
Rauisuchus tiradentes, VON HUENE, 1938
|
publication ID |
https://doi.org/ 10.1111/zoj.12196 |
|
persistent identifier |
https://treatment.plazi.org/id/03EA6076-FF9F-441C-FF4A-9FBBA696FE1C |
|
treatment provided by |
Felipe |
|
scientific name |
Rauisuchus tiradentes |
| status |
|
RAUISUCHUS TIRADENTES VON HUENE, 1938 A
Lectotype
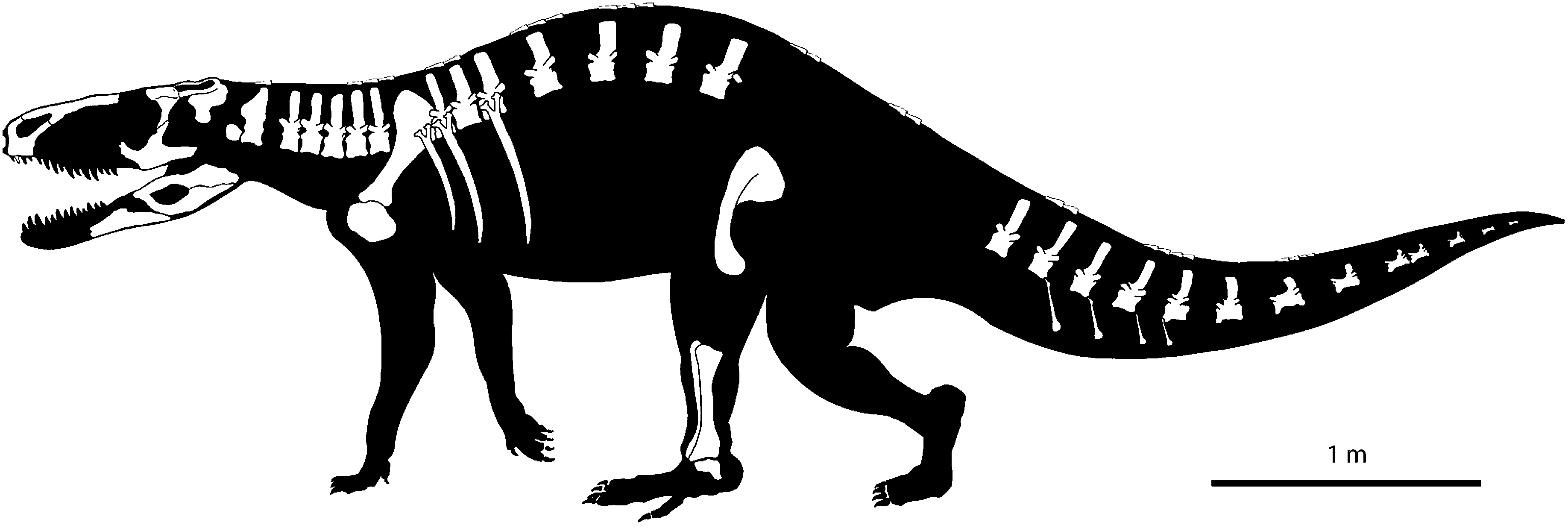
BSPG AS XXV 60–68, 71–100, 105–119, 121. Right premaxilla, right postorbital, left squamosal, left jugal, right pterygoid, right nasal, both splenials, left surangular, prearticular and angular, odontoid(?), axis, cervical, dorsal and caudal vertebrae, ribs, chevron bones, right scapulacoracoid, left pubis, right tibia, fibula and astragalus, and body osteoderms.
Type locality and horizon
‘Zahnsanga’ close to Santa Maria, Province Rio Grande do Sul, Southern Brazil; Alemoa Member, Santa Maria Formation, Lower Carnian (Late Triassic).
Diagnosis
Rauisuchus tiradentes differs from all rauisuchian archosaurs in the following unique combination of characters (*indicates autapomorphies): main body of premaxilla quadrangular; posterior process of premaxilla longer than main body; *bulging protuberance on the base of the posterior process of the premaxilla; maxilla excluded from external naris; massive, transversely broadened articulation between the anterior margin of the ascending process of the maxilla and the posterior ascending process of the premaxilla and the nasal; anterior part of the nasal notably flexed ventrally; rugose ridge on squamosal and on main body of jugal; dorsolaterally orientated upper temporal fenestra with raised lateral margin; flattened and expanded anterior process of jugal; stepped postorbital/jugal bar; *short and ventrally keeled cervicals without postzygodiapophyseal laminae; *elongated and ventrally grooved caudals with an accessory neural spine and a postspinal lamina; chevron bones with single articular facet; pubis without pubic foot; lunate depression on distal part of fibula; body osteoderms.
DESCRIPTION
The lectotype material of R. tiradentes consists of cranial and postcranial elements ( Fig. 1 View Figure 1 ). The preservation is generally rather poor. Most of the material is completely disarticulated, and in parts broken or distorted. Delicate surface structures and bone sutures are only rarely visible. The pronounced alteration of the size and shape of some elements is a result of replacement by calcite and haematite, giving the respective bones the ‘swollen’ appearance that is typical of fossils from the Santa Maria Formation ( Holz & Schultz, 1998).
SKULL
Premaxilla
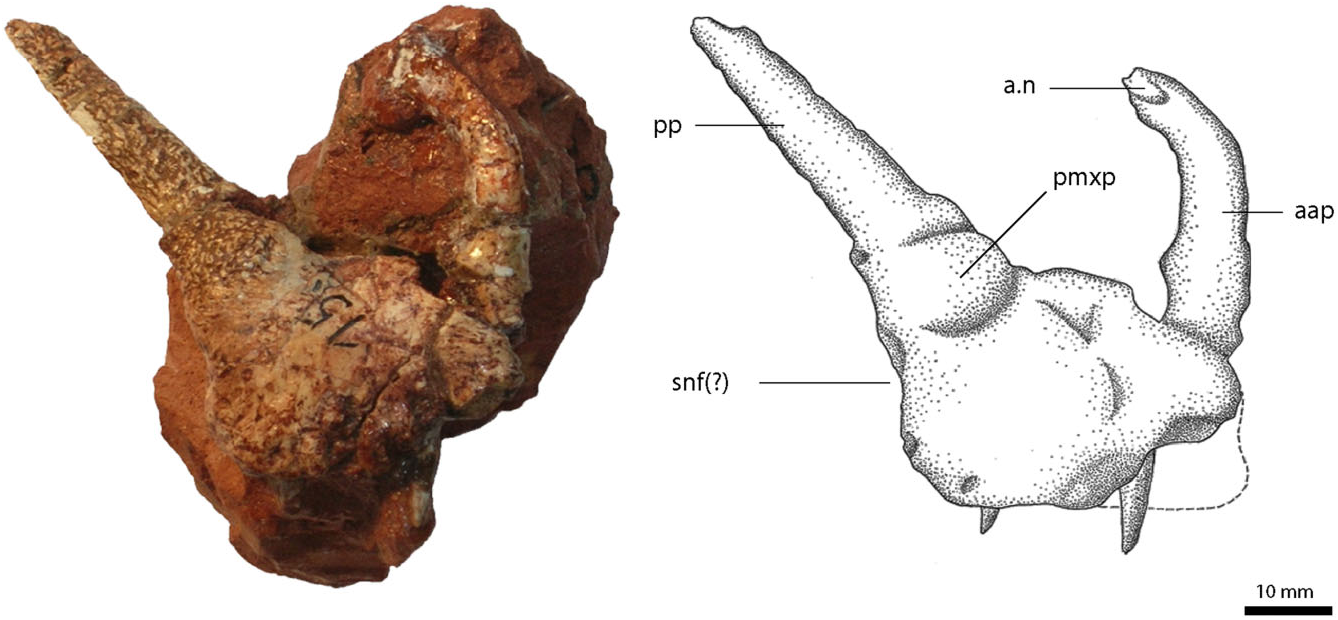
The main body of the right premaxilla ( Fig. 2 View Figure 2 ) is nearly rectangular and only slightly longer than high. Two processes, the anterior ascending process and the posterior process, extend from the main body. Unlike the condition in Batrachotomus kupferzellensis Gower 1999 (SMNS 52970), large nutrient foramina on the premaxillary body are absent, and the narial fossa is present only as a small inflection of the lateral wall below the external nares. On the lateral surface, at the base of the posterior process, the premaxilla has a prominently bulging protuberance, above which the bone surface seems to be more notably rugose than on the premaxillary body. A similar protuberance on the ascending process, as found in Decuriasuchus quartacolonia ( De França et al., 2013), is absent in R. tiradentes .
The anterior ascending process is mostly complete, and forms the anterior and anterodorsal border of the external naris. Its basal portion is more or less vertical and the distal part abruptly flexes posterodorsally at an angle of approximately 60° towards the basal part, which gives the tip of the snout a blunt shape. The process is rather thin, much longer anteroposteriorly than wide transversely, and has a flattened medial surface for the contact with the process of the other premaxilla. The distal tip of the anterior ascending process bears a small triangular facet laterally, for the articulation with the nasal. The process is very thin in this region and its distalmost portion is missing.
The posterior process, forming the ventral and partly the posterior border of the external naris, is at least 1.5 times the length of the premaxillary body, although a few millimetres might be missing distally. It tapers towards its posterior end, where it meets the descending process of the nasal. Its cross section is semicircular with a medially flattened surface, wider than high, and with a flat posteroventral side that overlies the anterior margin of the ascending process of the maxilla. On its dorsomedial surface, there is a small, posterodorsally widening facet that extends over approximately 10 mm anteroventral to the distal end of the process. It obviously represents the contact with the descending process of the nasal, indicating that the maxilla was excluded from the narial margin, unlike the condition in Batrachotomus kupferzellensis ( Gower, 1999) . As noted by von Huene (1942), the lateral side of the process shows a roughened bone surface, whereas the anteromedial narial border is smooth. Because the maxilla of R. tiradentes is missing, the presence or absence of a subnarial fenestra and a possible kinetic premaxilla–maxilla joint, which has been proposed as a characteristic feature of either Rauisuchia (e.g. Parrish, 1993; Long & Murry, 1995) or a more inclusive taxonomic subunit (e.g. Benton & Clark, 1988), cannot be determined with certainty. Furthermore, the distribution of this character within Crurotarsi is not yet completely understood ( Gower, 2000); however, the facet for the maxilla on the posterior process ends slightly above the base of the premaxilla, which would suggest a firm contact between premaxilla and maxilla up to this point. There is only a slight indention below the base of the posterior process on the posterior margin of the premaxilla, which might mark the anterior margin of the subnarial fenestra. In Batrachotomus kupferzellensis (SMNS 52970; Gower, 1999) and Postosuchus kirkpatricki ( Weinbaum, 2002) , however, the subnarial fenestra is nearly entirely formed by a small recess on the anterior margin of the maxilla, without any obvious contribution of the premaxilla. As a result, a subnarial fenestra was not necessarily absent in R. tiradentes . Despite the absence of a maxilla, Nesbitt (2011: character 12) regarded the aforementioned indentation as conclusive for a subnarial foramen in R. tiradentes .
Two teeth, presumably numbers two and four, are preserved. They are comparably short and project only a short distance from the oral margin of the premaxilla, so that these may be replacement teeth that are not yet fully erupted. The anterior and posterior carinae are finely serrated with approximately 25 denticles per 5 mm. The dimensions and positions of the teeth, as well as the length of the premaxilla, indicate that there were no more than four teeth present, as is the case in most rauisuchoids ( Sill, 1974; Bonaparte, 1981; Gower, 1999; Weinbaum, 2002; De França et al., 2013). As preserved, neither the shape nor the number of alveoli can be established with certainty, but a total number of six alveoli, as suggested by von Huene (1942: 203) seems highly unlikely.
Subsequent preparation of the medial side revealed the remains of the median symphyseal facet. The palatal process is broken and not preserved. The area below the base of the palatal process shows two deep, nearly horizontal excavations. Chatterjee (1985) described a similar structure in Postosuchus kirkpatricki and suggested an association with olfactory nerves. Remains of a medial shelf extend over the anterior three-quarters of the bone. At its posterior end, just below the base of the posterior ascending process of the premaxilla, there is a pronounced, posteriorly opening pit of some 5 mm in diameter, which most probably marks the articulation with an anteromedial process of the maxilla, as in Batrachotomus kupferzellensis ( Gower, 1999) . The alveolar border is too poorly preserved to say anything about the possible presence and shape of interdental plates.
The general appearance and outline of the premaxilla, with its blunt snout and long posterior process, re- semble that of Saurosuchus galilei Reig, 1959 ( Alcober, 2000) and Postosuchus kirkpatricki ( Weinbaum, 2002) . These taxa also share the nearly quadrangular morphology of the premaxilla base, which is not seen in other rauisuchian taxa such as Batrachotomus kupferzellensis (SMNS 80260, SMNS 52970; Gower, 1999) that possess a more elongated premaxilla.
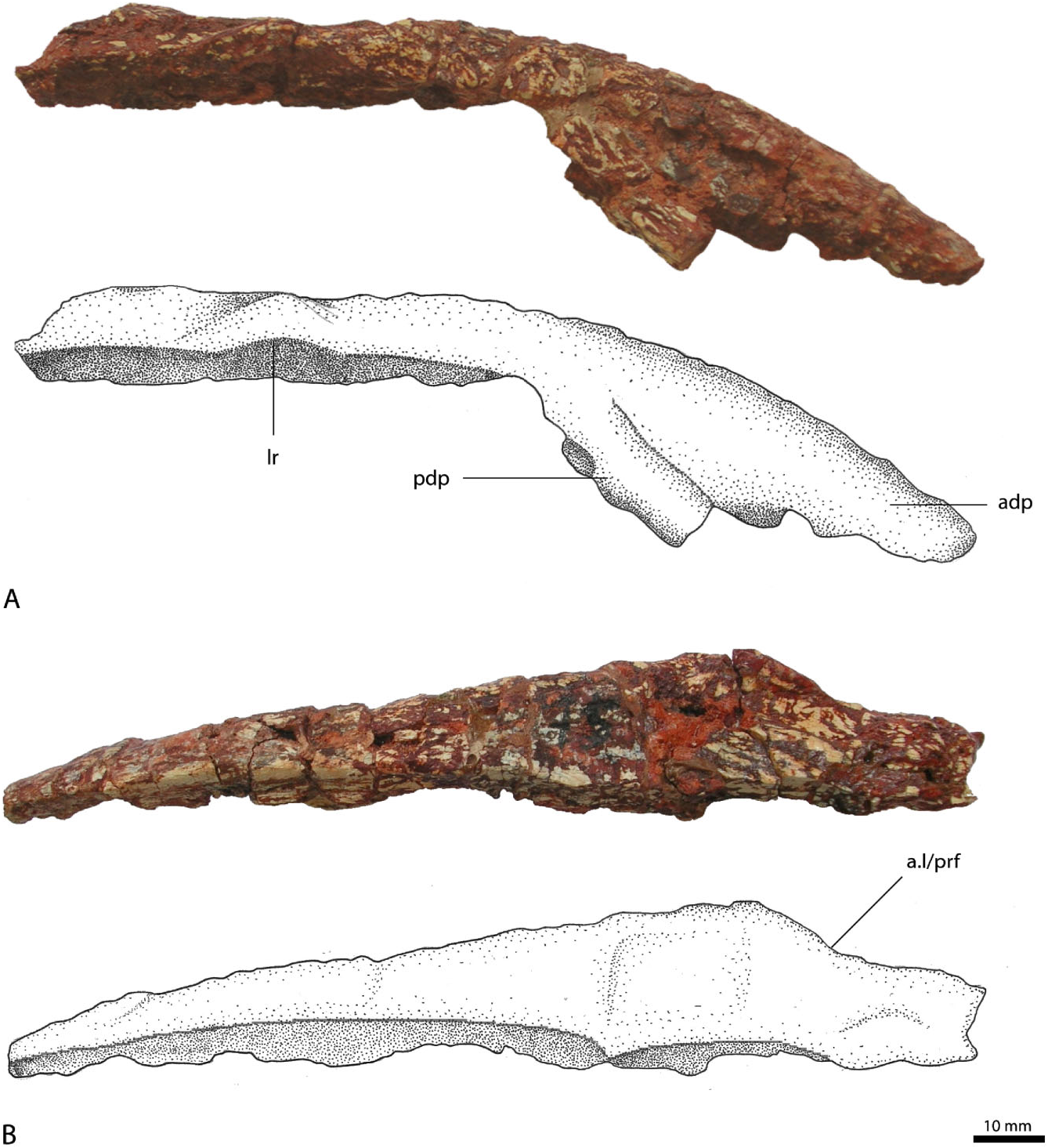
Nasal
The posterior part of the right nasal ( Fig. 3A, B View Figure 3 ) is long and straight, and faces mainly dorsally, whereas its anterior part is strongly curved anteroventrally and splits into two processes. The anterior process is much longer than the posteroventrally located process and is mostly complete, with only the anteriormost part missing. It obviously articulated with the anterior ascending process of the premaxilla, with the comparably massive anterior end laterally overlying the thin posterodorsal end of the ascending process of the premaxilla. The more posteriorly located descending process is incomplete and forms an acute angle with the anterior process, being almost parallel with the anterior process. This process is massive and considerably wider than high at its distal break. It has a flat posteroventral surface, with a slight longitudinal groove medially that is flanked by a small ridge laterally, obviously forming a stout suture with the ascending process of the maxilla. A prominent external narial fossa, located ventrally between the anterior and the descending processes, as seen in Batrachotomus kupferzellensis (SMNS 52970; Gower, 1999), cannot be recognized in R. tiradentes , although a slight depression might have been present.
Because the maxilla of R. tiradentes is not preserved, the exact articulation between the nasal and the maxilla cannot be established; however, the massive shape and outline of the posterior process of the premaxilla and the small facet described above suggest that there was a strong contact between it and the descending process of the nasal. The external naris was thus probably completely framed by the premaxilla and the nasal. This morphology closely resembles the naris of Saurosuchus galilei ( Alcober, 2000) in overall appearance, being considerably longer than high and tapering posteriorly. In Saurosuchus galilei the maxilla only contacts the lateroventral margin of the nasal, and the same condition can be inferred for R. tiradentes .
The posterior end of the nasal is almost complete; only the posteriormost part with the suture to the frontal is missing. The dorsal surface of the nasal is wide and flattened, and has only a weakly developed lateral ridge, in contrast to the very pronounced rugose ridges in Batrachotomus kupferzellensis (SMNS 52970; Gower, 1999) and Postosuchus ( Chatterjee, 1985; Peyer et al., 2008). The dorsal surface gradually widens from the anterior end posteriorly up to the suture with the lacrimal and prefrontal, which, in dorsal view, is marked as a lateral constriction of the nasal ( Fig. 3B View Figure 3 ). Approximately beginning at this point, the medial side of the nasal is slightly arched dorsally, although this might be exaggerated by preservation. The main body of the nasal is very massive and forms a plane butt joint with the other nasal medially. The bone only becomes slightly thinner towards the posterior end.
The lateral surface of the nasal slopes medioventrally some 30 mm posterior to the base of the descending process, forming the facet for the lateroventral articulation with the lacrimal. As in Batrachotomus kupferzellensis ( Gower, 1999: figs 6A, 7), the anteriormost part of this suture is marked as a dorsally expanded, semicircular facet, at which the laterodorsal margin of the bone is slightly raised. The lateral constriction of the nasal in dorsal view is placed immediately posterior to this expanded facet.
The nasal of R. tiradentes differs from that of most other rauisuchian taxa in the marked ventral flexure of its anterior half. In Batrachotomus kupferzellensis ( Gower, 1999) , Saurosuchus galilei ( Alcober, 2000) , Postosuchus kirkpatricki ( Weinbaum, 2002) , and Polonosuchus silesiacus ( Sulej, 2005) (compare also Brusatte et al., 2009) the bone is straight over most or all of its length in lateral view. Given the morphology of the nasal and the premaxilla, the snout of R. tiradentes must have been long and narrow, making up at least 30 to 40% of the total length of the skull. Postorbital
A slender, elongate bone that von Huene (1942: 199) considered problematic was described by him as the left postfrontal, with a small part of the postorbital attached. However, the line interpreted as a suture by von Huene (1942: pl. 24, fig. 2) rather seems to be an overprepared break, judged by the continuing surface structure across this line on the posterior part of the medial side. The element is unusual in its morphology, but we interpret it as the right postorbital ( Fig. 4A– C View Figure 4 ). The only alternative interpretation would be that as a partial left lacrimal, but the lack of a lacrimal foramen and the lack of an expanded anterior process, as well as the tapering rather than expanded ventral end argue against such identification.
The bone consists of a prominent ventral process and an indicated, although incomplete posterior process. The anterior process is completely missing, but the striated anterior surface of the dorsal part of the anterior margin is reminiscent of a suture rather than a break, so that this process might have been very short, as in Arizonasaurus babbitti Nesbitt 2005 , and shows a massive suture with the postfrontal [assuming that the postfrontal had a similar position as in Batrachotomus kupferzellensis ( Gower, 1999) , rather than the more medial position seen in some other noncrocodylomorph pseudosuchians ( Weinbaum, 2002; Sulej, 2005)]. Just medial to this sutural surface, a smaller anterior process along the anterolateral margin of the supratemporal fenestra, as present in Batrachotomus kupferzellensis ( Gower, 1999) , might be broken off. The dorsal surface of the bone is flat and slightly depressed, with a raised medial shelf or ridge along the margin of the supratemporal fenestra. This margin is developed as a slightly dorsoventrally expanding, narrow, medially-facing surface anteriorly, which gently twists into a dorsomedially-facing facet posteriorly. As preserved, the dorsal surface is approximately trapezoidal in shape and narrows from the medial side posteriorly. Anteriorly, its medial part considerably overhangs the base of the ventral process. Because the anterior and the posterior processes are missing, no conclusions can be drawn regarding the articulations with the frontal anteriorly and the squamosal posteriorly.
The ventral process tapers towards its end and is directed slightly anteriorly, although it is not kinked or stepped, to meet the ascending process of the jugal. The ventral process forms the posterior border of the orbit and the anterior margin of the infratemporal fenestra. It is relatively straight and resembles the postorbital of Saurosuchus galilei ( Alcober, 2000) , which also possesses a straight ventral process as opposed to the anteriorly kinked condition of the ventral process seen in Batrachotomus kupferzellensis ( Gower, 1999) and Postosuchus kirkpatricki ( Chatterjee, 1985) . The lateral surface of the ventral process bears a thick, slightly rugose, triangular area dorsally. It becomes gradually lower ventrally and fades into the smooth surface of the transversely thin ventral part of the process at about half its length. The posterior border of this thickened area is marked by a concave, posterolaterally-facing depression along the margin of the lower temporal fenestra. It curves posterodorsally along the gradual transition, ventral to the posterior process of the bone. The lateral side of the ventral half of the ventral process is smooth and slightly convex anteroposteriorly. In medial view, the posteri- or margin of the ventral process is deflected medially, resulting in the formation of a longitudinal depression along the process, which becomes more prominent dorsally.
The ventral end of the process abruptly narrows posteriorly some 15 mm above the ventral tip, which might be missing a few millimetres. This part of the postorbital obviously overlapped the postorbital facet of the jugal anterolaterally.
Although the postorbital of R. tiradentes (if identified correctly) is highly unusual in its morphology, it shows some similarities with the postorbitals of Arizonasaurus babbitti ( Nesbitt, 2005) and Batrachotomus kupferzellensis (SMNS 80260; Gower, 1999). In addition, the element is distinctly different from the lacrimal of Postosuchus kirkpatricki ( Chatterjee, 1985) , Batrachotomus kupferzellensis (SMNS 80260; Gower, 1999), and Teratosaurus silesiacus ( Sulej, 2005) in lacking an anterior process and a pronounced anterior–posterior expansion.
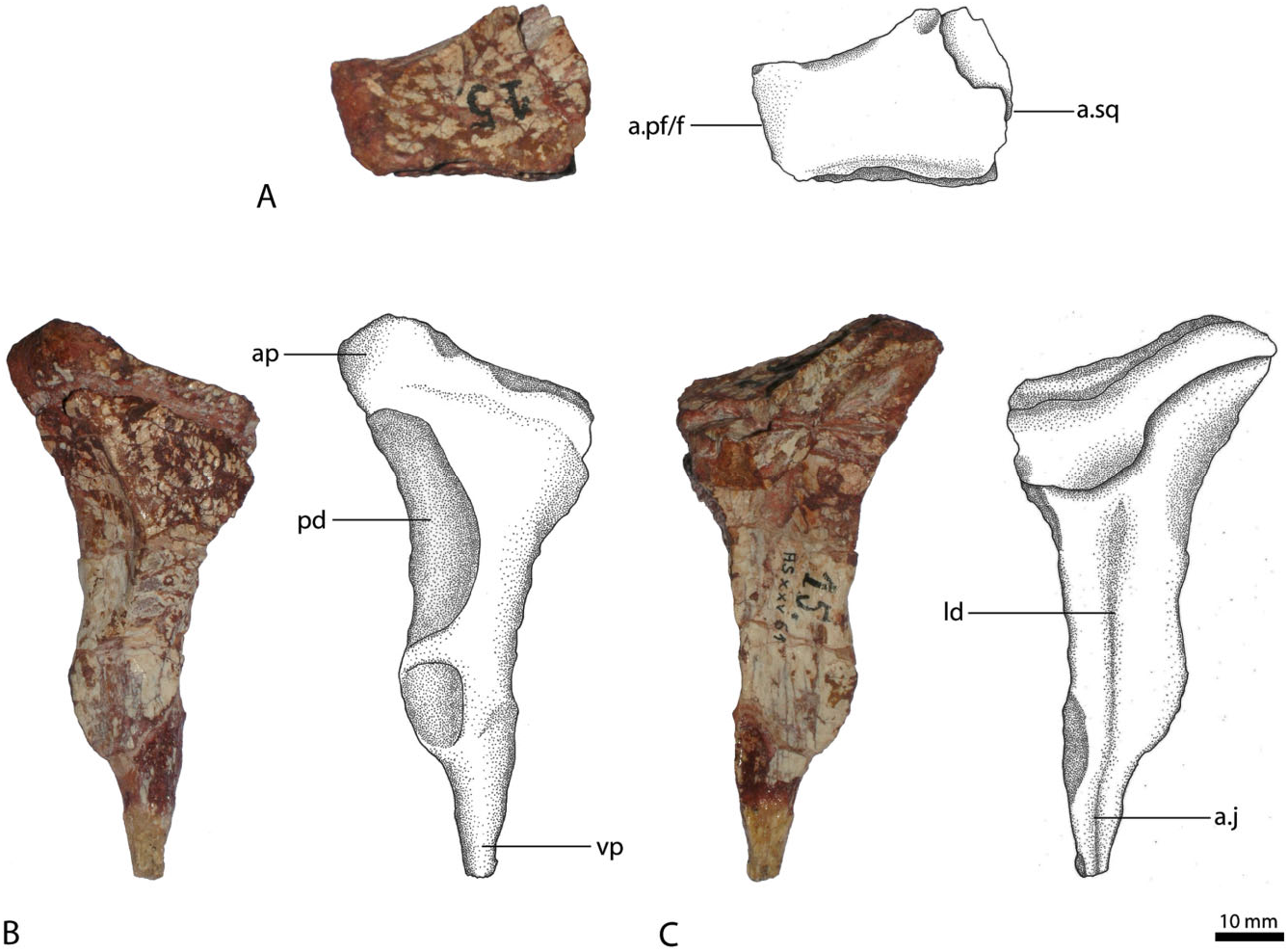
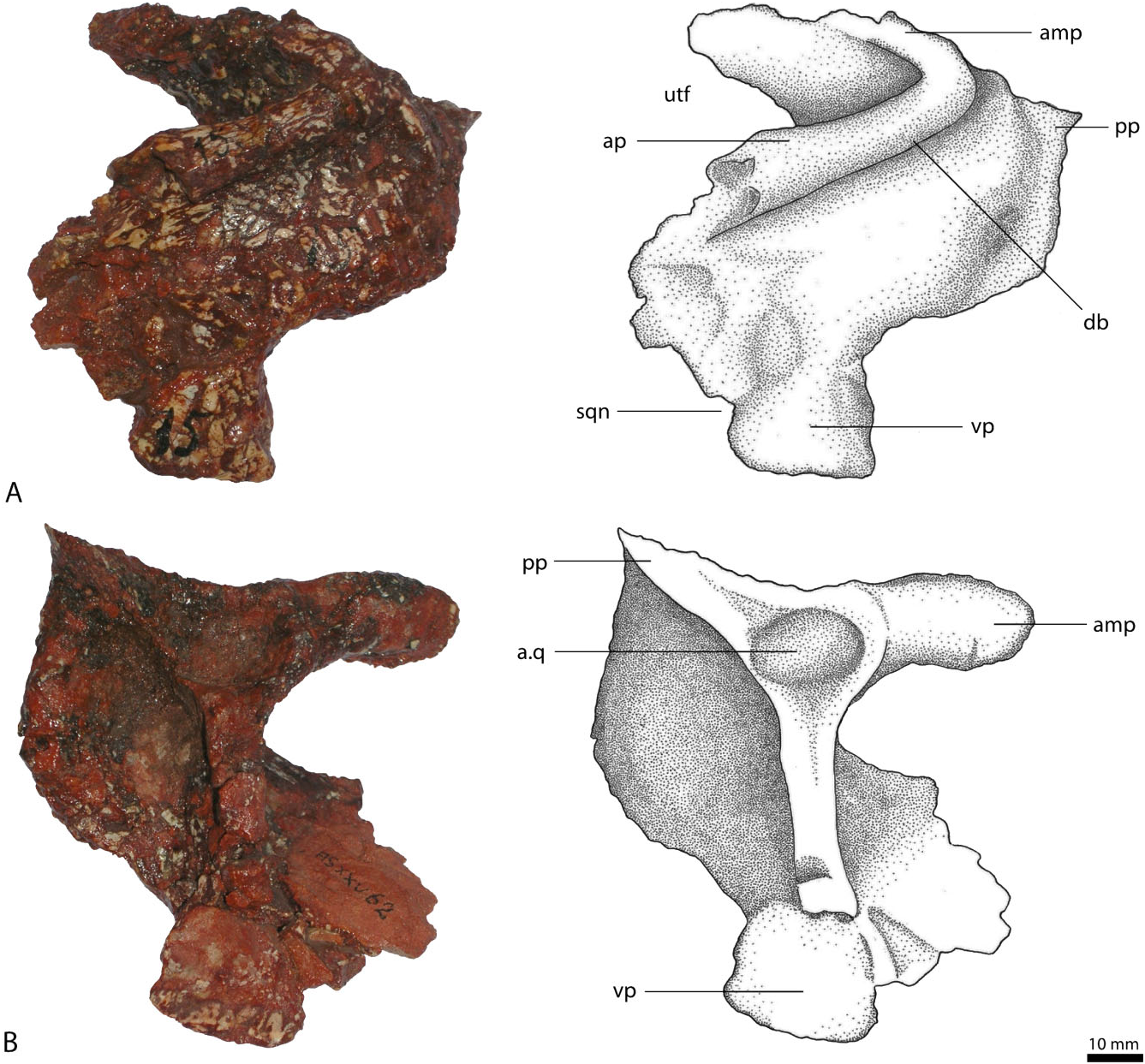
Squamosal
The preserved squamosal ( Fig. 5A, B View Figure 5 ) is from the left rather than the right side (contra von Huene, 1942: 198). It has four processes directed approximately anteriorly, anteromedially, ventrally, and posteriorly. The anterior process projects slightly ventrally and laterally, and only its base is preserved. The anterior end with the socket for the reception of the postorbital is missing. The dorsolateral margin of the anterior process is strongly thickened and forms a raised, rugose ridge along the lateral margin of the supratemporal fenestra, overhanging the dorsolateral part of the squamosal. Gower (1999) described a similar structure in Batrachotomus kupferzellensis as a ‘dorsolateral brow’. Below this ridge, the body of the squamosal forms a ventrolaterally extended shelf, although this might be slightly exaggerated by deformation of the generally poorly preserved bone.
As preserved, the anteromedial process turns slightly in the medial direction and is as long and prominently developed as the anterior process, the medial end of which, and thus the suture with the parietal, is missing. The poorly preserved posterior side forms a vertical to slightly dorsoposteriorly inclined, flat surface for the articulation with the lateral process of the parietal and the paroccipital process of the exoccipital/ opisthotic. The form of articulation with the parietal is unknown. Together with the anterior process, the anteromedial process forms a major part of the posterolateral border of the upper temporal fenestra. The two processes are set at an acute angle of approximately 45° to one another. Because of the positions of these two processes, the upper temporal fenestra is located further laterally than in other rauisuchians, with the exception of Saurosuchus galilei ( Alcober, 2000) . The ventral process is comparably thin transversely, flattened and slightly convex in lateral view, and correspondingly concave in medial view. The complete outline of the process, particularly its anterodorsal margin, is unclear. Nesbitt (2011: fig. 11) reconstruct- ed this process as anteriorly enlarged, contacting the ascending process of the jugal, partitioning the infratemporal fenestra. Although this condition is present in Postosuchus kirkpatricki ( Chatterjee, 1985) and Polonosuchus silesiacus ( Sulej, 2005) , neither the jugal nor the squamosal of R. tiradentes show any indication for such morphology. In fact, the anterior and ventral processes of the squamosal are set off by a distinct notch on the posterior border of the infratemporal fenestra. A similar notch is found in Saurosuchus galilei ( Alcober, 2000) , where the ventral process perforates the infratemporal fenestra, but does not bisect it. The same morphology thus seems more likely for R. tiradentes . The posterior process is inconspicuous and bears a small, spike-like projection.
On the ventral surface, the squamosal possesses a deep depression for the reception of the head of the quadrate. It is bordered by a three-pronged structure, formed by the anterior and posterior ends of the anteromedial process and the posterior process. The dorsal part of the quadrate was therefore covered by the lateral overhang of the squamosal. In dorsolateral view the squamosal of R. tiradentes resembles that of Batrachotomus kupferzellensis (SMNS 80260, SMNS 52970) and Postosuchus kirkpatricki ( Weinbaum, 2002) , in particular in the morphology and presence of the dorsolateral brow.
The squamosal of R. tiradentes is relatively enlarged regarding the other preserved cranial elements. By contrast the squamosal in almost all other known basal pseudosuchians are in comparison often very small and gracile ( Weinbaum, 2002; De França et al., 2013), and make only a minor contribution to the skull roof. By comparison with the examples given by Holz & Schultz (1998) in their taphonomic study on fossils from the Santa Maria Formation, it seems likely that the squamosal is also bloated, which accounts at least in part for the increased size. The heavily deformed bone texture supports this assumption.
Jugal
The left jugal ( Fig. 6A, B View Figure 6 ) is triradiate, presenting three distinct processes: an anterior process contacting the maxilla and/or the lacrimal; a dorsal ascending process joining the postorbital; and a posterior process articulating with the quadratojugal. The anterior process is directed slightly dorsally and is transversely flattened and dorsoventrally expanded. Laterally, it bears a large, but shallow, triangular posteriorly pointed depression, which is separated from the also depressed ventral margin by a thin, anteroposteriorly extending ridge. This structure continues on the main body and forms a low, horizontal ridge that becomes broader and more rounded posteriorly until it fades into the lateral surface of the quadratojugal process. A similar ridge is seen in the jugals of Postosuchus kirkpatricki ( Long & Murry, 1995; Weinbaum, 2002) and Arizonasaurus babbitti ( Nesbitt, 2005) , where it was described as a rugose ridge. In both Postosuchus and Arizonasaurus this rugose ridge continues on the maxilla, which might also have been the case in R. tiradentes , although the ridge becomes notably lower towards its anterior end. The depressed facet ventral to it on the anterior process of the jugal probably represents the facet for the articulation with the maxilla, as is the case in Batrachotomus ( Gower, 1999: fig. 13B). On the medial surface the anterior process of the jugal has a medially thickened dorsal margin and a notable step at about its mid-height. The area between these structures is slightly depressed, forming a subrectangular facet, possibly for the overlap of the ventral end of the lacrimal, but as neither the lacrimal nor the maxilla are preserved, the exact contact between these elements and the jugal is unknown. Gower (1999) described a corresponding, longitudinally striated shelf on the lateral surface of the anterior process of the jugal in Batrachotomus kupferzellensis , which articulates with the maxilla and a lightly striated medial area that articulates with the lateral surface of the ventral tip of the preorbital bar of the lacrimal.
The dorsal process projects slightly posteriorly and tapers towards its distal end, the tip of which is missing. On the upper part of its lateral surface, the dorsal process bears an angular facet for the articulation with the ventral process of the postorbital. This facet occupies the entire breadth of the dorsal end of the process, but narrows towards the anterior margin more ventrally, where it flexes onto the anterior surface of the process and ends some 40 mm below the dorsal tip. Along its anterior margin, the dorsal process shows a characteristic ‘step’, where the facet for the postorbital ends at the anterior margin of the process – a character that has traditionally been employed to define Rauisuchidae (e.g. Benton & Clark, 1988; Juul, 1994). The medial surface of the dorsal process possesses a prominently developed, thin, dorsoventrally extending ridge that arises dorsally from the medial flexure of this facet. This ridge continues to the base of the main body and separates it centrally. On the medial side of the main jugal body, this ridge defines a deep, dorsoventrally elongate, but transversally narrow, anteromedially facing depression. The ridge and thus the border of this depression flexes anteriorly some 20 mm above the ventral margin of the main body of the jugal. A small dorsoventral rib extends ventrally from the ventral margin of the depression and defines the anterior rim of a small, anteroposteriorly concave area on the ventral part of the medial side of the jugal body, below the dorsal process. This concave area probably represents part of the articulation for the ectopterygoid, as is the case in Batrachotomus ( Gower, 1999) and Postosuchus ( Weinbaum, 2002) . The ventral margin of the main body is strongly convex ventrally, so that both the anterior and posterior processes are directed somewhat dorsally. Together, the anterior and dorsal processes form a large part of the posteroventral border of the orbit.
The posterior process is curved and upturned and, compared with the rest of the jugal, is more robust and rounded. Its posterior end is missing. It forms the lower margin of the infratemporal fenestra. Laterally, the posterior process is convex and smooth. The rugose ridge on the main body of the jugal does not continue on the posterior process, but merges with the latter process. Medially the posterior process is slightly flattened, but maintains its circular cross section.
Compared with other basal pseudosuchians, the jugal of R. tiradentes differs notably from that of Batrachotomus kupferzellensis . In R. tiradentes , the posterior process is tapering and curved, whereas it is massive and straight in Batrachotomus kupferzellensis (SMNS 52970 and SMNS 80260). Batrachotomus kupferzellensis also lacks the distinct rugose ridge along the lower lateral side of the main body of the jugal (SMNS 52970 and SMNS 80260). This structure is also absent in Saurosuchus galilei ( Sill, 1974; Alcober, 2000), but is present in Postosuchus kirkpatricki ( Weinbaum, 2002) and Arizonasaurus babbitti ( Nesbitt, 2005) . As in R. tiradentes , this rugose ridge is restricted to the main body and parts of the anterior process of the jugal in these taxa.
Pterygoid
The element described as a right prefrontal by von Huene (1942: 201) is in fact a right pterygoid ( Fig. 6C, D View Figure 6 ), which is somewhat distorted and poorly preserved. The triradiate bone consists of an anterior palatal ramus, the ventrolaterally directed ectopterygoid wing, and the posteriorly directed quadrate wing (which is largely missing). The anterior palatal ramus is elongated. It tapers anteriorly, but the anteriormost part is missing. In ventral view, the anterior portion is narrow transversely, but widens towards its base posteriorly. The medial edge is angled sharply in the dorsal direction and forms a high vertical medial crest, along which both pterygoids were originally joined. This crest also bears a depression running parallel with the anterior palatal ramus, which von Huene (1942: 201) erroneously interpreted as forming the dorsal margin of the orbit. The medial side of the crest is flattened anteriorly, but bears a longitudinal depression in its dorsal part posteriorly. The ventral side of the main pterygoid body and the base of the palatal ramus is slightly inclined dorsomedially. It bears a large, shallow depression posteriorly, which becomes narrower and medially sharply defined on the base of the palatal ramus, but fades more anteriorly.
At the posterior end the pterygoid splits into a ventrolaterally orientated ramus joining the ectopterygoid and a posterior quadrate ramus, neither of which are completely preserved, although the ectopterygoid wing is only missing minor portions of its posterolateral extremity and anteromedial margin. The ectopterygoid wing forms a triangular flange and is as thin and flattened as the anterior palatal ramus. von Huene (1942: 201) considered this lateral ramus to represent a lateral process of the prefrontal, contacting the lacrimal. The posterior part of the ectopterygoid wing is dorsoventrally thickened, although the posterior margin forms a sharp edge. Anterolateral to the thickened part, a depression on the dorsal side of the pterygoid marks the facet for the overlap by the ectopterygoid. In ventral view, the flat ventral surface of the ectopterygoid wing is separated from the main pterygoid body by a notable step.
The basipterygoid articulation expands slightly posterior from the point where the ectopterygoid wing meets the main pterygoid body. Here, the pterygoid is most massive and has a short, but stout longitudinal ventral ridge. The articulation for the basipterygoid process of the basisphenoid is developed as a narrow, poste- riorly opening groove that is semicircular in outline, lateroventrally orientated, and opens dorsally. Thus, assuming that the basipterygoid process was direct- ed lateroventrally, not or only insignificantly anteriorly, the orientation of this facet indicates that, in the articulated skull, the main body and palatal process of the pterygoid sloped considerably anterodorsally, as is the case in most archosaurs. In front of the basipterygoid articulation facet, the median crest of the main pterygoid body is drawn out dorsally into a small, triangular dorsal process.
The quadrate wing of the pterygoid starts at the lateroventral end of the basipterygoid articulation. Its base is dorsoventrally low and ventrally continuous, with the ridge ventral to the basipterygoid articulation. The wing expanded posterodorsolaterally, but is mostly missing.
MANDIBLE
The left surangular, prearticular, and articular are preserved in their original articulation, although the articular seems to be considerably deformed and incomplete. In addition, both splenials are preserved in isolation.
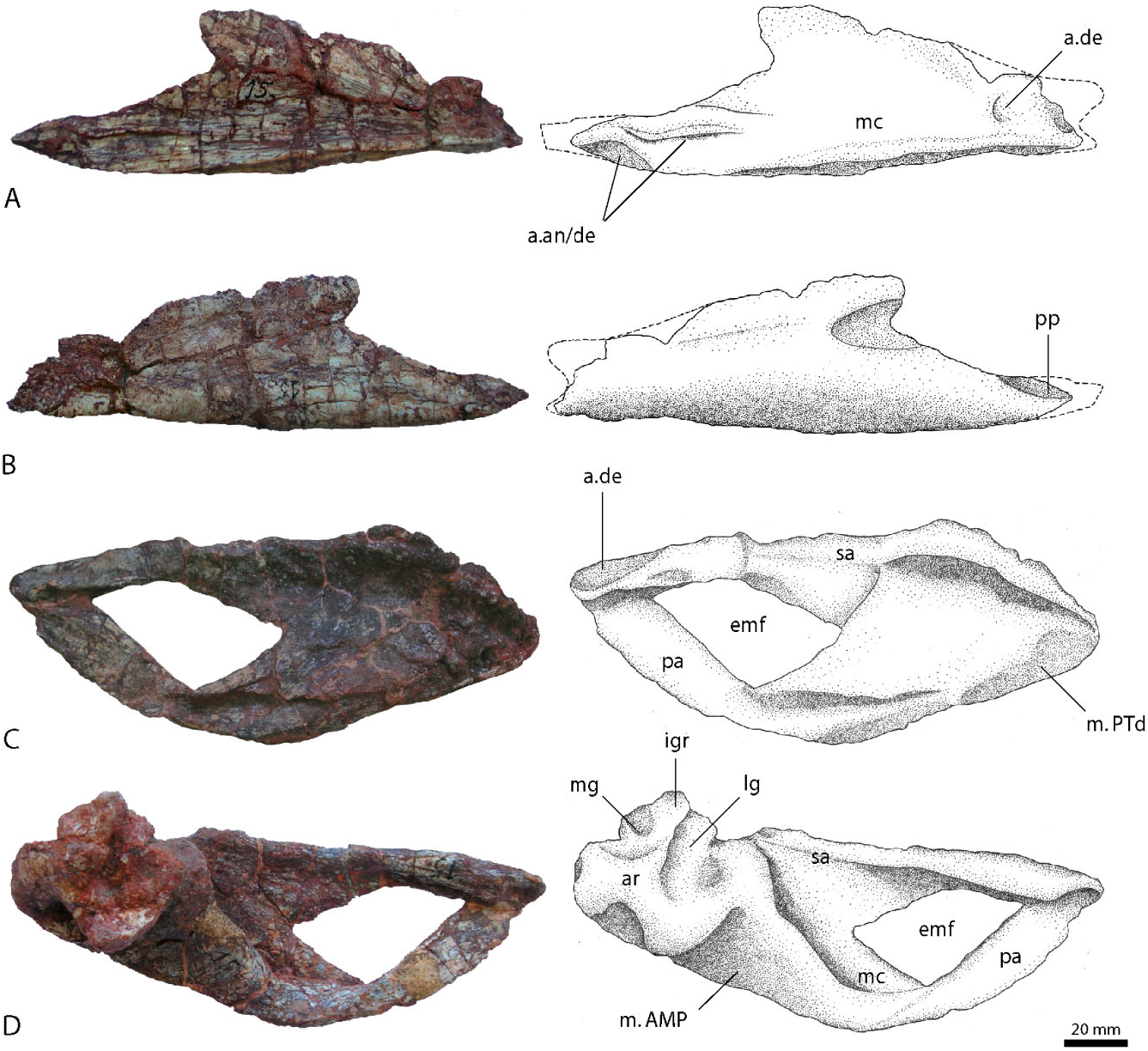
Splenial
The splenials ( Fig. 7A, B View Figure 7 ) are mostly complete, the left element more so than the right, and complement each other in the missing portions. They are elongated and form thin, laminar sheets of bone. Posteriorly the splenial is bifurcated, with a small, spike-like dorsal posterior process (missing in the left splenial) and a much longer ventral posterior process. The ventral margin is thickened and its ventrolateral surface is turned upwards, forming an overlapping ridge, which envelops the ventral border of the dentary and forms the ventral and medial wall of the Meckelian canal. In the right splenial this overlapping ridge is slightly flattened and noticeably grooved lateroventrally, possibly for the articulation with the dentary and the angular. The ridge runs all the way along the posteri- or process in both elements, but becomes gradually lower towards the end. Anteriorly, the ridge becomes a ventral shelf. A second depression, probably also for the reception of the dentary, is located near the dorsal edge on the lateral surface. An articular facet for the coronoid cannot be identified on either side. Unlike the condition in Postosuchus kirkpatricki ( Chatterjee, 1985) and Prestosuchus chiniquensis (BSPG AS XXV 1), the splenials of R. tiradentes lack any evidence for an anterior mylohyoid foramen; however, the left splenial has a distinct notch in the anterior end, which is also seen in the splenial of Prestosuchus chiniquensis (BSPG AS XXV 1), and may be equivalent to the foramen for the mandibular ramus of the trigeminal nerve.
Compared with other taxa, the splenials of R. tiradentes are elongated and more similar to those of Batrachotomus kupferzellensis (SMNS 80260; Gower, 1999) than those of Postosuchus kirkpatricki ( Chatterjee, 1985) , with their triangular shape. The left splenial resembles that of Prestosuchus chiniquensis (BSPG AS XXV 1) in general shape as well as in its dimensions, although this bone is even more elongate and has a suture with the other splenial anteroventrally in the latter taxon.
Surangular
The surangular ( Fig. 7C, D View Figure 7 ) is an elongate element with a triangular or wedge-shaped cross section above the obviously large external mandibular fenestra and a vertical plate in its posterior part. The dorsal part is thickened and forms a prominent, medially and laterally overhanging shelf, especially in the posterior part in front of the articular. Dorsally, this shelf is flattened and bears a broad depression, although this might be amplified by the preservation. In its anterior extreme, the surangular becomes thin and sheet-like, with a thin dorsal margin. The tip has a V-shaped notch on the anterolateral surface for the reception of the posteri- or end of the dentary. Towards the anteroventral margin, which forms the upper border of the lateral mandibular fenestra, the posteroventral plate is mediolaterally compressed and becomes gradually thinner. The posteroventral plate has a well-developed, sharply pointing, triangular anterior process ventrally, so that it forms the posterior border and the posterior part of the ventral border of the mandibular fenestra. The medial surface of the surangular is largely featureless. A surangular foramen as described for Batrachotomus kupferzellensis ( Gower, 1999) and Postosuchus kirkpatricki ( Chatterjee, 1985; Weinbaum, 2002, 2013) cannot be found in R. tiradentes , although this might result from the poor preservation. Posteriorly, the surangular expands transversely towards the anterior margin of the jaw articulation. Unfortunately, no suture between the surangular and articular is visible, so to what extent the surangular participated in the jaw articulation cannot be determined.
The broad and flattened dorsal surface of the surangular most probably served as an insertion for the musculus adductor mandibulae externus superficialis (m. AMES) laterally and the musculus adductor mandibulae externus medialis (m. AMEM) medially, indicating a similar myology as in Alligator mississippiensis ( Busbey, 1989) . The comparably large muscle attachment site suggests the presence of welldeveloped jaw adductor muscles in R. tiradentes .
Prearticular
The prearticular ( Fig. 7C, D View Figure 7 ) is strongly curved and has a medially convex cross section similar to that of Batrachotomus kupferzellensis (SMNS 52970, SMNS 80260). The anterior part of the element is turned dorsally and has a thin, spatula-like appearance, the dorsal end of which reaches the level of the ventral margin of the anterior process of the surangular. Posteriorly it becomes considerably broader and expanded. The ventral border is thickened. The medial wall is low and contacts the articular posterodorsally, whereas the lateral wall continues dorsally to join the surangular. Between the medial and the lateral wall runs a deep groove, probably housing Meckel’s cartilage, as inferred for Batrachotomus kupferzellensis ( Gower, 1999) .
Articular
The articular ( Fig. 7C, D View Figure 7 ) is for the most part poorly preserved and considerably deformed, as von Huene (1942: 204) already noted. It is closely attached to the surangular and both elements have been tightly amalgamated during diagenesis, so that a suture between them is no longer visible. Thus the extent of the ar- ticular in the mediolateral direction seems to be very broad, although this is amplified by the deformation.
Dorsally, the articular bears a large facet for the jaw articulation, which starts at the anteromedial margin of the articular and continues posterolaterally. The facet consists of two depressions, which are separated by a small ridge. The lateral depression is anteroposteriorly elongated and narrow, and extends further anteriorly than the medial depression. The latter is anteroposteriorly shorter, but approximately twice as wide transversely. These facets articulated with the paired condyles of the ventral part of the quadrate. Because of the poor preservation, it remains unclear whether the surangular contributed to the mandibular joint, as in Batrachotomus kupferzellensis ( Gower, 1999) or Postosuchus kirkpatricki ( Chatterjee, 1985) . At its posterior margin, the articular facet for the quadrate bears a slightly elevated rim, which continues medially and grades into the dorsal part of the ascending process of the retroarticular region.
The medial surface bears the ascending process and an adjacent posterolateral depression. The ascending process is developed as a dorsally sloping tubercle at the posterior end of the medial articular facet, and is triangular in medial view. The depression is anteroposteriorly elongate and triangular in outline, tapering anterolaterally. There seems to be one or two large foramina present in the anteromedial part of the depression, as in Batrachotomus ( Gower, 1999) , although some uncertainty remains because of the poor preservation. In dorsal view, the region of the ascending process and this depression form a triangular medial tubercle, as is also present in other basal pseudosuchians, although in contrast to Batrachotomus , the tubercle is less well-developed and confluent with the retroarticular process posteriorly. In medial view, there is a pronounced, rounded triangular ventral process below the ascending process, as in Postosuchus ( Chatterjee, 1985; Weinbaum, 2002). A large, rounded depression is present on the anterior part of this process, its posterior rim extending vertically dorsally to the highest point of the ascending process. The ventral margin of the depression is formed by the medially thickend ventral margin of the process. The area lateral to the dorsal depression on the medial tubercle is poorly preserved, but seems to have been somewhat concave. This area bears a depression for the attachment of the musculus depressor mandibulae in Batrachotomus ( Gower, 1999) .
In ventral view, the articular tapers to a blunt and gently rounded point. This part is offset from the main body of the articular and resembles that of Postosuchus kirkpatricki ( Chatterjee, 1985) . Anteriorly the articular connects to the surangular and prearticular via a diagonally inclined, prominent, and rounded ridge, which forms the posteromedial wall of the mandibular adductor chamber. Posteriorly the articular reduces in size and becomes sagittally constricted.
The strongly developed and inclined ridge on the anterior margin of the articular indicates a firm attachment of the musculus adductor mandibulae posterior (m. AMP) along its length, as well as the insertion of the musculus pterygoideus dorsalis (m. PTd) anteroventrally (compare Busbey, 1989).
Dentition
Apart from the two teeth in the premaxilla, 39 isolated teeth were collected and referred to R. tiradentes by von Huene (1942: 203). They are posteriorly recurved, mediolaterally compressed, and have serrated carinae anteriorly and posteriorly, as it is typical for carnivorous archosaurs. Their size ranges from 7 to 26 mm, with a varying morphology from small, broad, and blunt shapes to long, slender, and pointed forms. These teeth are similar to those preserved in the premaxilla, whereas the remaining teeth – particularly the bigger specimens – might be derived from the maxilla. The serrations on the carinae are very fine, with 20–25 denticles per 5 mm in all teeth. The denticles are chiselshaped and perpendicular to the long axis of the carinae. The mesial carina extends to the base of the crown in many but not all specimens; in some it ends a few millimetres above the crown–root junction. As demonstrated by Batrachotomus kupferzellensis ( Gower, 1999) , such variations in shape and size of the various teeth are not uncommon in rauisuchians. Nevertheless, none of these teeth hold any diagnostic value, so that they cannot be referred to R. tiradentes with certainty. Some might have been derived from scavenging carnivorous archosaurs, especially as many seem to have been shed, as evidenced by well-developed resorption pits at their bases. On the other hand, there are several teeth present that are complete and the preserved elements of R. tiradentes show no obvious signs of scavenging, such as bite or gnaw marks. As the respective sediments were deposited in a floodplain facies (see geological setting), an accumulation of these teeth by water currents can most probably be ruled out, also because a more diverse range of elements would then be expected.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |