
Ootheca
|
publication ID |
https://doi.org/ 10.5281/zenodo.276235 |
|
DOI |
https://doi.org/10.5281/zenodo.6209545 |
|
persistent identifier |
https://treatment.plazi.org/id/03EA500A-FF87-C956-84F1-FCC7FC91DD6F |
|
treatment provided by |
Plazi |
|
scientific name |
Ootheca |
| status |
|
Redescription of Ootheca
Ootheca Chevrolat, 1837: 402 .
= Ergana Chapuis, 1879: 27 ; Seeno & Wilcox 1982: 111. Type species: Crioceris mutabilis Sahlberg, 1829 ; in Chevrolat (1837: 402) by monotypy; later designated by Chevrolat (1849).
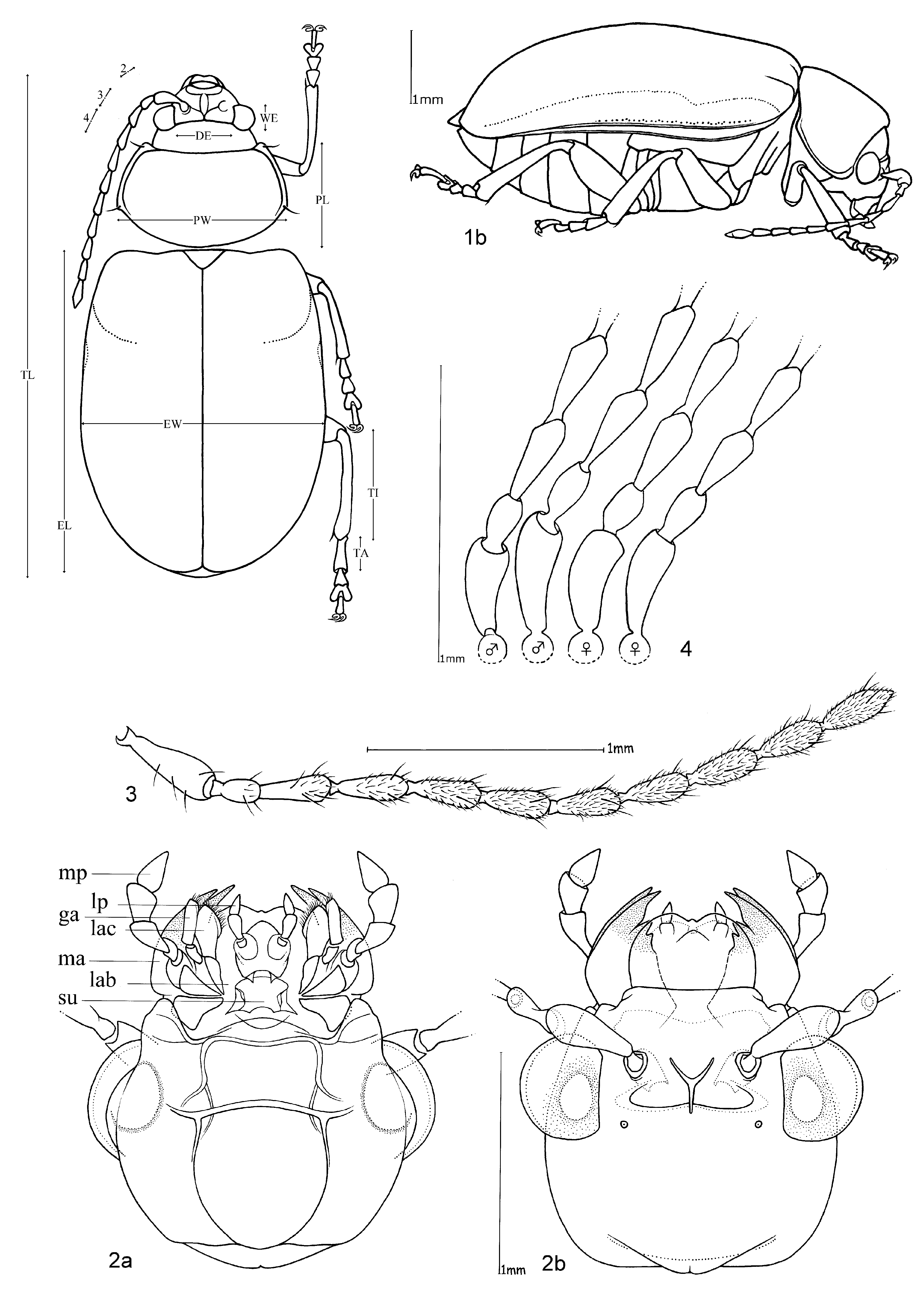
Total length. 3.60–6.80 mm; means of total length for the species range between 4.19–5.98 mm. Females are somewhat larger than males.
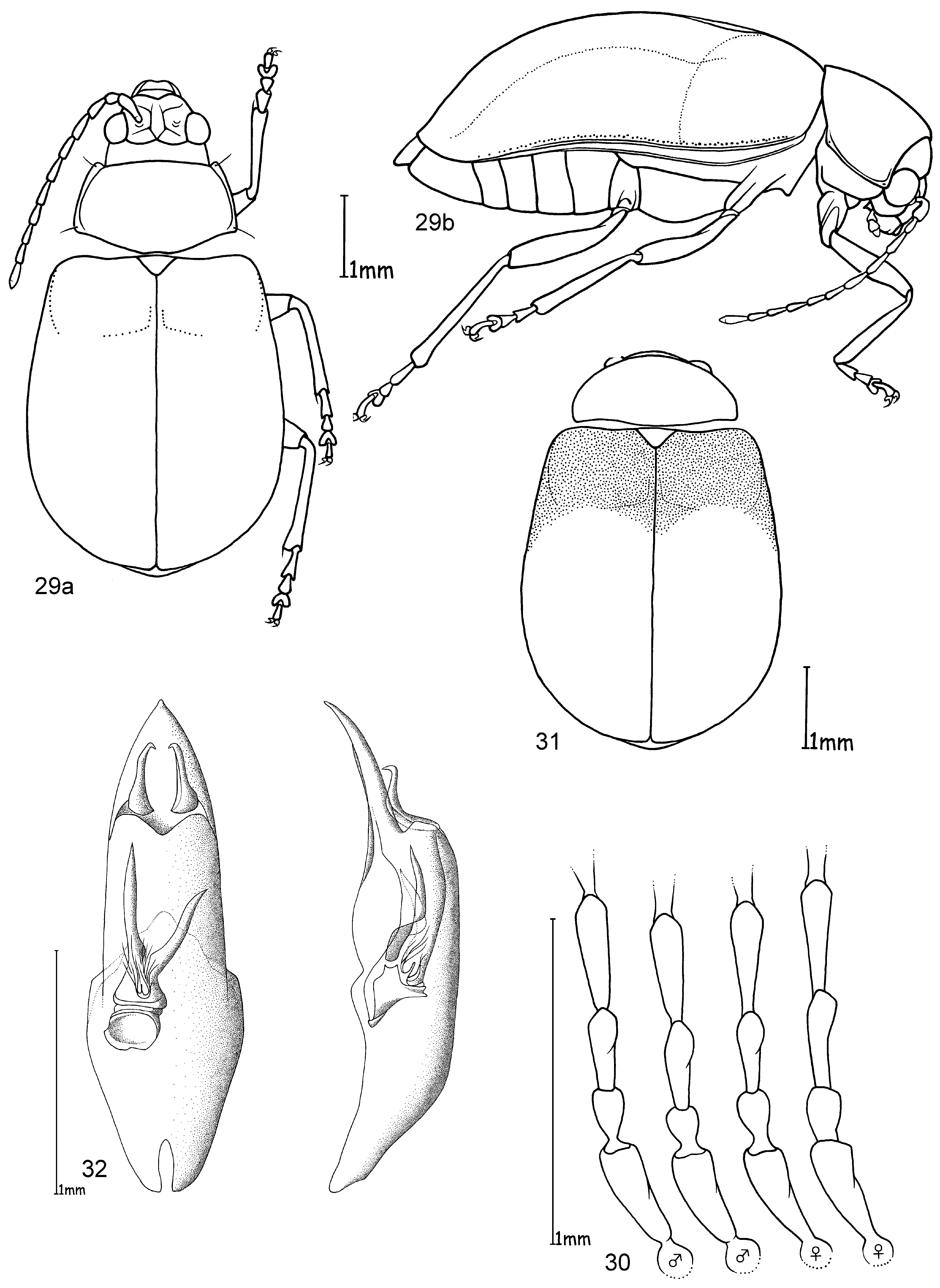
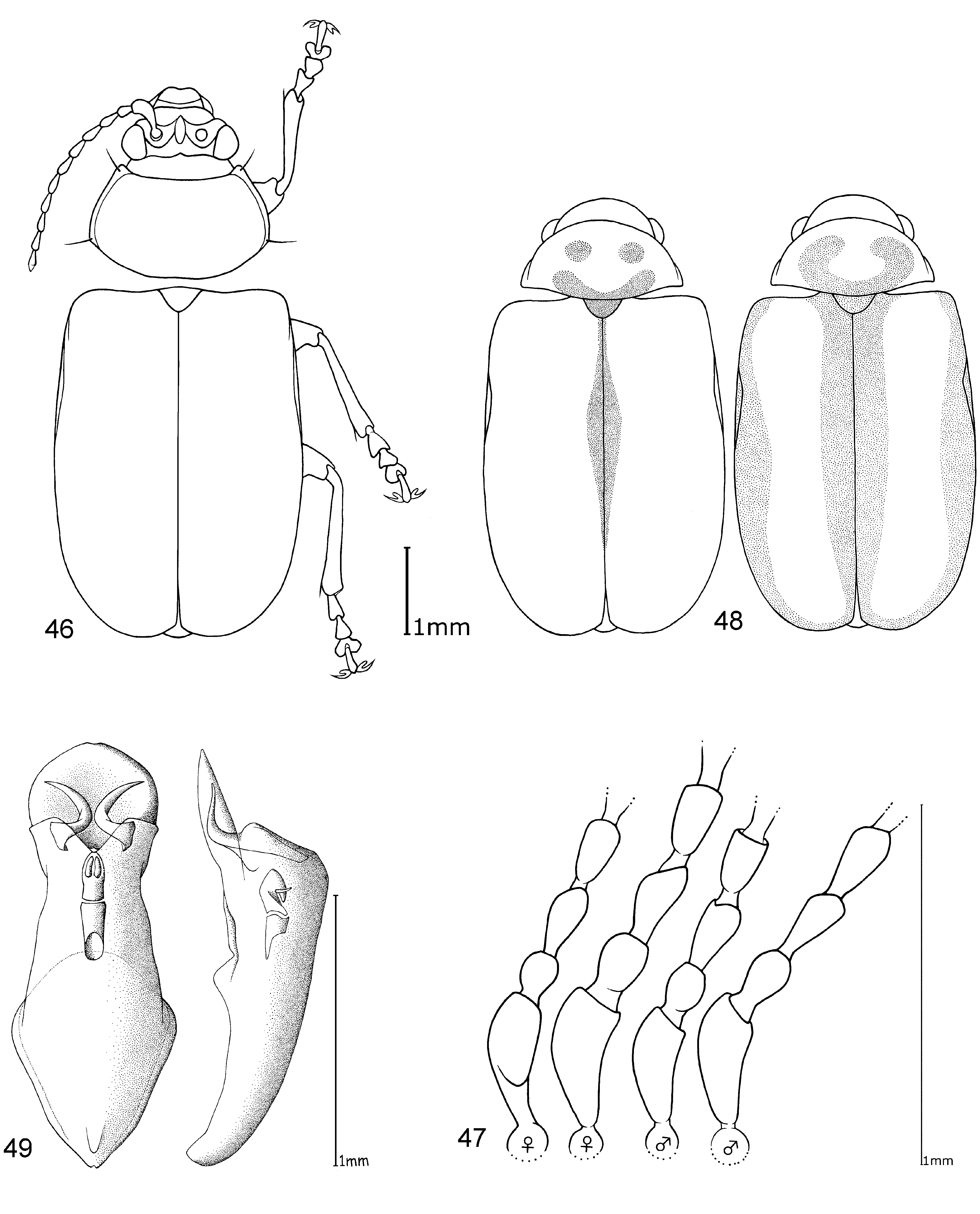
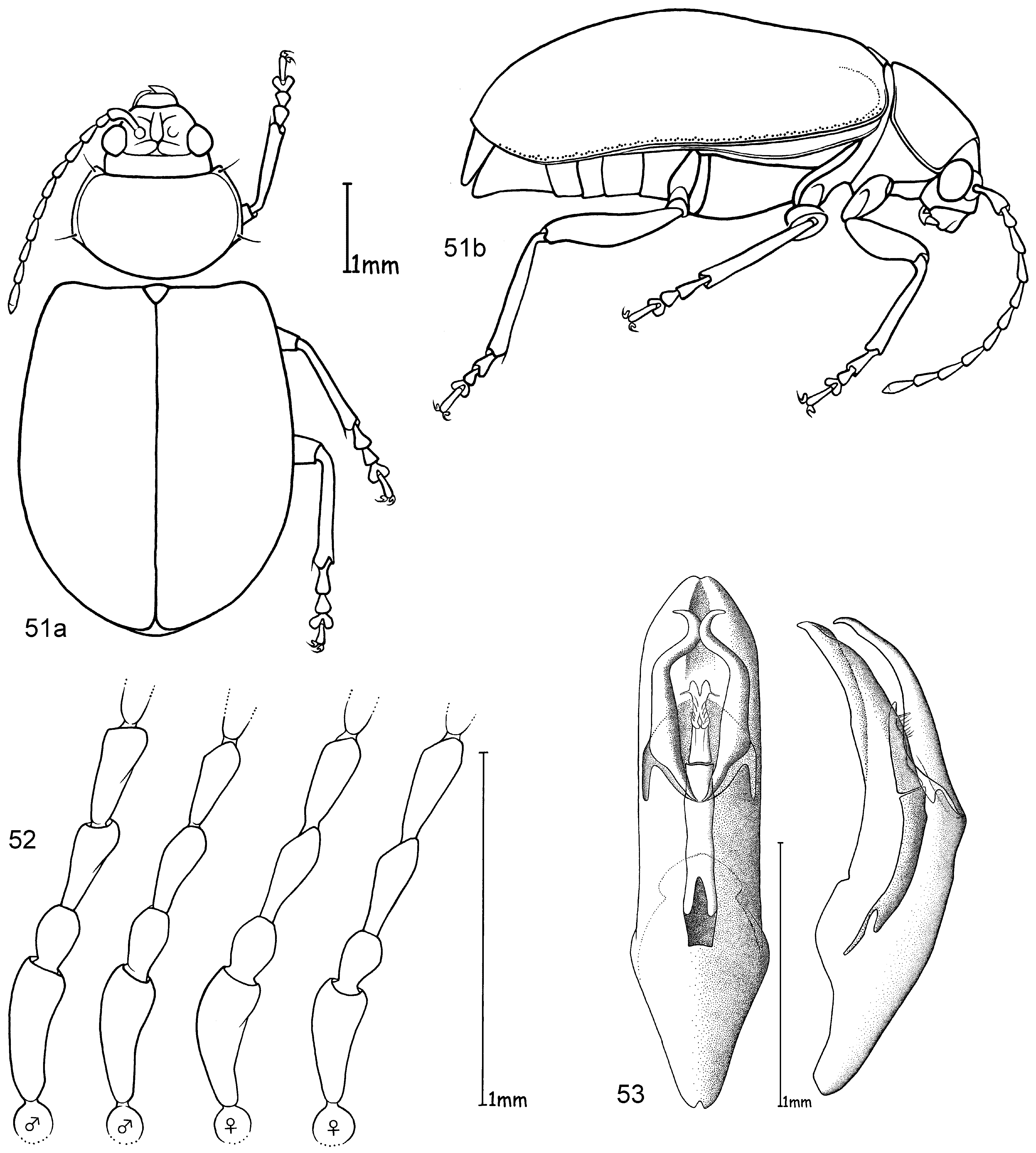
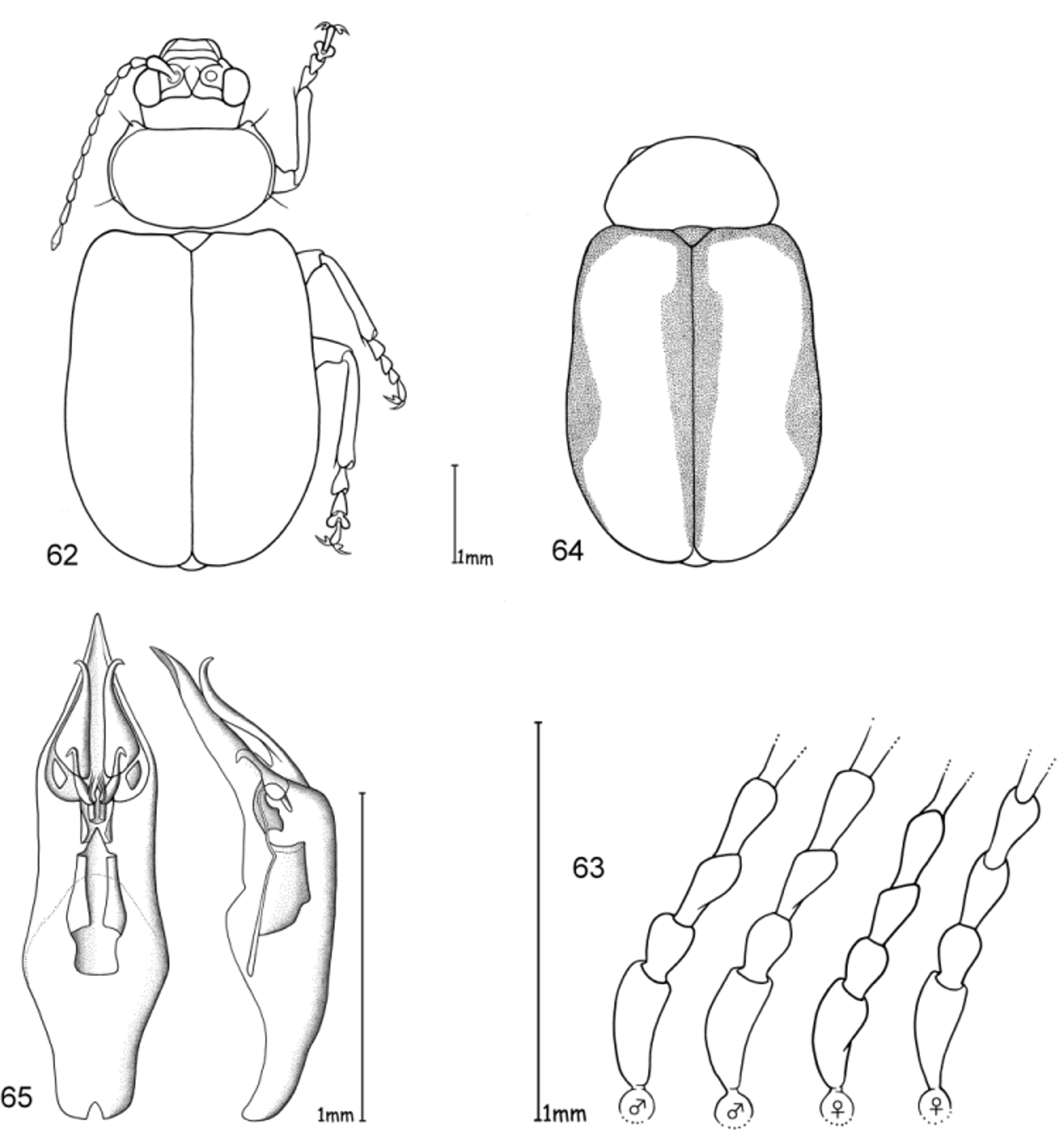
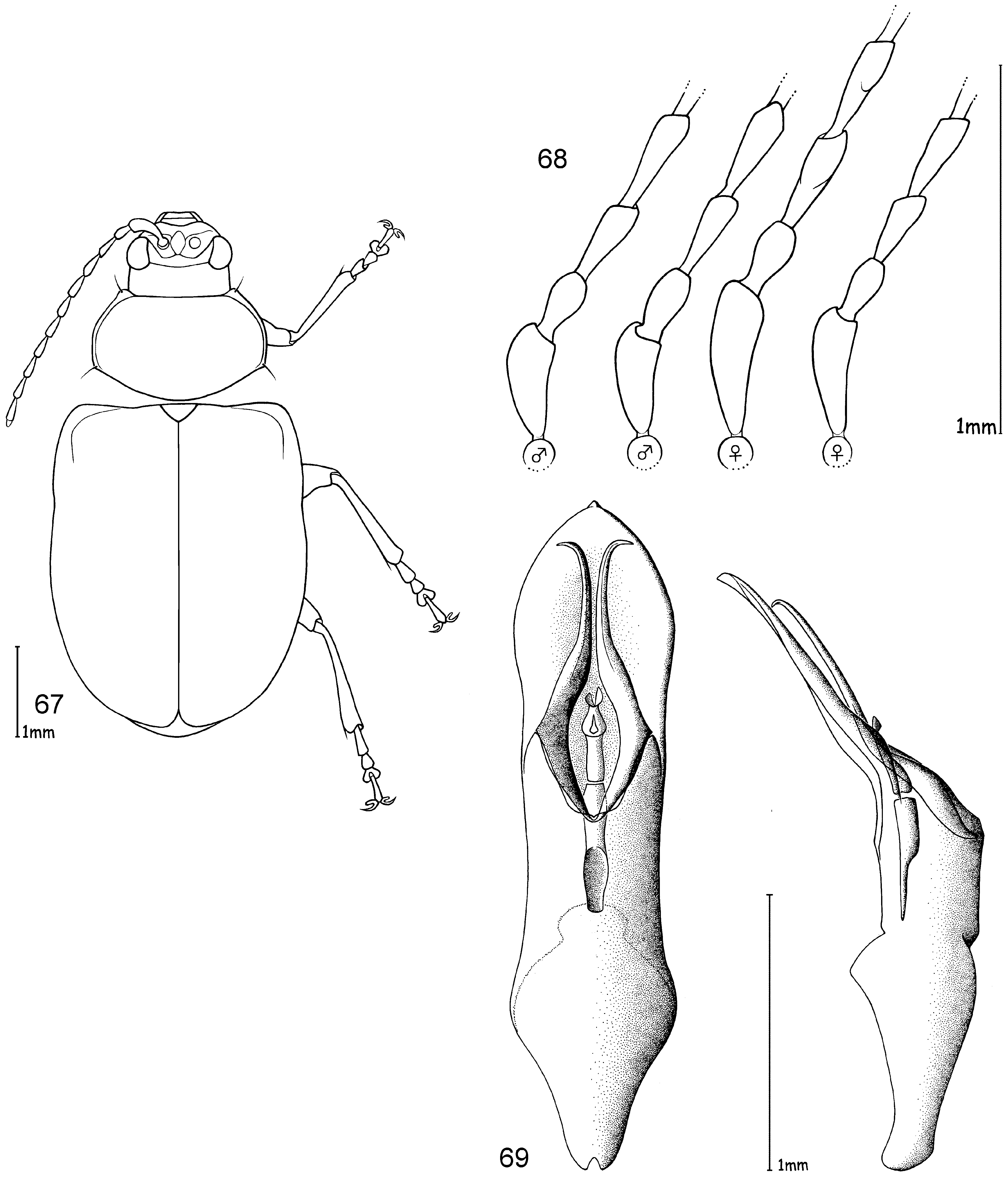
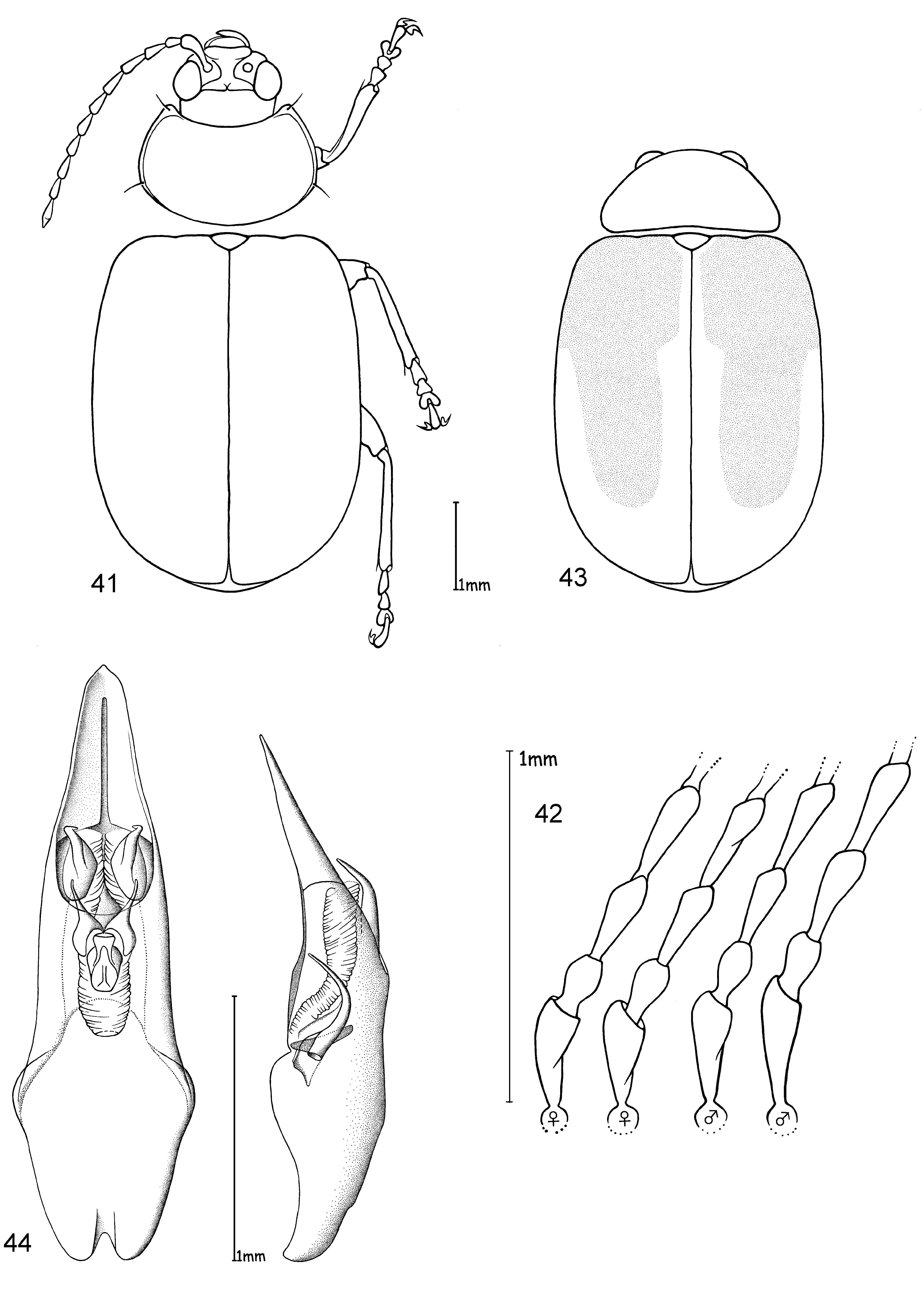
Head. Pale yellowish to nearly red, brown or black. Maxillary palp yellowish-red, brown or black, last palpomere conical ( Fig. 2 View FIGURES 1 – 4 ). Eyes: convex and ovate ( Fig. 2 View FIGURES 1 – 4 b); a nearly straight, depressed line between the posterior margins of eyes; means of ratios for maximal eye width to minimal distance between eyes vary from 0.50 to 0.69 between species. Labrum yellow to brown or black. Basal antennomeres usually yellowish, from either the fourth or fifth antennomere to terminal antennomere gradually becoming dark-brown to black towards antennal apex, antennae in a few species entirely yellowish or black; mean antennal length to total body length ratio 0.47–0.61; mean length ratio of antennomeres two to three 0.59–0.88 mm; mean length ratio of antennomeres three to four 0.76–0.93 mm ( Figs 3, 4 View FIGURES 1 – 4 ).
Thorax. Prothoracic coxal cavities closed ( Fig. 5 View FIGURES 5 – 9 b). Pronotum convexly rounded dorsally, lateral margins distinctly demarcated from disc, anterior angles rounded or protrudent in some species ( Fig. 5 View FIGURES 5 – 9 ); some species, particularly large specimens, with slightly impressed inwardly curved line beginning near to the posterior pronotal angle to the basal third of pronotum, parallel with lateral margins, this fine line defines a slight elevation on the pronotal disc; finely punctuated, mostly a unicolourous pale yellow, reddish or black, with two species displaying a bicolourous pattern; length 1.00– 1.47 mm; width 1.70–2.58 mm; pronotal width to length ratio 1.60–1.83 ( Fig. 1 View FIGURES 1 – 4 ). Elytral colouration yellowish to reddish, brown or black, mostly unicolourous, some species with a pattern (e.g. Fig. 12 View FIGURES 10 – 12 ); coarsely and more deeply punctuated than pronotum; punctuation mostly irregular, only O. chapuisi and O. frontalis show a partly seriate punctuation on elytral disc; mean length 3.36–4.72 mm; mean width 2.64–3.76 mm; mean width of elytra together to length of elytron ratio 0.71–0.82. Scutellum obtusely / nearly triangular, yellow to black. Meso- and metathorax from yellow to black. Meso- and metasternum very broad ( Fig. 6 View FIGURES 5 – 9 ). Alae fully developed ( Fig. 7 View FIGURES 5 – 9 ). Legs either unicolourous in all variations of brown, reddish to nearly black, or the tibial apex and tarsi darker than femur; four species show paler brownish femoral apices and /or tarsomeres; mean length of metatarsus to length of metatibia ratio 0.21–0.25; claws appendiculate ( Figs 1 View FIGURES 1 – 4 , 8 View FIGURES 5 – 9 ).
Abdomen. Five visible ventrites (seven sternites, usually the same colour as the meso- and metathorax. Only two species with different colours in meso-/metathorax and abdomen: O. chapuisi has much darker abdomen than metasternum and O. julianae has much paler abdomen than metasternum. Apical visible sternite in females evenly rounded, males have two deep incisions in outer apical margin ( Fig. 9 View FIGURES 5 – 9 ).
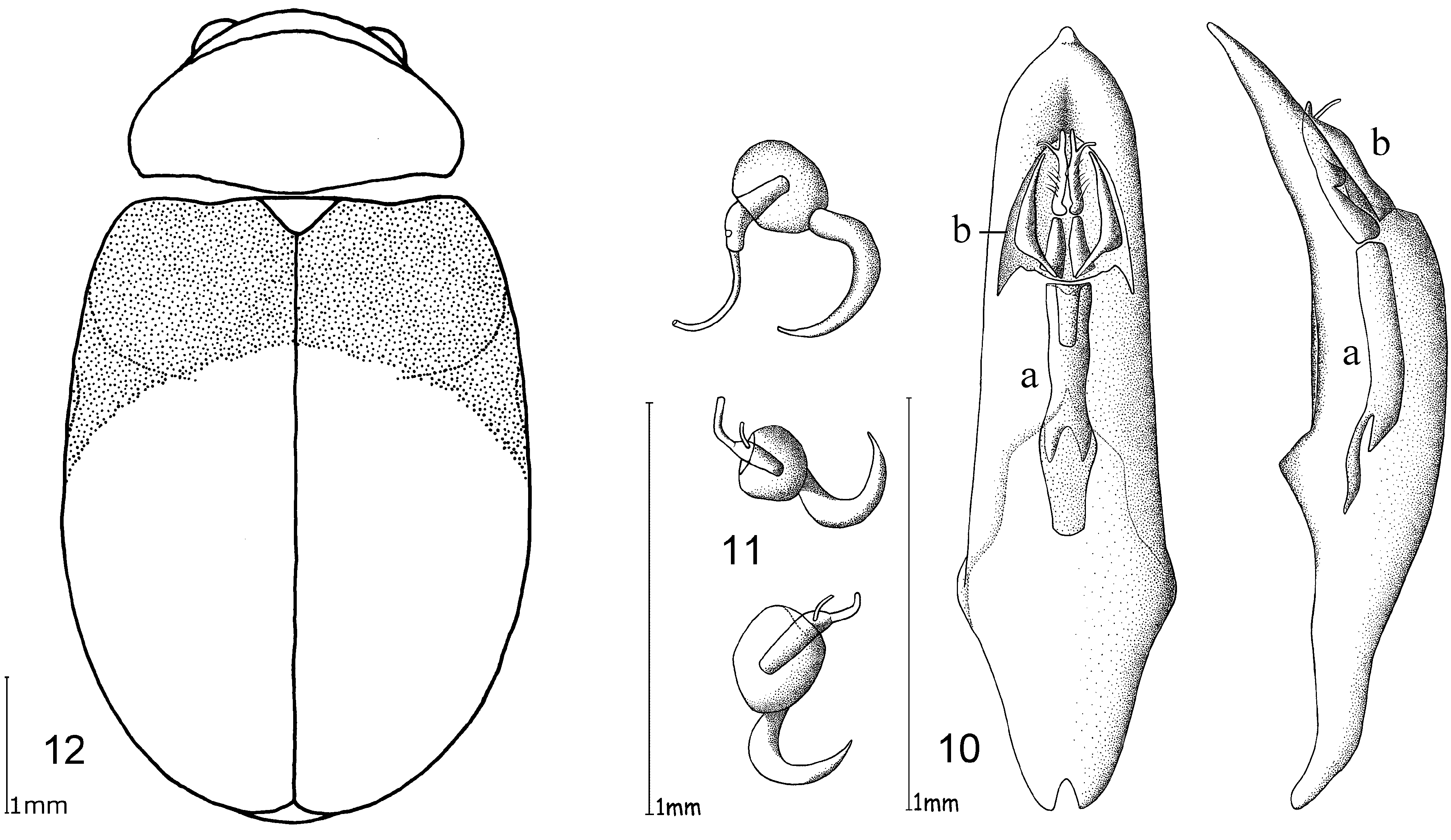
Male genitalia. Median lobe, tectum and endophallus are symmetrical (e. g. Fig. 10 View FIGURES 10 – 12 ), except O. orientalis which has asymmetrical endophallic armature. Median lobe slightly enlarged apically, parallel-sided, or strongly conical towards apex, and in lateral view slightly curved ventrally. Median lobe in three species more or less deeply incised at apex, in all other cases not incised, but rather rounded or slightly pointed. The tectum is usually divided into two tectorial spurs, with apical hook-like structures in some species. Endophallus slender in most cases with a tube-like basal sclerite and apical endophallic brush; some species with larger endophallic spines. Orifice broad and ovoid.
Female genitalia. Spermatheca with spherical nodulus. Spermathecal ductus slightly sunken within the nodulus, cornu slender, evenly and strongly curved ( Fig. 11 View FIGURES 10 – 12 ).
Diagnosis. Ootheca are Galerucinae with strongly convex, nearly parallel-sided elytra, and pronotum only slightly narrower than elytra, giving the body an elongate-ovate shape ( Fig. 1 View FIGURES 1 – 4 ).
The head is very broad with widely separated, strongly protruding eyes. Large, triangular frontal calli separate the eyes, with a distinct depressed line at their base between the posterior ocular margins. Legs are short and broad, the basi-tarsomere slightly enlarged and the claws appendiculate ( Figs 1 View FIGURES 1 – 4 , 8 View FIGURES 5 – 9 ). Antennae are short, the third antennomere about one third longer than second, and the fourth antennomere one third longer to equal length of the third ( Figs 3, 4 View FIGURES 1 – 4 ).
Colouration is either unicolourous yellowish- to reddish-brown, in most specimens bicolourous with dark brown to black elytra, rarely with black longitudinal elytral stripes. It is not easy to distinguish members of this genus from some other genera by external characters. Most similar are some species of Exosoma Jacoby, 1906 which can usually be differentiated by a more dorso-ventrally compressed body, and a more depressed pronotum with nearly rectangular posterior angles, while those in Ootheca are less angular and more rounded ( Fig. 5 View FIGURES 5 – 9 ).
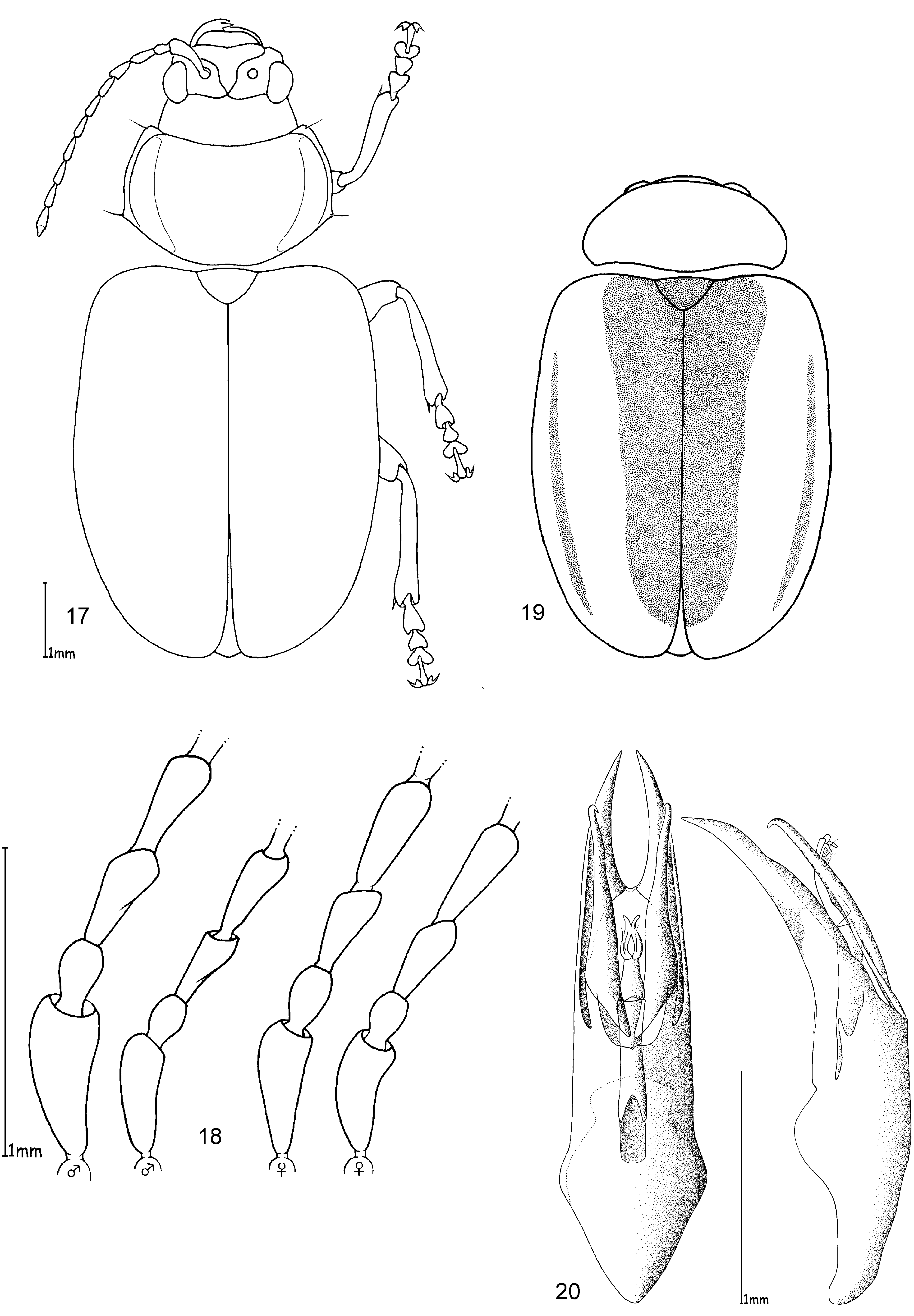
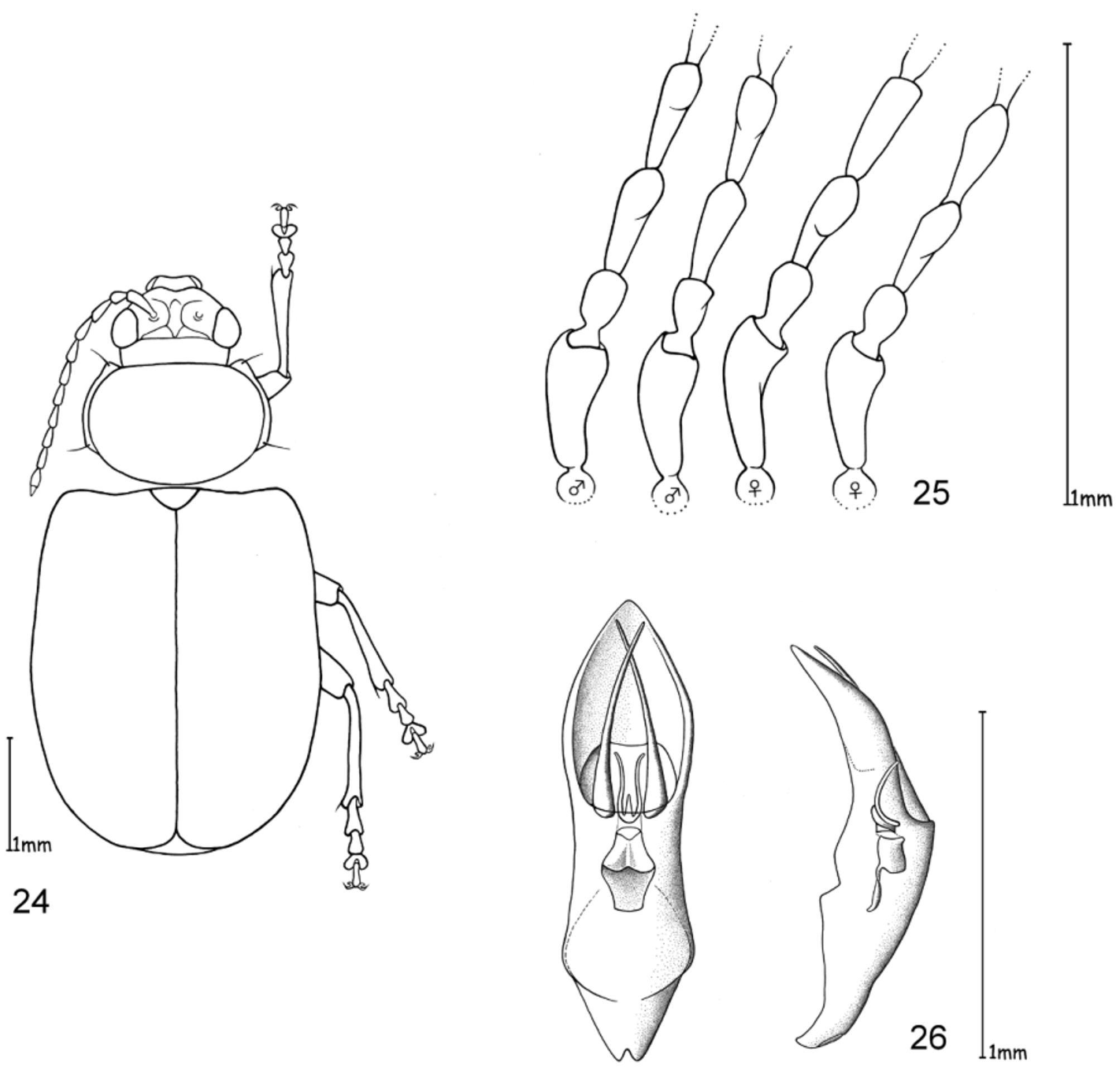
The only reliable method to distinguish members of this genus is by dissection of the male genitalia. The median lobe is short, less than four times longer than its maximal width in dorsal or ventral view. The basal quarter to third shows a very wide opening (e. g. Figs 10 View FIGURES 10 – 12 , 20 View FIGURES 17 – 20 , 26 View FIGURES 24 – 26 , 32 View FIGURES 29 – 32 , 49 View FIGURES 46 – 49 ). The median lobe is usually parallelsided ( Figs 10 View FIGURES 10 – 12 , 37 View FIGURES 35 – 37 , 53 View FIGURES 51 – 53 , 65 View FIGURES 62 – 65 , 69 View FIGURES 67 – 69 , 73 View FIGURES 71 – 73 ), slightly conical towards the apex ( Figs 20 View FIGURES 17 – 20 , 32 View FIGURES 29 – 32 , 44 View FIGURES 41 – 44 , 55 View FIGURE 55 , 59 View FIGURES 57 – 59 ) or constricted in the middle ( Figs 26 View FIGURES 24 – 26 , 49 View FIGURES 46 – 49 ). The apex can either be pointed without an apical incision, or has a deep u-shaped apical incision ( Figs 20 View FIGURES 17 – 20 , 37 View FIGURES 35 – 37 , 55 View FIGURE 55 ). The median lobe in lateral view is more or less evenly bent ventrally, or the apical third is bent at an angle to the basal part. Very characteristic for Ootheca is the tectum, which is specifically modified. It consists of two dorsal, more or less slender, often apically hooked tectorial spurs that are attached at the base of the apical third of the median lobe and extend from its middle sometimes to near the apex. These moveable spurs surely play an important role during copulation. The apical part of the endophallus can usually be seen distinctly between the base of the tectorial spurs. While males can be reliably identified utilising their genital structures, spermathecae in females are not distinctive at species level. In sympatric species a reliable identification of females is sometimes not possible without males from the same population. Weise (1900) was already aware of the strong morphological ‘overlap’ between species and wrote that the differentiation of his newly described O. orientalis and O. bennigseni was only possible by studying the male genitalia.
Host plants and biology. Food plants are known for three species and the life history was studied for O. mutabilis ( Ochieng 1977; see details in the redescription of this species below). The beetles feed on different species of Fabaceae , including trees like Erythrina and herb-like crops such as beans and ground nuts. The larvae feed on the roots, and the adults on the leaves.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Genus |
Ootheca
| Kortenhaus, Sabine & Wagner, Thomas 2010 |
Ergana
| Seeno 1982: 111 |
| Chapuis 1879: 27 |
| Chevrolat 1837: 402 |
Ootheca
| Chevrolat 1837: 402 |