
Cephalodasys interinsularis, Kieneke, Alexander, Schmidt-Rhaesa, Andreas & Hochberg, Rick, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3947.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:41AF948D-57C2-4622-B4A5-55EF72BE1F01 |
|
DOI |
https://doi.org/10.5281/zenodo.6098435 |
|
persistent identifier |
https://treatment.plazi.org/id/03E9E710-FFC2-FFAD-FF3F-546FFF5FFCFE |
|
treatment provided by |
Plazi |
|
scientific name |
Cephalodasys interinsularis |
| status |
sp. nov. |
Cephalodasys interinsularis View in CoL sp. nov.
Figs. 2–5A View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5. A , 6 View FIGURE 6
Habitat/ type locality. The new species was collected from rather fine calcareous biogenous sand from a sublittoral shoal between Lee Stocking Island and Norman’s Pond Cay at a water depth of approximately 2 m (N 23°45.972´; W 76°06.897´). The area between both islands (Fig. 1) has the properties of a tidal channel and hence a strong tidal current was present at the time of sampling. Apart from Cephalodasys , the sample also contained specimens from the gastrotrich taxa Macrodasys , Paraturbanella , Tetranchyroderma (Macrodasyida) and Draculiciteria (Paucitubulatina) .
Material examined. Two specimens were studied alive and a third specimen was studied after fixation. Among the two live specimens, one individual was fully mature and the second was a subadult. The third specimen was fixed and studied with light and electron microscopy. Since only a single mature specimen was studied alive, this specimen is designated as the holotype even though it is no longer extant (International Code of Zoological Nomenclature/ ICZN, Articles 73.1.1 and 73.1.4, International Commission on Zoological Nomenclature 1999, 2008, 2012). The specimen prepared for SEM (see Fig. 6 View FIGURE 6 A-F) is designated as a paratype specimen ( ICZN, Recommendation 73D) and is stored in the collection of the National Museum of Natural History/Smithsonian Institution ( USNM 1274540). Digital light microscopic and SEM images of this paratype are also kept in the collection. Digital files (images and videos) of the holotype and the other live specimen (see Figs. 3 View FIGURE 3 , 4 View FIGURE 4 , 5A View FIGURE 5. A ) are also deposited at the National Museum of Natural History. The latter individual is designated as a further paratype specimen ( ICZN, Recommendation 73D), which is no longer extant. A fourth specimen of Cephalodasys that was studied alive (see Fig. 5 View FIGURE 5. A B) probably does not belong to the new species (see discussion for details).
Diagnosis. Cephalodasys with a body length of 471 µm. Body divided into pyriform head, neck, and trunk regions, the latter two inconspicuously separated from each other at the level of the pharyngeal pores that are positioned close to the posterior end of the pharynx. Caudal trunk end rounded. The rather long buccal cavity is hourglass-shaped and has a strengthened cuticular lining. Intestine/midgut linear and without obvious differentiation. Arrangement of TbA/TbVL/TbP is 2x 3/ 2x 5/11–12, respectively. TbA arise directly from the body surface, a fleshy base is not present. TbVL present in the trunk region only, TbD and ventral adhesive tubes (TbV) are absent. Unpaired ovary in a dorsolateral position in the posterior half of the trunk, pair of short and stout testes posterior to the pharyngeal pores, with short and caudally directed sperm ducts that fuse ventral to the midgut. An unpaired male gonopore is present at the site of sperm duct fusion. A frontal organ as putative sperm storing device is situated just posterior to the mature oocyte, a caudal organ is absent. Arrangement of cilia does not deviate from other Cephalodasys (see diagnosis above).
Etymology. The species name is compound from the Latin words inter (= between) and insularis (= belonging to an island) and denotes the type locality, a sublittoral sandbar between two islands. The name shall not indicate that the new species is limited to that special habitat. However, during our stay on Lee Stocking Island in 2010, we did not find the species at any other sampling station.
Description. The following description is based on the adult specimen that was studied alive and digitally documented, viz. the holotype ( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). A full overview of morphometric variability across all three studied specimens, including one subadult individual, is provided in Table 1 View TABLE 1 . It has to be stressed, however, that in particular length measurements of the fixed specimen may be erroneous due to contractions and/or shrinkage artefacts.
Habitus. The new Cephalodasys is a medium-sized macrodasyid gastrotrich with a total body length of 431– 471 µm (n=2). Like most other species of Cephalodasys , it has a distinct, pear-shaped head that is delineated from the neck region at U11 by a constriction ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 4 View FIGURE 4 A–C, 5A, 6A, C). The trunk region (approximately U35– U100) is slightly wider than the neck region (approximately U11–U35) and ends with a rounded caudum ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 5A View FIGURE 5. A ). Body widths are 47/37/42/48/51/25 µm at U 5/11/25 /50/75/94.5, respectively (see Table 1 View TABLE 1 ). The outline of the animal appears irregularly undulated, which is likely the result of the soft and folded integument ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 A– B). The body is ventrally flat but dorsally convex. In the testicular region, the trunk cross section appears to be trilobate. The cuticle is smooth and shows no ornamentations; epidermal glands per se are absent but the epidermis appears finely granular under the light microscope (e.g. Fig. 4 View FIGURE 4 A, C, E).
Adhesive tubes. The new species of Cephalodasys has adhesive tubes in an anterior (TbA), ventrolateral (TbVL), and posterior (TbP) series; ventral tubes and dorsal tubes are absent ( Fig. 2 View FIGURE 2 ). There are three TbA per side (n=3) present at the transition from head to neck regions (U10.5–U12, n=3). The tubes are of equal size (length: 7– 11.5 µm, width: 1.5–2.5 µm, n=3) and are arranged in parallel ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 A, 4C, 6C–D). Their longitudinal axis is slightly tilted toward the midline of the animal (approximately 7°, see Fig. 6 View FIGURE 6 C). If a living, anaesthetized specimen gets slight pressure from the cover slip, TbA may appear aligned parallel to the longitudinal axis of the animal (see Fig. 4 View FIGURE 4 C). TbA arise directly from the ventral body surface but there is a shallow pit anterior to them ( Fig. 6 View FIGURE 6 B).
Posterior to the TbA and inside the body, one can observe two parallel filaments per TbA that seem to anchor each adhesive tube. Each adhesive tube has a slight furrow extending down its longitudinal axis, which probably indicates the presence of a duo-gland adhesive system ( Fig. 6 View FIGURE 6 D). Terminally, each TbA has two tiny pores ( Fig. 6 View FIGURE 6 D). Between U35 and U75, there are five pairs of bilateral-symmetrically arranged TbVL (n=3). The tubes appear long and slender and measure 10–16.5 µm in length and 1–1.8 µm in width (n=3). Their insertion points are somewhat ventral in position but mainly lateral to the bands of locomotory cilia ( Figs. 2 View FIGURE 2 , 4 View FIGURE 4 E, 5A). Hence, the TbVL are hardly visible from the dorsal side. In both adults and subadults, there is a consistent pattern concerning the arrangement of TbVL: the spaces between the 1st, 2nd and 3rd tubes is about half the distance (25.5–34.5 µm) of that between tubes 3, 4, and 5 (47–60.5 µm). Caudally, there are 11–12 TbP (n=3) arranged along the posterior margin ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 A, 5A). Lengths of individual TbP vary within and between specimens, generally from 6.5–13 µm (n=3), while the width is more consistent (2–2.25 µm, n=3). One or two short cilia may arise from the distal tip of each TbP ( Fig. 4 View FIGURE 4 F).
Cilia. Locomotory cilia on the ventral side are arranged in two longitudinal columns beginning at the anterior end and extending to the ventral anus at U94.5. Posterior to the anus, there is a short uniform field of ventral locomotory cilia, i.e. both longitudinal columns fuse ( Figs. 2 View FIGURE 2 , 4 View FIGURE 4 E, 6A–C). In the neck region, the space between the two columns is closer than on the rest of the body, giving the false impression that they fuse ( Figs. 4 View FIGURE 4 C, 6C). The lateral flanks of the pear-shaped head bear additional cilia ( Fig. 6 View FIGURE 6 B) that extend around the lateral and dorsal portion of the head to create a closed ciliary ring ( Fig. 4 View FIGURE 4 A). The length of the locomotory cilia is ca. 10 µm. Dorsolaterally on the head region and along the whole trunk there are long (25–30 µm) and stiff sensory cilia (e.g. Fig. 6 View FIGURE 6 B). These cilia are present along the length of the body: five pairs of cilia are present on the head and 11–12 pairs are bilaterally distributed along the neck and trunk. Shorter sensory cilia (5–7 mm) are present on the flanks of the neck, trunk, and the dorsal side of the caudum. Approximately 25 short cilia surround the mouth opening ( Fig. 6 View FIGURE 6 B–C).
Sensory organs. Apart from the sensory cilia, there are no other obvious sensory organs (e.g., tentacles, pigmented eye spots) on the head region. However, dorsal to the pharynx there is a bilateral pattern of small vesicles ( Fig. 4 View FIGURE 4 B). These may belong to a pair of photoreceptive head sensory organs that are, for instance, also known from Cephalodasys maximus (see, e.g., Wiedermann 1995).
Digestive tract. The gut tube is divided into an anterior glandulo-muscular pharynx and a posterior intestine ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 B). The pharynx makes up 38–40% (adult specimens, n=2) of the total length of the whole gut tube ( Tables 1 View TABLE 1 and 2 View TABLE 2 ). The circular mouth opening is terminal (only inconspicuously tilted, see Fig. 6 View FIGURE 6 B) and has a diameter of ca. 10 µm. Apart from the short cilia mentioned above, there are no other structures such as longitudinal ridges or hooks that line the mouth. However, the cuticular wall of the cylindrical to slightly hourglass-shaped buccal cavity (25–30 µm long) is thickened ( Fig. 3 View FIGURE 3 B). The width of the pharynx (22 µm) is nearly equal along its whole length; it is only slightly narrower around the buccal cavity (U0–U05), and increases in width at the level of the pharyngeal pores. The position of the pores is at U31–U 35 in adult specimens (n=2). The actual pores are inconspicuous slits that lie dorso-laterally in the body wall ( Fig. 6 View FIGURE 6 B); the pharyngeo-intestinal junction is at U36–U 38 in adult specimens (n=2). The shape of the intestine is quite irregular and the diameter varies considerably along its course. In the region where a large ovum is situated in adult specimens, the intestine forms a drawn-out curve to the right side of the trunk and its diameter is slightly reduced ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 B). The intestine ends in a ventral anus at U94.5 (n=2). In this area, the terminal part of the intestine may be widened (see, e.g., Fig. 4 View FIGURE 4 D). Although no distinct regionalization of the intestine is visible, the abundance of small refractive spheres in its wall cells increases from anterior to posterior, possibly due to increased resorption processes in the midgut.
Reproductive organs. The new species is hermaphroditic with paired testes and an unpaired ovary. The paired testes are present between U36.5–U38 and U49–U50 (n=2) and are broad and club-shaped (12–13 µm wide) with short, caudally projecting vasa deferentia ( Figs. 2 View FIGURE 2 , 4 View FIGURE 4 G). The sperm ducts immediately curve medio-ventrally and specimen ID (name of folder with digital notes Lt[µm] LPh PhJIn pos. PP pos. PP pos. anus material, i.e. images and videos) [µm] [U] [µm] [U] [µm] specimen ID (name of folder with digital notes Wtat pos. TbA per pos. TbA pos. TbA L TbA material, i.e. images and videos) anus testes side [µm] [U] [µm]
[µm] plus vasa
deferenti
a [µm]
_ interinsularis _n_sp_adult mature holotype 25 172–237 3 53 U11 10 _ interinsularis _n_sp_adult_F- 100414 -2A mature (SEM and LM) paratype 27 165–212 3 49 U11 7
_ interinsularis _n_sp_subadult developing testes visible, no ovary paratype 27 164–209 3 44.5 U12 11.5
Cephalodasys _sp_subadult no testes, no ovary different species 32 n. a. 3 63 U10.5 10.5
… …continued on the next page specimen ID (name of folder with digital notes WTbA TbVL L TbVL W TbVL TbP L TbP material, i.e. images and videos) [µm] per side [µm] [µm] [µm)
interinsularis _n_sp_adult mature holotype 2–2.5 5 13.5– 1.8 11 6.5–10.5
16.5
interinsularis _n_sp_adult_F- 100414 -2A mature (SEM and LM) paratype 1.5 5 10–11.5 1 11 n. a. interinsularis _n_sp_subadult developing testes visible, no ovary paratype 2 5 12 –14 1.7 12 7–13
Cephalodasys _sp_subadult no testes, no ovary different species 2.5 4 10–12 1.7 11 7–10
specimen ID (name of folder with digital notes W TbP [µm] ratio of LPh to total pos. testes plus vasa deferentia material, i.e. images and videos) digestive tract [U]
interinsularis _n_sp_adult mature holotype 2.25 38% U36.5–U50 interinsularis _n_sp_adult_F- 100414 -2A mature (SEM and LM) paratype 1.5–2 40% U38–U49 interinsularis _n_sp_subadult developing testes visible, no ovary paratype 2 46% U43–U55
Cephalodasys _sp_subadult no testes, no ovary different species 2.25 37% n.a.
PhJIn total digestive
tract a
… …continued on the next page …….continued on the next page
…….continued on the next page Cephalodasys sp. nov. shoal close to Lee Stocking Island, Bahamas (N 23°45.972´; fine calcareous sand 2 m this study
W 76°06.897´) (Caribbean Sea)
this ratio has been re-measured from the drawings of the original descriptions
'common' means that TbA are comparably wide as other tubes of the animal
considered are TbVL anterior to the pharyngeal pores
termination of vasa deferentia was not observed
description based on protogynous specimens
posterior termination of testes in C. littoralis is unsure
description based on subadult specimens
join in an unpaired ventral male gonopore ( Figs. 4 View FIGURE 4 E, 6E). Based on light microscopic observations, the male gonopore appears as a simple circular opening ( Fig. 4 View FIGURE 4 E). However, scanning electron microscopy of that region indicates a slightly more complex, valve-like structure ( Fig. 6 View FIGURE 6 E). Furthermore, light microscopy reveals a trefoilshaped structure directly underneath the male gonopore, possibly a glandular organ that aids sperm transfer in that species (indicated in the schematic drawing, see Fig. 2 View FIGURE 2 ). At the anterior pole and along the median wall of each testis, cell-like compartments with granular content can be observed in some optical layers ( Fig. 4 View FIGURE 4 G). We interpret this as early germ cell stages. According to this interpretation, spermatogenesis proceeds from posterior to anterior and spermiogenesis medio-laterally. A single mature egg is present dorso-lateral to the intestine and on the left side of the trunk between U65 and U85. The egg contains numerous small spheres, probably yolk material, and has a large globular nucleus, ca. 15 µm in diameter ( Fig. 3 View FIGURE 3 B). Smaller oocytes with ovoid nuclei (6 x 12.5 µm) are present anterior the single large egg. Hence, oogenesis proceeds antero-posteriorly. Posterior to the mature egg on the left side of the posterior trunk (between about U82.5 and U88.5) there is an accessory reproductive organ, viz. the frontal organ. The shape of this structure is somewhat ovoid but irregular and lacks any obvious musculature. In its center, the putative organ lumen, there is a grain-like inclusion and few filiform structures coiled around it ( Figs. 3 View FIGURE 3 B, 4D). We interpret these structures as foreign spermatozoa of a mating partner. The putative wall epithelium of the frontal organ is probably secretory since some small vesicles can be observed. Light and SEM observations indicate a lateral external pore to the frontal organ ( Figs. 4 View FIGURE 4 D, 6F); an internal pore was not observed. Apart from the structure associated with the male gonopore, no further accessory reproductive organ (i.e. a caudal organ) is present.
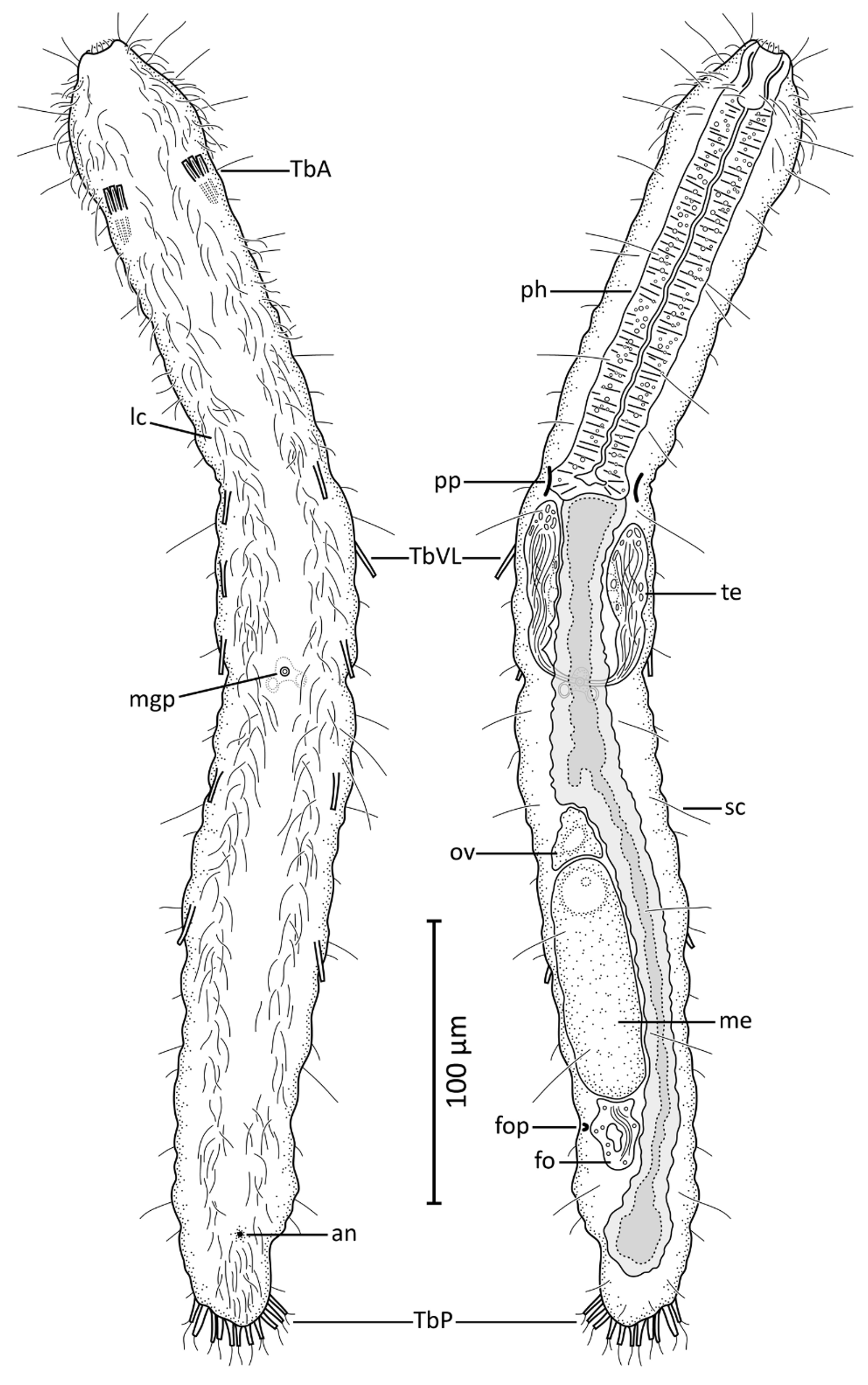

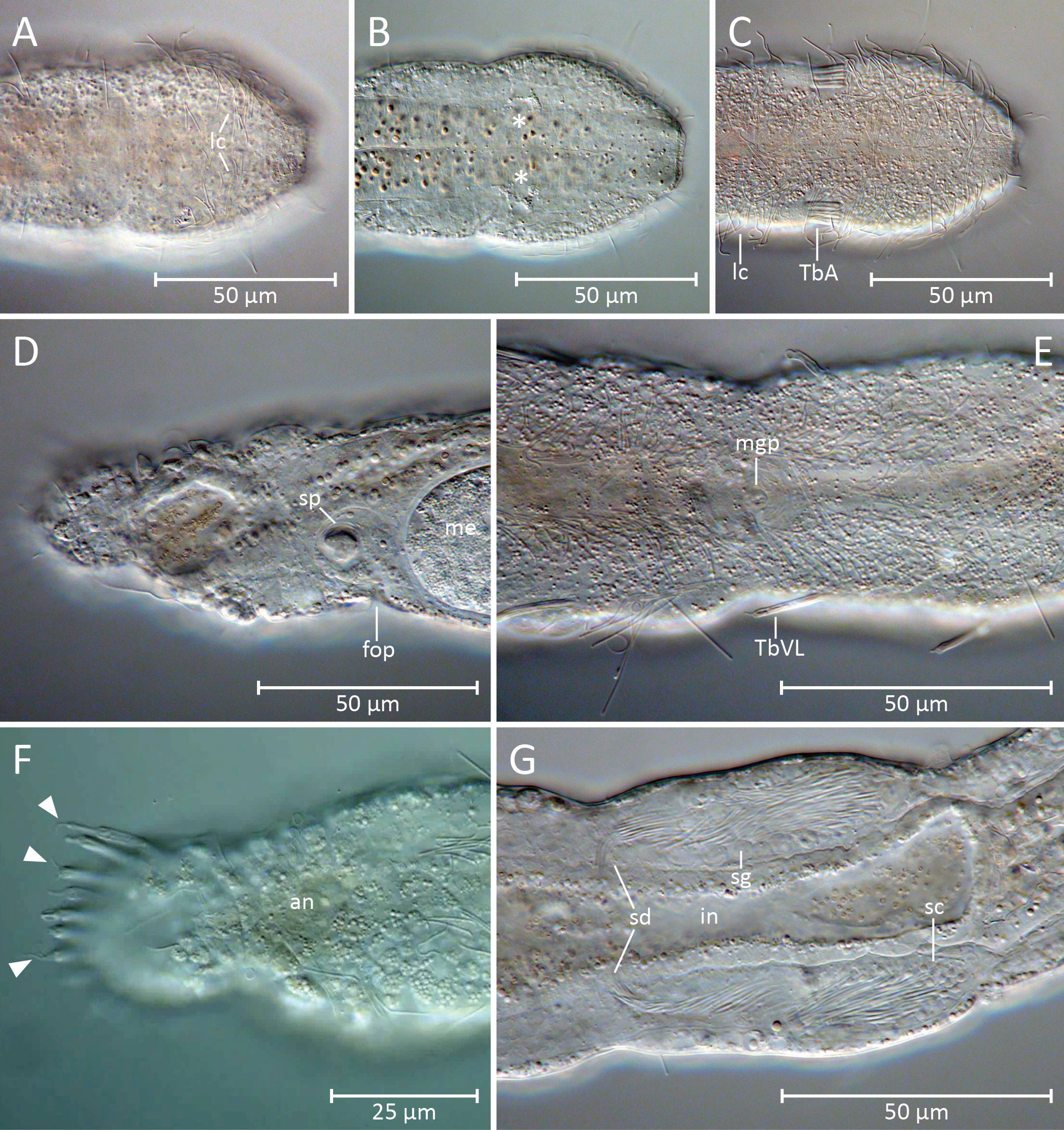
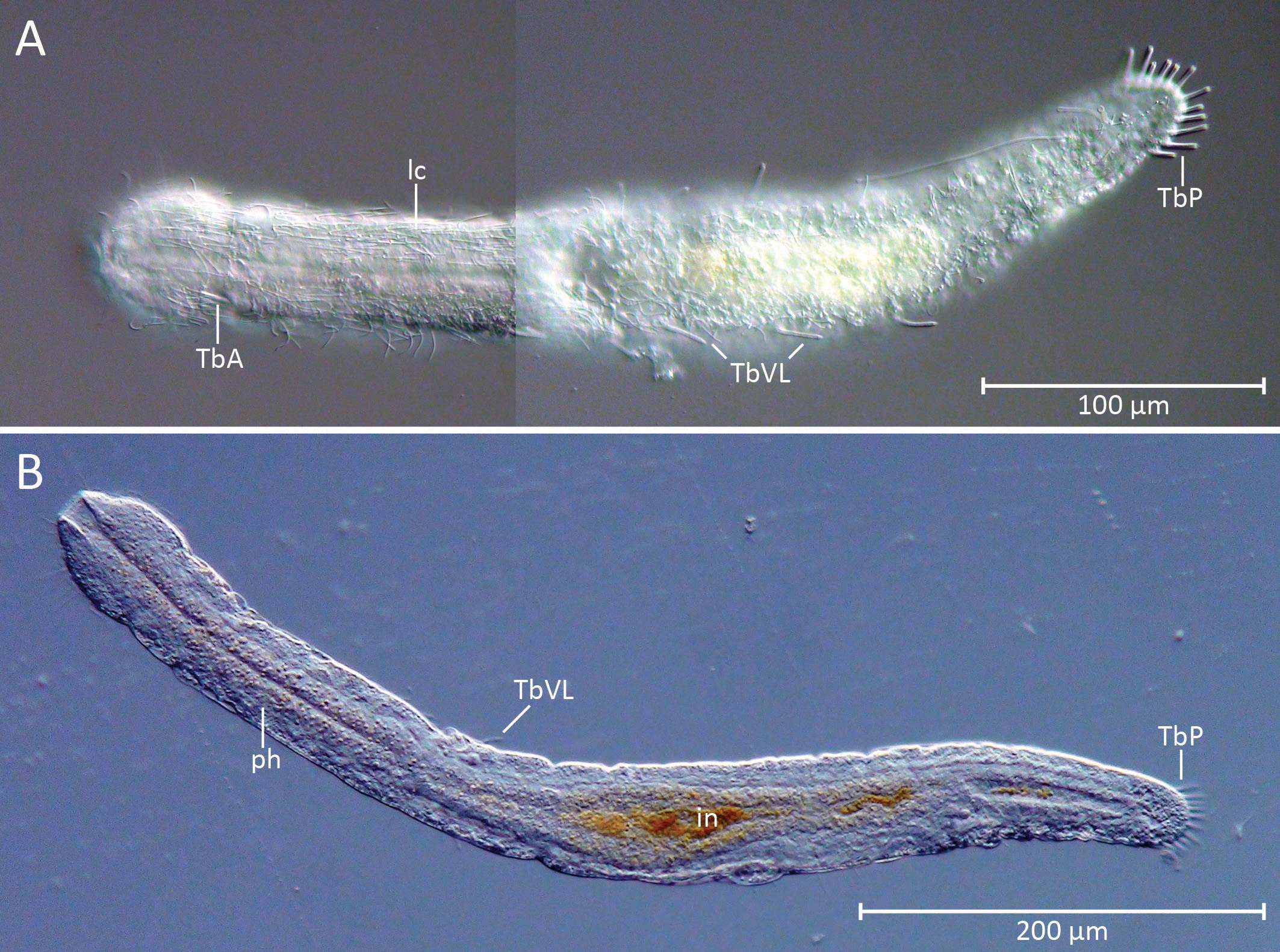
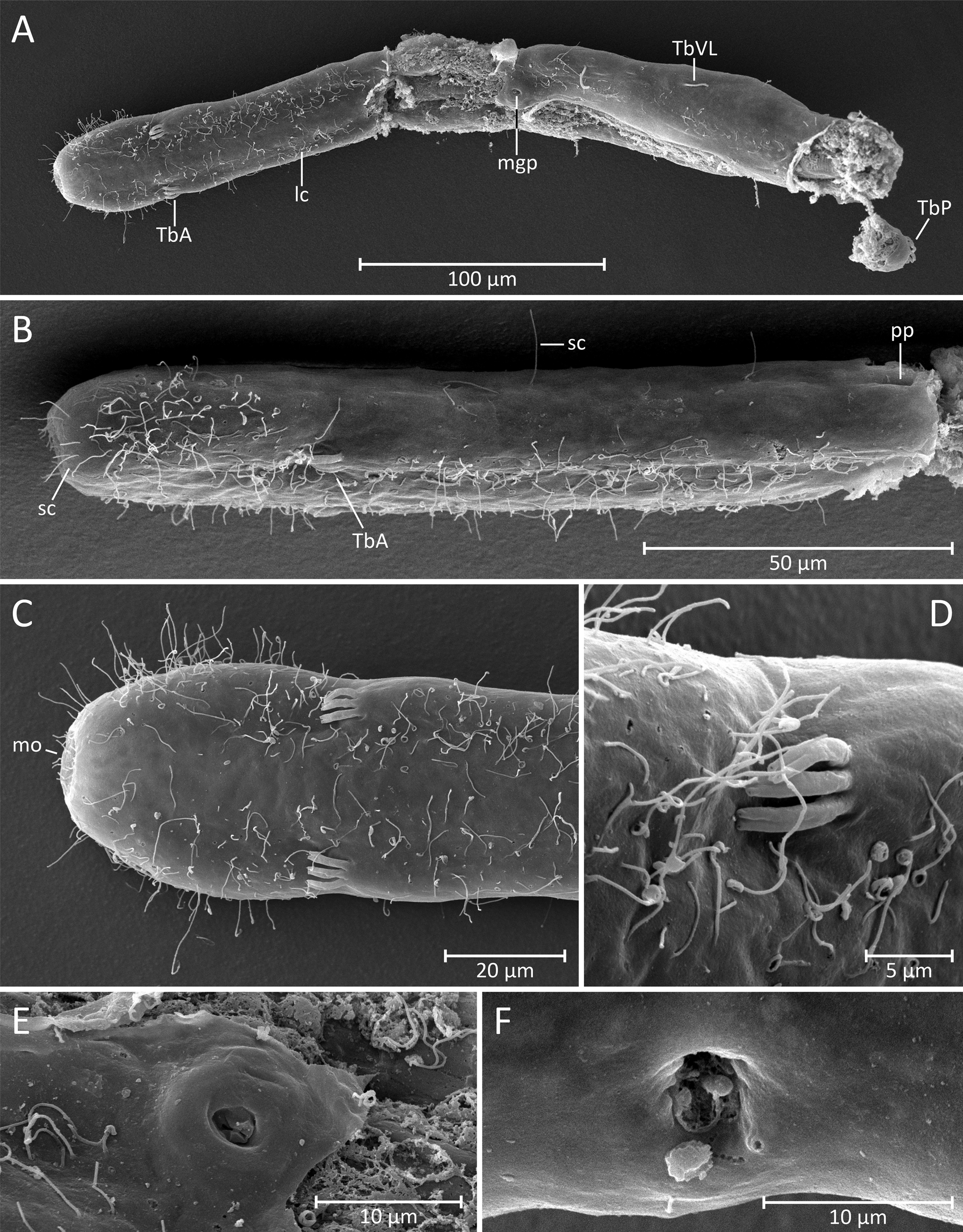
TABLE 1. Meristic and morphometric characters of the four investigated specimens of the new and one undescribed species of Cephalodasys from the Bahamas. Abbreviations: L, length; LPh, length of pharynx; Lt, total body length; n. a., not applicable; pos., position; PP, pharyngeal pores; PhJIn, pharyngeo-intestinal junction; TbA, anterior adhesive tubes; TbP, posterior adhesive tubes; TbVL, ventrolateral adhesive tubes; W, width; Wt, trunk width.
| C_ interinsularis _n_sp_adult | mature holotype | 471 | 171 | U36 | 164 | U35 | 445 |
|---|---|---|---|---|---|---|---|
| C_ interinsularis _n_sp_adult_F-100414-2A | mature (SEM and LM) paratype | 431 | 165 | U38 | 135 | U31 | 408 |
| C_ interinsularis _n_sp_subadult | developing testes visible, no ovary paratype | 378 | 163 | U43 | 153 | U40.5 | 355 |
| Cephalodasys _sp_subadult | no testes, no ovary different species | 603 | 215 | U35.5 | 199 | U33 | 586 |
TABLE 2. Meristic and morphometric characters with diagnostic value of all known species of Cephalodasys. Abbreviations: a, absent; IT, intertidal; n. a., not applicable; p, present;?, unknown character; the question mark in brackets (?) denotes an unsure character state. Further abbreviations: See Table 1. species Lt [µm] LPh [µm] position of ratio of LPh to shape of anterior end (" head ") shape of caudum
| Cephalodasys cambriensis | 990 | 290 | U30 32% | pyriform rounded disc |
|---|---|---|---|---|
| Cephalodasys caudatus | 700–800 | 220 | U33 39% | rhombic, Dactylopodola -like elongate lobe |
| Cephalodasys dolichosomus | 615–772 | 205–215 | U28–U33 29% | pyriform rounded, slightly flared |
| Cephalodasys hadrosomus | 322 | 152 | U47 53% | rounded rounded |
| Cephalodasys littoralis | 600–650 | 200 | U35.5 39% | pyriform tapering |
| Cephalodasys maximus Cephalodasys miniceraus | 700 464g | 235 138 | U33.5 36% U32 36% | pyriform rounded disc with hornlike lobes, Turbanella -like tapering |
| Cephalodasys pacificus | 294–368 | 105–125 | U34–U37 37% | pyriform rounded, slightly flared |
| Cephalodasys palavensis | 400–500 | ca. 140 | U38 44% | rhombic, Dactylopodola -like tapering with blunt apex |
| Cephalodasys saegailus | 492–517 | 191–196 | U38–U39 37% | pyriform rounded, slightly flared |
| Cephalodasys swedmarki | 500 | 238 | U47 51,5% | pyriform to ovoid rounded |
| Cephalodasys turbanelloides | 1000 | 367 | U36.5 38% | pyriform rounded to parable-shaped |
| Cephalodasys sp. nov. | 431–471 | 171–215 | U35.5–U36 38–40% | pyriform rounded |
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.