 5718451114
13.516.921.6 / 10.814.116.2 8.112.316.2 / 4.17.213.5
16.21921.6 (n= 28) 8.114.324.3 (n= 32) SMF 10563 (specimen 2) 14861796 2086 6007851271
9.51317.6 / 8.11116.2 6.89.512.2 / 4.16.18.1
13.517.520.3 (n= 30) 8.111.716.2 (n= 60) SMF 10564 15431864 2543 125291520
10.81518.9 / 8.112.214.9 6.49.213.5 / 3.25.28.1
12.816.727 (n= 29) 5.48.313.5 (n= 57) SMF 10565 957 1567 2671 400591871
12.215.118.9 / 8.110.813.5 8.11013.5 / 2.75.88.1
13.518.724.3 (n= 36) 8.111.416.2 (n= 54) SMF 10566 (specimen 1) 12861712 2057 5107961178
813.627 / 6.811.825.7 6.48.720.3 / 3.25.518.9
12.816.925.7 (n= 43) 5.411.121.6 (n= 84) SMF 10567 (specimen 1) 11021606 2090 480684940
11.214.417.6 / 6.410.414.4 6.49.912.8 / 2.45.78.8
12.815.318.4 (n= 20) 811.617.6 (n= 40) SMF 10568 (specimen 1) 12161525 1748 470663870
12.815.222.4 / 9.612.816 810.812.8 / 4.85.98
12.817.622.4 (n= 21) 81012.8 (n= 43)
SMF 10568 (specimen 2) 13301594 2014 490707900
13.615.817.6 / 10.412.214.4 810.212.8 / 4.86.18
14.416.619.2 (n= 20) 7.21012.8 (n= 40) SMF 10569 13301568 1786 460649850
14.417.722.4 / 12.814.117.6 6.410.114.4 / 1.65.48
1618.422.4 (n= 26) 410.216 (n= 41)
SMF 10570 10261429 1634 420641950
12.819.122.4 / 6.414.817.6 6.49.212.8 / 2.45.49.6
1619.320.8 (n= 28) 3.29.416 (n= 54) Remarks. No species of Radiella has previously been reported from the Antarctic, although two species were described from other regions of the Southern Hemisphere, namely R. irregularis ( Ridley & Dendy, 1886) from the Chilean Coast, and R. sarsi ( Ridley & Dendy, 1886) from Australia. Both species are from the deep sea. In contrast to the Antarctic sponges, the choanosomal skeleton of R. irregularis and R. sarsi consists of a confused mass of small tylostyles, while the upper cortical skeleton is formed by intermediary spicules, and long spicules constitute the basal envelope and the marginal fringe. Radiella irregularis is also distinguished by the presence of several papillae. The status of R. sarsi is somewhat controversial because its type locality simultaneously includes Australia and the Azores. Meanwhile later findings of this species have been made exclusively in the Northern Hemisphere (BouryEsnault et al. 1994; Plotkin 2004). Another similar deepwater species, R. straticulata ( Wilson, 1925) from the Philippines, is distinguished from other Radiella spp. by the threelayered cortex.
The status of R. sol Schmidt, 1870 , the type species of the genus, is rather confused. Schmidt (1870) described it from Cuba. His illustration of the sponge habit resembles R. sarsi and also some of our Antarctic sponges, but the description is very brief and gives no information about the skeleton architecture. Boury Esnault (2002) discovered a specimen collected by the ‘Blake’ expedition 1878–1879 from the Western Central Atlantic deep waters and identified by Schmidt as R. sol . She considered this specimen as the holotype but made a question mark and emphasized that the discovered specimen differs from the original illustration of Schmidt (1870). The differences are not listed but the comparison of Schmidt’s drawing and BouryEsnault’s description and photo reveals that the former sponge had a single papilla and a hispid upper surface whereas the latter specimen possessed 15 papillae and a velvety surface. Additionally, Schmidt’s drawing indicates a very long marginal fringe whereas BouryEsnault’s photo shows a considerably damaged or probably just weaklydeveloped fringe; although in her description the fringe is alleged to be very long. Meanwhile, the possession of several papillae, the smooth or velvety upper surface and the moderatelydeveloped fringe are typical of R. hemisphaerica (Sars, 1872) , widely distributed in the North Atlantic and Arctic deep and shallow waters ( Koltun 1966; Plotkin 2004). BouryEsnault writes (2002, p.213): “…Presently it is impossible to ascertain if this specimen from the West central Atlantic is a synonym of ‘ hemisphaericum ’ from the Arctic and North Atlantic…”
Albeit a detailed revision of Radiella being necessary, it appears quite clear that the materials dealt with here belong to a new species. We should just mark out high variability of external morphology of R. antarctica sp. nov. Sponges free of substrate, or attached to very small substrata, have lenticular or discoid shape with a prominent basal surface enveloped by main spicule tracts, which is typical of other Radiella spp. On the contrary, specimens attached to larger substrata are hemispherical or cushionshaped and the enveloping of their basal surface is reduced. However, in both cases a marginal fringe of additional spicules is developed to a greater or lesser extent.
| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.