
Oligoryzomys brendae Massoia, 1998
|
publication ID |
https://doi.org/10.11646/zootaxa.3641.4.9 |
|
publication LSID |
lsid:zoobank.org:pub:0C0C060E-3D7C-40FA-97A6-CA25D7A5A8EB |
|
DOI |
https://doi.org/10.5281/zenodo.5628813 |
|
persistent identifier |
https://treatment.plazi.org/id/03EA87AC-FFAD-FFE7-69FE-4B84A4BFFD15 |
|
treatment provided by |
Plazi |
|
scientific name |
Oligoryzomys brendae Massoia, 1998 |
| status |
|
Oligoryzomys brendae Massoia, 1998
Type locality: “…Tucumán (localidad típica: Cerro San Javier, Dpto. Tafí Viejo) (aprox. 1000 m. de altura)…” = Argentina, Tucumán, Tafí Viejo, Cerro San Javier (ca. -26.78º, -65.37º).
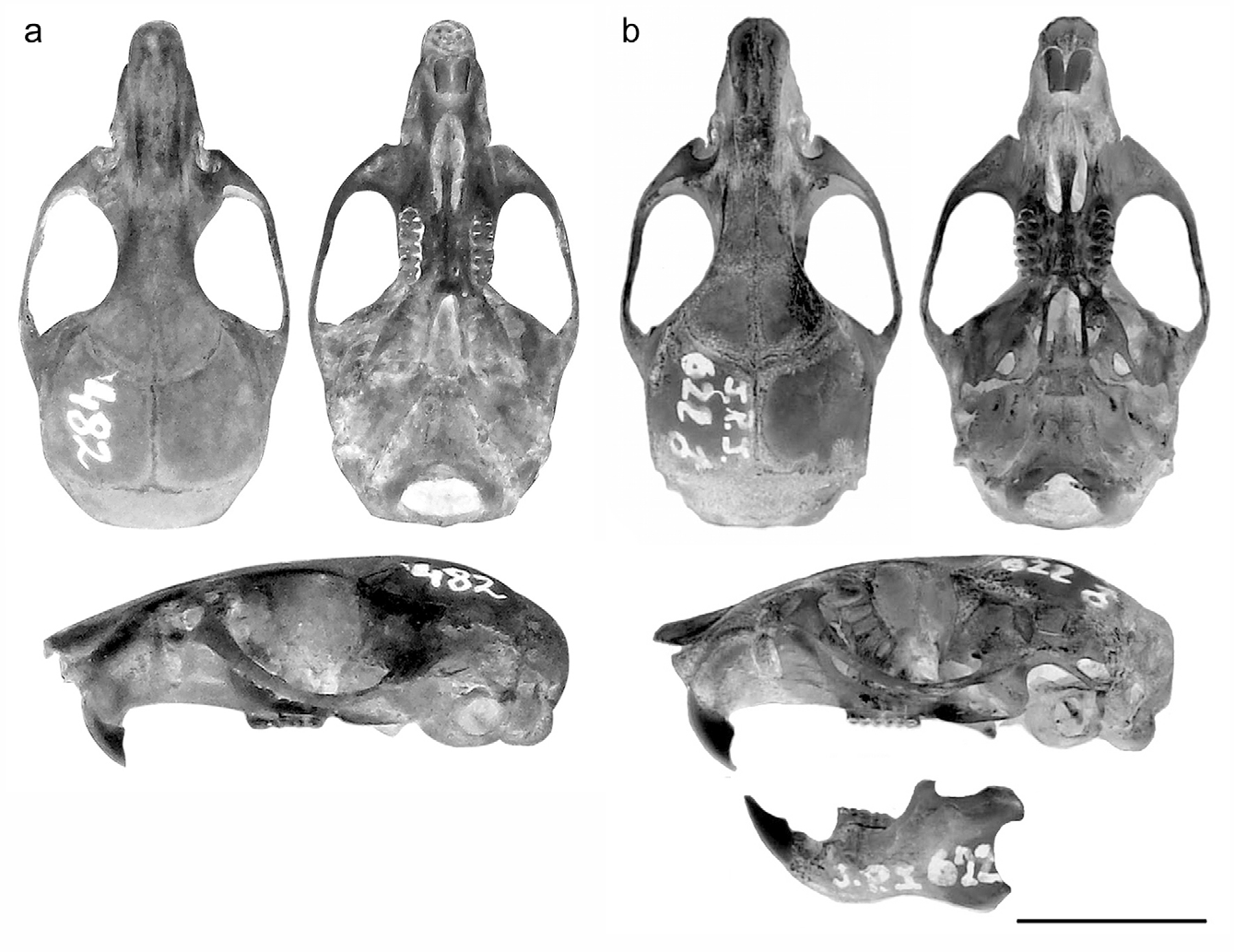
Emended diagnosis. A member of the sigmodontine genus Oligoryzomys characterized by the following combination of characters: size large for the genus (HBL: 90 mm; CIL: 22.9, MTL: 3.9, BB: 12.1; all measurements in average); dorsal coloration orangish brown, with somewhat grizzled appearance; ventral hairs basally grey and whitish to ochraceous at the tip; ears short, rounded, and dark brown; tail longer than head and body and sharply bicolored; skull ( Fig. 3 View FIGURE 3 ) relatively robust, with a short and broad rostrum, well expanded zygomatic arches and a braincase inflated and broad; interorbital constriction narrow and hourglass shaped, with slightly defined supraorbital ridges; incisive foramina relatively long, posteriorly extended to the anterior border of M1; karyotype with 2n = 58, FN = 74.
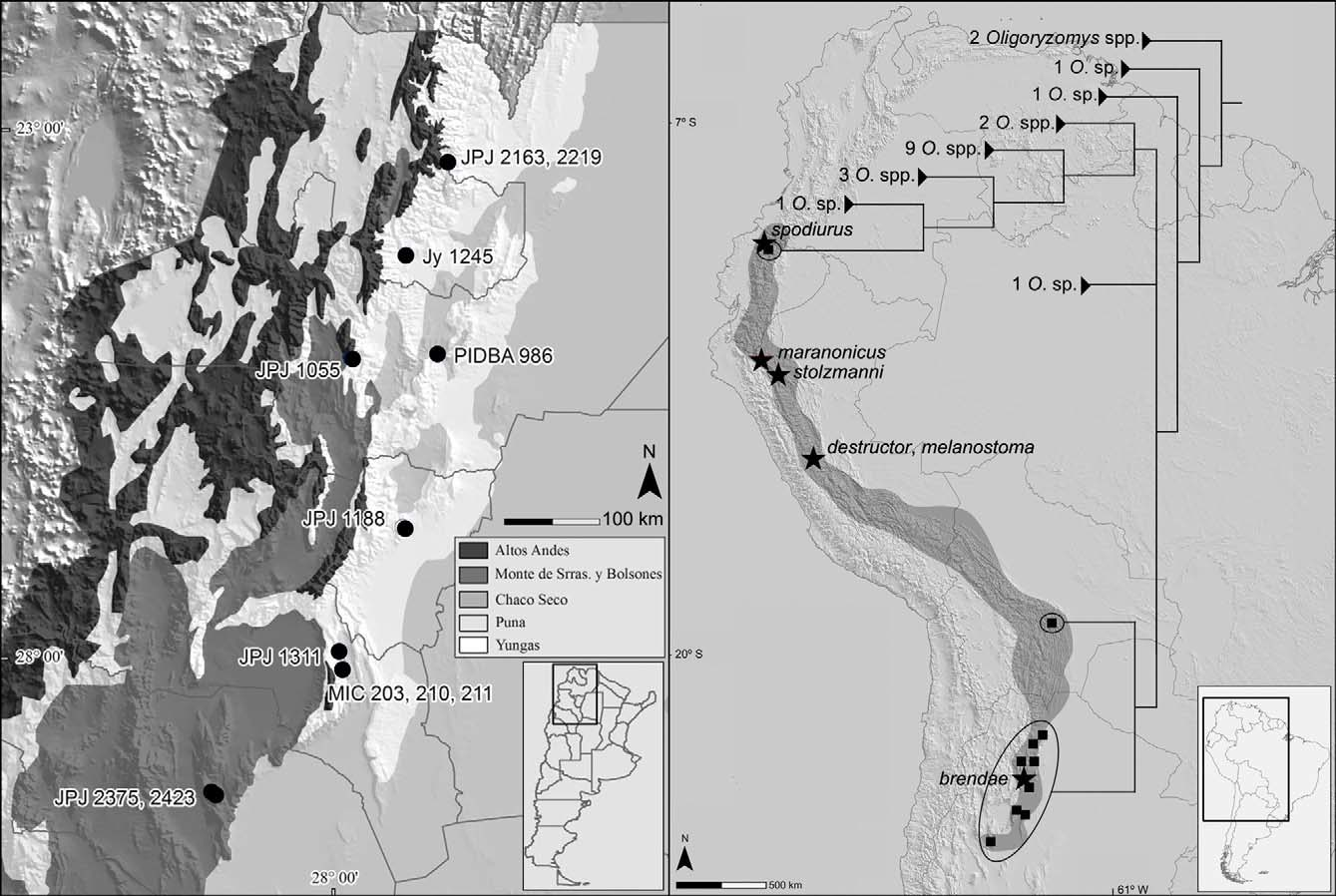
Distribution. Oligoryzomys brendae is mainly found in forested and highland grassland environments of the Yungas between 700 and 2900 m elevation, from northernmost Salta province southward to Catamarca, Argentina. Additionally, we recently recorded the species further south in La Rioja province where it is restricted to isolated humid ravines at the ecotone between the xeric Chaco Seco and Monte desert formations. Jayat et al. (2011b) cited a large species of Oligoryzomys for the Famatina range in La Rioja province; nevertheless, the specimens from Pampa de la Viuda reported here (see Appendix 2) constitute the first unequivocal mention of O. brendae for the province of La Rioja.
Description. Oligoryzomys brendae is a large-sized species within the genus ( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 ; Table 1); its dorsal pelage is soft, and dense; individual hairs have gray bases and ochraceous to reddish tips, giving a general grizzled orangish-brown appearance, slightly darker at the head and midline and orangish towards the cheeks and flanks. Cover hairs are 10 to 12 mm long, being larger in the mid rump region; guard hairs are longer and darker, projecting 3–4 mm beyond the fur in the rump. Ventral hairs are basally grey and whitish to ochraceous at the tip. Ears are short, rounded, and dark brown in coloration; its inner surface is covered by short brown hairs with yellowish tips. Eyes are large and are surrounded by a conspicuous ring of dark brown hairs. Mystacial vibrissae are abundant and moderately long, reaching or slightly surpassing the base of the ear. Interamal and submental vibrissae are short and white. Fore and hindfoot are dorsally covered by short white hairs. Ungueal tuft are whitish and slightly larger than claws. The tail is longer than head and body and sharply bicolored, dark grey dorsally and whitish in the belly.
The skull ( Fig. 3 View FIGURE 3 ) is relatively robust, with a short and broad rostrum, well expanded zygomatic arches and braincase inflated and broad. Nasals are slightly expanded in its distal third. The interorbital constriction is narrow and hourglass shaped, with slightly defined supraorbital ridges. Temporal and mastoid crests are scarcely developed. The zygomatic notches are wide and deep. The dorsal profile of the skull is nearly convex. Zygomatic plate is broad, with its anterior margin nearly straight and the upper root slightly slanted. Long and delicate hamular process separates a well developed postglenoid foramen from a small subsquamosal fenestra. Incisive foramina are relatively long, posteriorly extended to the anterior border of M1. Mesopterygoid fossa is broad, with straight and slightly divergent lateral borders; its anterior margin is well behind the posterior plane defined by the M3. Posterolateral palatal pits are rounded and large, located nearly the anterior border of the mesopterygoid fossa. Parapterygoid fossae are wide and relatively deep, with convex external borders. Auditory bullae are inflated and with relatively broad and long eustachian tubes. The associate foramina of the otic capsules and cephalic arterial patterns do not differ from the typical condition seen in other Oligoryzomys species and fully described by Carleton & Musser (1989). Both specimens lack alisphenoid strut, sphenofrontal foramen and the accompanying squamosalalisphenoid groove.
The mandible is robust and comparatively deep. The masseteric crest is well developed, with their anterior end situated above the mental foramen and extending at the level or slightly beyond the anterior border of m1. The capsular projection is strongly developed and situated below a shallow sigmoid notch. The coronoid process is comparatively broad and very short, extending slightly above the level of the condyle. The angular process is also heavily constructed and does not surpass posteriorly the level of the condyle.
Upper incisors are ungrooved and opistodont, with orange enamel in its anterior surface. Molars are pentalophodont and brachyodont, with the lingual cusps slightly anterior to the labial ones; the anteromedian flexus is deep and defines prominent anterolingual and anterolabial conules; the anteroloph and parastyle are comparatively short and broad; the mesoloph and mesostyle are well developed and the posteroloph is near to metacone. The M2 is nearly as a truncate oval and closely resembled M1 except for the lack of a well developed procingulum. The M3 is small and in both specimens it has a large inner enamel island. Procingulum and anterolabial cingulum on m1 are well developed; anteromedian flexid is expressed only in young specimens. The anterolophid-metastylid and mesolophid-mesostylid are very short and the posterlophid is well developed. In the m2 the procingulum is vestigial, the mesostylid is small and the posterolophid is well developed. The m3 is large, reaching 50% of the m2, with anterolabial cingulum and ectostylid vestigial.
Morphological comparisons. Our examination of the holotype and paratype of O. brendae and large series of Oligoryzomys from northwestern Argentina allow us to state that there are only one large-sized form of Oligoryzomys inhabiting forested and high altitudinal grasslands areas of northwestern Argentina. This species is Oligoryzomys brendae , which differs from the sympatric O. flavescens by its larger size, darker ears, absence of white hairs in chin and throat, broader rostrum, broader zygomatic notches and more expanded zygomatic arches.
Oligoryzomys chacoensis ( Myers & Carleton, 1981) , that is found at lower elevations and Chacoan environments, has conspicuously posteriorly divergent interorbital edges, being intermediate in size between O. brendae and O. flavescens , and having a more strong orangish dorsal fur that contrast with the whitish underside. A small but distinctive tuft of orangish hairs are anterior to the relatively larger ears. Furthermore, hairs on chin and throat are white to the base. Compared with specimens from populations referred to O. destructor , O. brendae has longer ears ( 17–20 mm, vs. 12–14 mm [ Peru], 14–17 [ Ecuador] and 15–17 [ Bolivia]), larger hindfeet ( 24–28 mm, vs. 22.5–24.5 [ Peru], 22–25 [ Ecuador] and 19–25 [ Bolivia]) and a wider braincase (11.5–12.6 mm, vs. 11.4 mm [ Peru], 10.4–11.2 [ Ecuador] and 10.4–11.8 [ Bolivia]; see Osgood, 1914; Hershkovitz, 1940; Olds & Anderson, 1987; Anderson, 1997). Bolivian populations have more grayish venters (cf. Olds & Anderson, 1987), while Peruvian and Ecuatorian forms (including true O. destructor , and the nominal forms stolzmanni , spodiurus and melanostoma ) are more reddish to ochraceous dorsally and brighter colored (Thomas, 1926a, 1926b; Hershkovitz, 1944; Olds & Anderson, 1987). It is worth noting that specimens referred by Hanson et al. (2011) to O. destructor (TEL 1479, 1481; assessed via photographs), and included in our phylogenetic analysis, match the description of the Ecuadorian populations made by Hershkovitz (1944) under Oryzomys ( Oligoryzomys) spodiurus (= O. destructor , see Musser & Carleton, 2005) in having narrow nasals, expanded palatine foramina, long molar toothrows, wide palates and mesopterygoid fossae, narrow zygomatic arches and grayish tails.
Genetic variation. Our sampling of O. brendae was not large enough to constitute a phylogeographic analysis, but our results provide a preliminary view on the geographic structure of the genetic variation of this species. We analyzed 12 sequences gathered at six localities from the Yungas of Catamarca, Tucumán, Salta and Jujuy provinces and two from Monte-Chaco Seco ecotone of La Rioja province, in northwestern Argentina. These haplotypes are quite similar, differing on average by an observed value of only 0.9 %. In addition, despite of the large geographic coverage of the sample covering more than 700 km (ca. six latitudinal degrees), and several environmental discontinuities (e.g., large rivers, xeric habitats) among the analyzed populations, there is a lack of phylogeographic structure in the analyzed sample ( Fig. 5 View FIGURE 5 ). These two facts reinforce the notion that only one species of large-sized Oligoryzomys , O. brendae , inhabits eastern mountain ranges of nortwestern Argentina. The lack of phylogeographic structure of O. brendae resembles the pattern of A. spegazzini (Jayat et al. 2010) , that has a similar distributional pattern, while contrasts with that shown by Necromys lactens (D'Elía et al. 2008) View in CoL over roughly the same distributional range but mainly restricted to the high altitudinal grasslands of the Yungas. The clarification of causes behind these similitude and difference awaits further studies with larger samplings. Notwithstanding, the environmental history of the region ( Iriondo, 1993; Ortiz et al., 2011 and references therein) and the ecological requirements of these species, with N. lactens View in CoL inhabiting open and isolated highland habitats ( Jayat & Pacheco, 2006), and O. brendae (Jayat et al., 2008, as Oligoryzomys sp.) and A. spegazzinii (Jayat et al., 2010) occurring in more continuous forested habitats, can explain, at least in part, the observed phylogeographic differences.
Final considerations. Given the results of the phylogenetic analysis, a re-evaluation of northern nominal taxa, such as Hesperomys melanostoma Tschudi, 1844 , Oryzomys stolzmanni Thomas, 1894 , Oryzomys stolzmanni maranonicus Osgood, 1914 and Oryzomys ( Oligoryzomys) spodiurus Hershkovitz, 1940 is needed to completely clarify the taxonomic scenario within large-bodied Oligoryzomys of the forested foothills of the eastern Andean region. It is also a pending task to determine the northern distributional limit of O. brendae , until now confidently established on northern Salta province, northernmost Argentina. Anderson (1997) referred many specimens coming from several localities of Bolivia (from the northern La Paz department to southern Tarija) to O. destructor , but the correct identity of these specimens remains to be resolved. In fact, given our phylogenetic results, these populations perhaps correspond to an unnamed form of this genus.
Finally, by showing that a distinct species of Oligoryzomys , O. brendae , inhabits the Yungas of northwestern Argentina, this study reinforces the biodiversity value of this eco-region, which counts with two recently discovered endemic species of Sigmodontinae View in CoL , Oxymycterus wayku and Phyllotis anitae ( Jayat et al. 2007, 2008). Our finding emphasizes the need to intensify field work in this biome, which unfortunately is being highly modified by human activities ( Brown et al. 2001).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Oligoryzomys brendae Massoia, 1998
| Teta, Pablo, Jayat, J. Pablo, Ortiz, Pablo E. & D’Elía, Guillermo 2013 |
A. spegazzini (Jayat et al . 2010)
| Jayat et al. 2010 |
A . spegazzinii (Jayat et al ., 2010)
| Jayat et al. 2010 |
Necromys lactens (D'Elía et al . 2008)
| D'Elia et al. 2008 |
Oligoryzomys chacoensis (
| Myers & Carleton 1981 |
Oryzomys ( Oligoryzomys ) spodiurus
| Hershkovitz 1940 |
Oryzomys stolzmanni maranonicus
| Osgood 1914 |
Oryzomys stolzmanni
| Thomas 1894 |
Hesperomys melanostoma
| Tschudi 1844 |