
Tanacetipathes barbadensis ( Brook, 1889 )
|
publication ID |
https://doi.org/10.5281/zenodo.170393 |
|
DOI |
https://doi.org/10.5281/zenodo.5615447 |
|
persistent identifier |
https://treatment.plazi.org/id/03EB87B1-283F-1245-FE89-63E9FBE3FE08 |
|
treatment provided by |
Plazi |
|
scientific name |
Tanacetipathes barbadensis ( Brook, 1889 ) |
| status |
|
Tanacetipathes barbadensis ( Brook, 1889) View in CoL
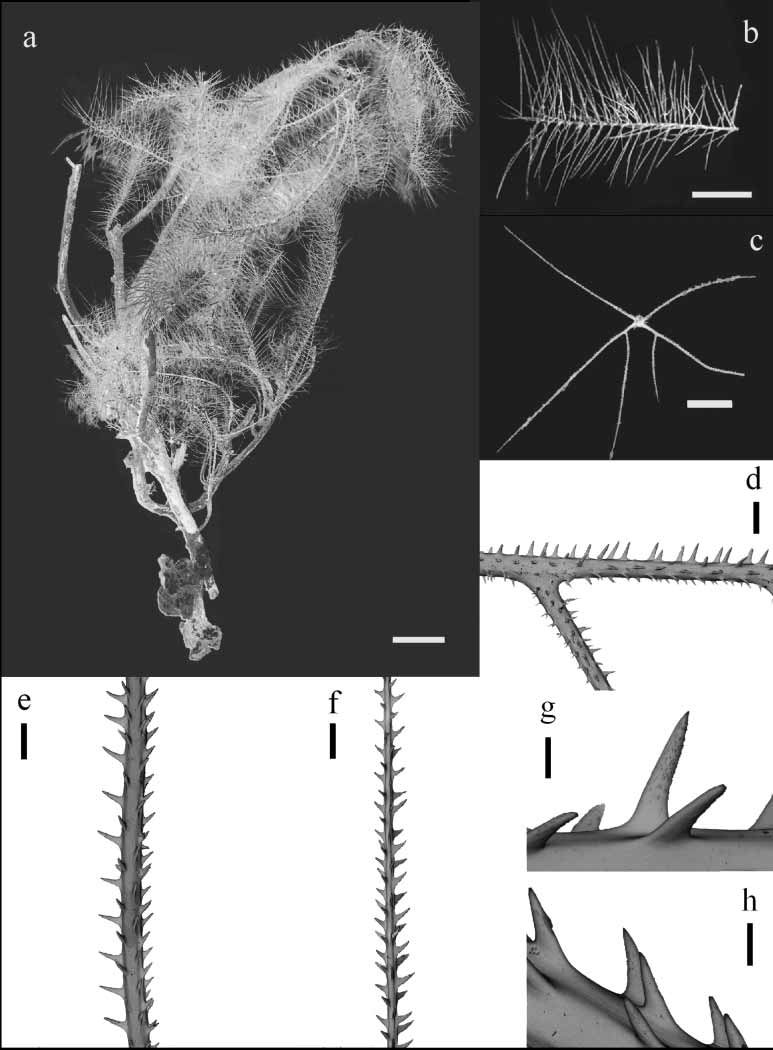
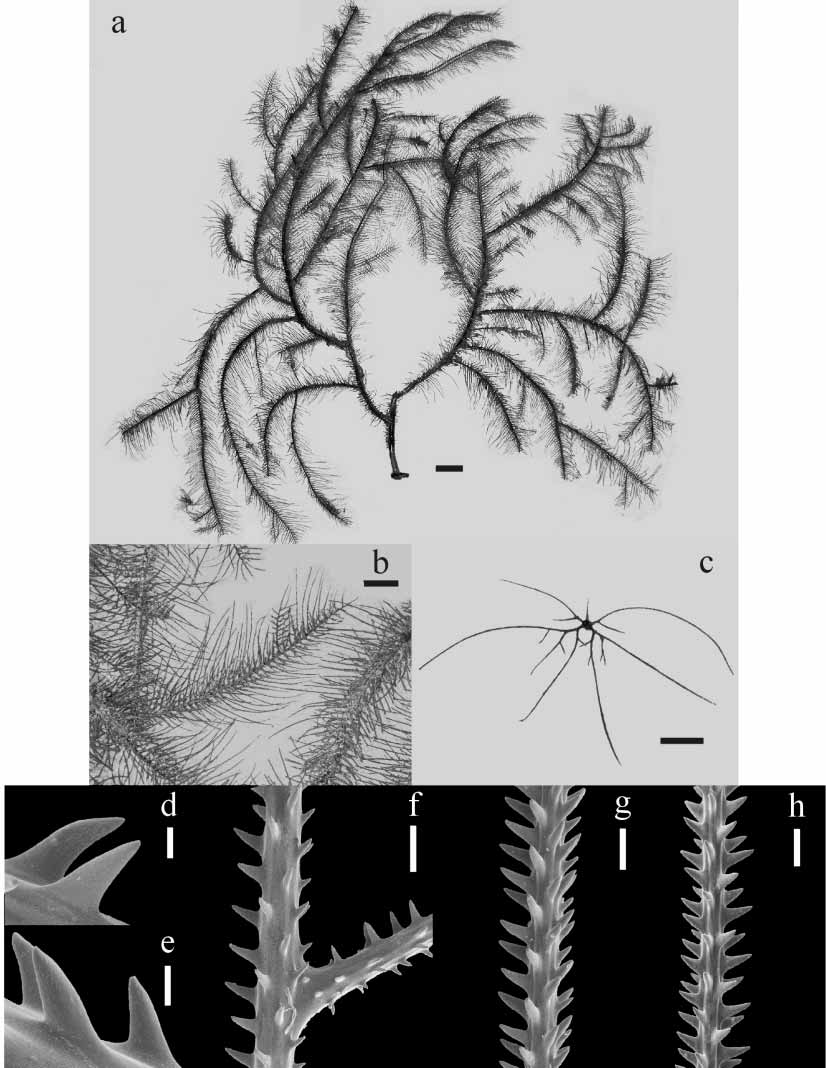
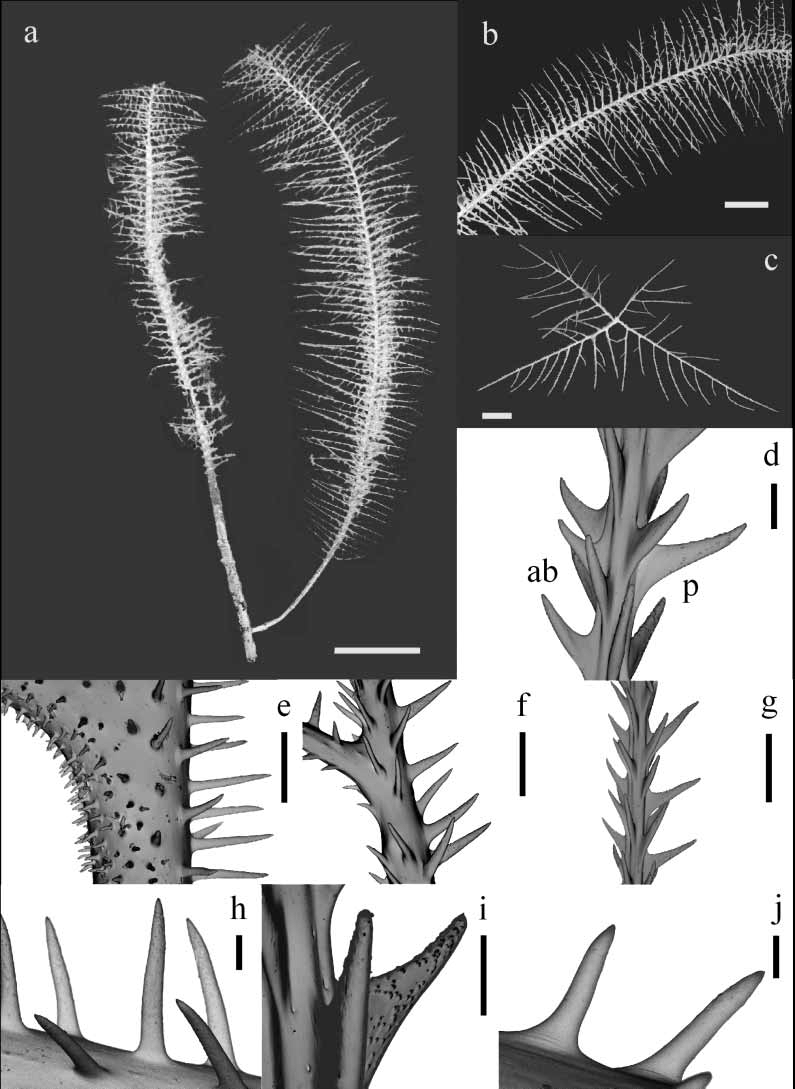
Figures 4–5
Aphanipathes barbadensis Brook, 1889: 128 , Pl. II, Fig. 10 View FIGURE 10 ; Pl. XI, Fig. 4. Antipathes barbadensis: Warner, 1981: 151 –152, Figs. 8 View FIGURE 8 and 9 View FIGURE 9 .
Tanacetipathes barbadensis: Opresko 2001a: 358 View in CoL –361, Figs. 11 View FIGURE 11 b and 12a; 2001b: 349. Antipathes hirta: Echeverría, 2002: 1072 –1075, Figs. 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 , tabs. 2–3 [non Tanacetipathes hirta ( Gray, 1857) View in CoL ].
Material examined. Brazil: Abrolhos Bank: Popa Verde Reefs ( MNRJ 2467, 2548, 2577, 2579, 3071, 4602, 5136: 18 colonies); Timbebas Reefs, 17º30’ S, 039º00’ W, 8m ( MNRJ 2847: 1 colony); Pedra da Lixa, Parcel das Paredes, 17º40’ S, 038º57’ W, about 18m ( MNRJ 4607: 4 colonies); Parcel das Paredes, 17º49’ S, 038º55’ W ( MNRJ 4608: 1 colony). Almost all specimens were collected between depths of 8 and 20 m (pers. obs.).
Diagnosis. Corallum unbranched, sparsely branched, or branched to the 5th order or more; stem and branches pinnulate in a bottlebrush pattern, with up to three orders of pinnules ( Figs. 4 a–c, 5a–c). Primary pinnules in 4–5 (rarely 6) rows, arranged in laterally alternating groups of 2 (when the second anterior pinnule is absent) or 3 pinnules; maximum length of anterior primary pinnules 12–43 mm (average 21.18 ± 7.35 mm), maximum length of posterior primaries, 16–45 mm (average 31.18 ± 8.94 mm). Secondary pinnules usually abpolypar, most often one or two, elongated, very close to the proximal end of the primaries ( Figs. 4 c, 5c); occasionally, primaries with numerous secondaries, up to 20 mm long (maximum length average 14.88 ± 6.44 mm) extending out within the plane delineated by the primaries in each lateral group ( Figs. 4 c, 5c). Primaries with numerous secondaries seem to be an intermediate condition between pinnule and branch, with larger secondary pinnules appearing as primaries of the new young branch. Tertiary pinnules small and rarely present, usually less than 0.5 cm long and occurring primarily near the basal part of the secondaries ( Fig. 4 c). Spines on pinnules simple, elongate, conical, acute, with small ornamentations, and inclined distally ( Figs. 4 d–h, 5d–h). Polypar spines 0.10– 0.30 mm tall, but mostly 0.20–0.25 mm from midpoint of base to apex, 0.03–0.08 mm wide at the base ( Figs. 4 g, 5g); abpolypar spines smaller, slenderer, and more distally inclined than polypar spines, 0.03–0.15 mm tall, 0.01–0.06 mm wide at the base ( Figs. 4 h, 5h). Spines arranged in rows extending along length of axis; 4–6 rows visible from one aspect; on average 4.5–5.5 spines per millimeter in each row; distance between spines in a row 0.15–0.5 times the length of a spine. Polyps 0.5–0.8 mm in transverse diameter, 9–14 per centimeter, in 1 or 2 rows, with small spaces between adjacent ones. (Diagnosis partially based on Brook, 1889, and Warner, 1981; variations observed in Brazilian specimens were also included in this diagnosis).
Remarks. This material agrees with most characteristics observed in Tanacetipathes barbadensis ( Brook, 1889) . There are, however, more secondary pinnules (up to 20, average 8 per primary) in some regions of some colonies. Brook (1889) and Warner (1981) indicated only one or two secondary pinnules, near the base of the primaries. However, three facts lead us to include our material in this species: (1) the condition of only two proximal secondary pinnules per primary is common in our specimens; (2) although having an axial diameter similar to adjacent pinnules, the primaries with a greater number of secondary pinnules seem to be changing into new branches of the colony; and (3) additional secondary pinnules (third and subsequent ones) are shorter than the two proximal ones. In an enlargement of a photo from a colony studied by Warner (1981, Fig. 8 View FIGURE 8 ), it was possible to see the characteristics described above in some primaries. A different interpretation of this character (as an unusual pinnule or as a branch) could explain this incongruity.
There are two groups of specimens from Abrolhos. The first group has one or two, at most three secondaries per primary, always set on the abpolypar side of primaries. Their branches are densely packed and overlapping ( Fig. 5 View FIGURE 5 ). The second group, more numerous, includes specimens that may have more than three secondaries on some primary pinnules ( Fig. 4). These specimens also present sparser and more diverging ramification, where few branches overlap. When several secondaries are present on a primary, one or two most proximal are more elongated, while the others are short. In this case, some secondaries may occur on the polypar side of primaries.
All specimens identified as Antipathes hirta Gray, 1857 , by Echeverría (2002) are among the specimens here studied. We believe his identification was in error, since several characters in his description are in disagreement with his own data. Although Echeverría’s description mentions 4–6 rows of pinnules, his illustrations and, especially, his tabulation indicate 4–5 rows ( Echeverría, 2002: Figs. 6–7 View FIGURE 6 View FIGURE 7 , tab. 2). Also, several aspects of Echeverría’s description of pinnulation are in disagreement with his figures. The secondary pinnules were described as arranged in a single series on the basal half of the abpolypar side of primaries, but his figures ( Echeverría, 2002: Figs. 6–7 View FIGURE 6 View FIGURE 7 ) show several instances where this did not occur. Among 28 pinnule cycles in his figures, only two showed tertiary pinnules and none showed quaternaries (we had access to his original, enlarged plates). This situation is closer to T. barbadensis (rare tertiaries and quaternaries absent) than to T. hirta (see following diagnosis of T. hirta ). Regarding spines, he had to greatly extend the range of the polypar spine length in the diagnosis of T. hirta , originally 0.07–0.13 mm long (see following diagnosis of T. hirta ), in order to accommodate his specimens. This would not be necessary if they were classified as T. barbadensis .
T. barbadensis is very close to T. cavernicola Opresko, 2001 b, from similar depths at Madeira (NE Atlantic). We could not find major differences between these species, except for the maximum length of primary pinnules, and that the latter species form small compact colonies with branches often from near the base. Further studies may show they represent a single species.
These are the first records of T. barbadensis from the South Atlantic; collected in recesses at the base of reefs on the Abrolhos Bank.
Distribution. Atlantic: Barbados ( Brook, 1889); Boca de Navios, NW Trinidad ( Warner, 1981). Brazil: Abrolhos Bank (about 17 ° –18º S—Fig. 1).




| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Tanacetipathes barbadensis ( Brook, 1889 )
| Loiola, Livia L. & Castro, Clovis B. 2005 |
Tanacetipathes barbadensis :
| Echeverria 2002: 1072 |
| Opresko 2001: 358 |
Aphanipathes barbadensis
| Warner 1981: 151 |
| Brook 1889: 128 |