
Biswamoyopterus laoensis, Sanamxay, Daosavanh, Douangboubpha, Bounsavane, Bumrungsri, Sara, Xayavong, Sysouphanh, Xayaphet, Vilakhan, Satasook, Chutamas & Bates, Paul J. J., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3686.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:8597B7A3-060C-4E61-A036-09C1EB351DE8 |
|
DOI |
https://doi.org/10.5281/zenodo.6165378 |
|
persistent identifier |
https://treatment.plazi.org/id/03EB87E4-9D0B-A973-FF08-F8DFBA30A55B |
|
treatment provided by |
Plazi |
|
scientific name |
Biswamoyopterus laoensis |
| status |
sp. nov. |
Biswamoyopterus laoensis View in CoL n. sp.
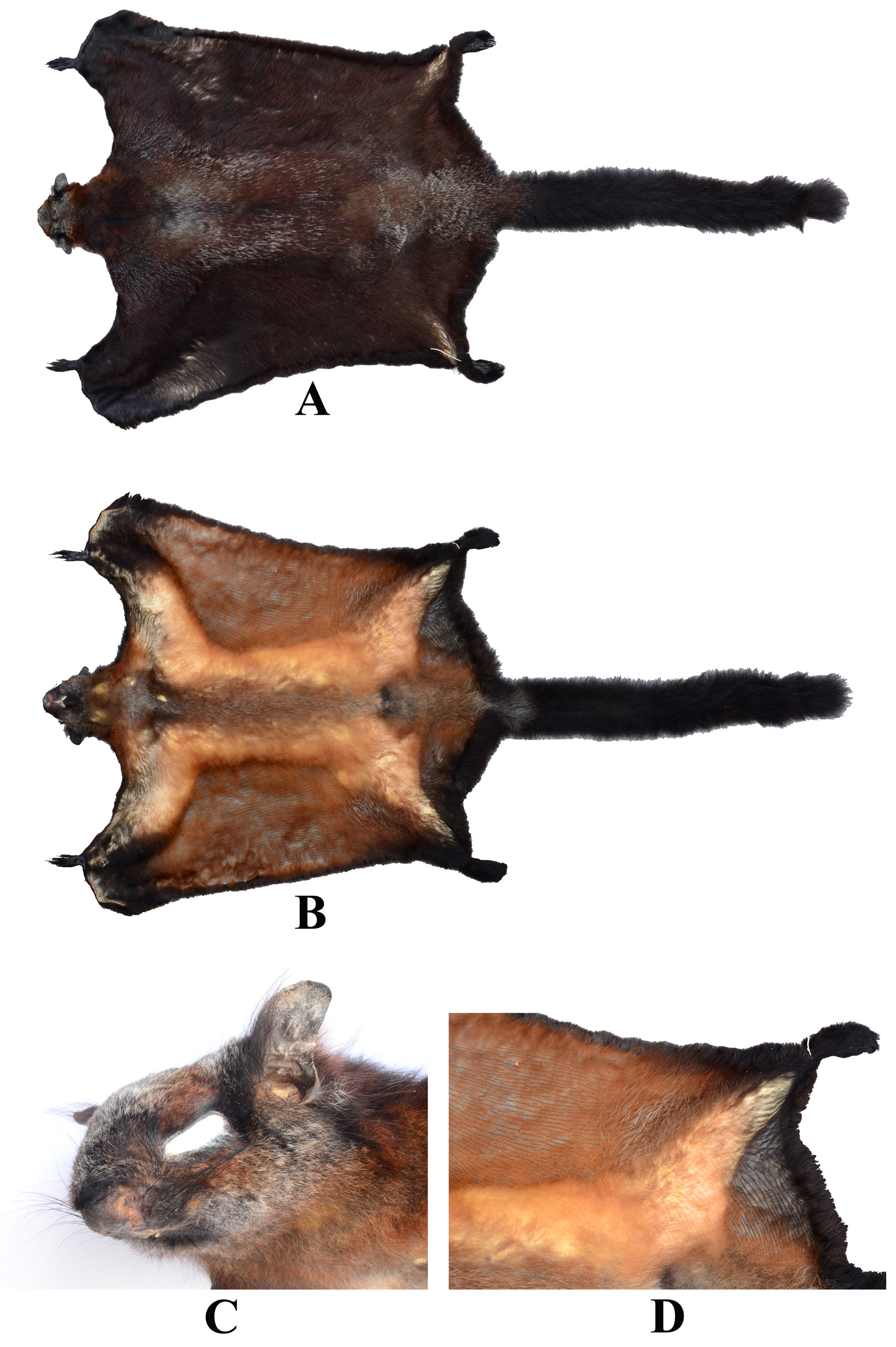
Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 , Table 1.
Type material. Holotype: FES.MM.12.163 (Field No. DS20120925.11), adult female, skull extracted, dry skin, captured by a hunter, collected from the market by Daosavanh Sanamxay, Sysouphanh Xayavong, and Vilakhan Xayaphet on 25 September, 2012.
Type locality. Collected from Thongnami market, Ban (village) Thongnami, Pak Kading District, Bolikhamxai Province, Lao PDR, 18°10.302'N, 104°14.115'E ( Fig. 1 View FIGURE 1 ). Its actual place of origin is not known. However, most of the ‘bush-meat’ sold in this market is sourced from two NBCAs (National Biodiversity Conservation Areas) namely Nam Kading NBCA, which is about 5 km Northeast of Thongnami and Khammouan Limestone NBCA, which is about 25 km Southeast of the village.
Etymology. The name is derived from Lao PDR, the country where the type specimen was collected and ensis (Latin for ‘belonging to’). Its proposed English name ‘Laotian giant flying squirrel’.
Diagnosis. This is a large flying squirrel (a head and body length of 455 mm, a tail length of 620 mm, and a body mass of 1800 grams). The tail is cylindrical, not distichous and is entirely black from its tip to the outer border of the well developed interfemoral membrane, with the exception of a small patch of grey hairs adjacent to the membrane on the under-surface ( Fig. 2 View FIGURE 2 ). The dorsal pelage is an admixture of dark reddish brown and black hairs, variably grizzled with whitish-grey. The ventral pelage is orange intermixed with grey. The upper surface of each patagium is thickly haired, comparable in colour to the upper body but with significantly less grizzling; the underside is sparsely covered in orange hairs and with numerous, clearly defined, discontinuous black lines; these lines are also present on the underside of the interfemoral membrane ( Fig. 2 View FIGURE 2 ). The ears are dark grey/black, virtually naked but with tufts of long black hairs at the base of the anterior and posterior margins; a small tuft of hairs is also present beneath the mid-part of the ear, posterior to the posterior margin of the eye. The skull is large with an occipital nasal length (ONL) of 74.4 mm. The tympanic bullae are large and closely aligned to one another, with a narrow basiocciput ( Fig. 3 View FIGURE 3 ); their internal structure includes numerous (12) septae in a complex honeycomb pattern. The anterior tips of the nasals are virtually straight, not rounded. The upper and lower incisors are yellow on their anterior surface, not pigmented with orange. In the cheekteeth, P3 is relatively well developed and P4 slightly exceeds M 1 in size. Both M1 and M2 have two well developed cusps on the labial side and one large and one smaller cusp on the lingual side; an additional smaller cusp is present on the posterior transverse ridge.
Description: This is a large flying squirrel (Table 1), the upper surface of which is predominantly an admixture of dark reddish brown and black hairs, variably grizzled with whitish-grey. Individual hairs are variable in colour but usually comprise the following components: metallic-ash at the base for up to 20 mm, reddish brown in the mid-part for about 15 mm, and a black tip of about 3 mm. A distinctive whitish-grey band, some 6 mm in length, is present in hairs that reside in areas of the upper surface that are extensively grizzled. These areas of extensive grizzling include the crown of the head, on the upper back, and on the rump extending on to the base of the tail—that part of the tail which is contained within the interfemoral membrane. There is also some grizzling on the upper forearms.
The upper surface of each patagium, which is thickly haired, is comparable in colour to the body but has little or no grizzling. The upper surface of the interfemoral membrane is also well haired, especially adjacent to the tail; except in the region of the tail, it is covered in deep reddish brown hairs. The fore- and hind-feet are covered in black hairs on their upper surface and dark pads on their under surface. The claws form a half circle with very sharp tips.
The ventral pelage is pale orange (peanut butter colour) with white tips to some hairs. On the midline of the body and chest it is dark orange with metallic-ash bases and white tips. The throat is grey and the chin has a dark metallic-ash coloured patch. The anterior margin of the forearms is black, the colour extending to the forefeet, wrist, pro-patagium and styliform cartilage. The undersides of the patagia are more sparsely covered in orange hairs and are marked with numerous, clearly defined, discontinuous black lines ( Fig. 2 View FIGURE 2 ). These lines are stronger and even more clearly defined on the underside of the interfemoral membrane. The lateral margins of the patagia and the interfemoral membrane have a well developed fringe of dark, blackish hairs, above and below. These hairs are metallic-ash at their bases, gradually darkening towards the tip. The interfemoral membrane is relatively large; it is connected to about one third of the basal part of the tail. The tail is cylindrical, not distichous, and is entirely black from its tip to the outer border of the interfemoral membrane, with the exception of a small patch of grey on the underside, adjacent to the membrane. The individual hairs are black at the tip and grey basally.
On the head, the ears are predominantly black and denuded of hairs. Small tufts of long black hairs are present at the base of the anterior and posterior margins of each ear and at the base of the ear. The eyes are ringed with a narrow band of black hairs, which extends posteriorly to the anterior base of the ear and anteriorly on to the nose. There is a patch of reddish brown above each eye. The cheeks are a variable black, deep red and grizzled whitishgrey in colour. The upper surface of the head is strongly grizzled in silver-grey. Three pairs of mammae occur on the distal part of the belly.
TABLE 1. Body Mass (in grams) and external and skull measurements (in mm) of two species of genus Biswamoyopterus , B. laoensis n. sp. from Lao PDR and B. biswasi from India ( Saha, 1981). For definitions, see Methods: M—body mass; HB—head and body length; T—tail length, not including hairs; HF—hindfoot length; E—ear length; ONL—occipitonasal length (= greatest length of skull); CBL—condylobasal length; MB—mastoid breadth; ZB—zygomatic breadth; ZH—zygomatic height; BB—breadth of braincase; BH—braincase height; RB—rostrum breadth; NL—nasal length; MWN—maximum width of nasals bone; IOB—interorbital breadth; POB—postorbital breadth; LIF—length of incisive foramina; LBP—length of bony palate; PPL—postpalatal length; LAB—length of auditory bulla; WAAM—width of auditory bullae across external auditory maeti; IBG—inter bullae gap; P3–M3—upper toothrow length; M2–M2—greatest palatal breadth; M1–M1—width of bony palate at first upper molar; P4–M3—lower toothrow length; ML—mandible length; MH—mandible height.
The skull is large with an occipito-nasal length of 74.4 mm. When viewed in lateral profile, it is relatively flat with a low braincase. The orbital regions are large. In lateral view, the zygomatic arches are low and the jugal bone is not greatly expanded anteriorly; when viewed from above, the zygomata are broad, especially posteriorly ( Fig. 3 View FIGURE 3 ). The rostrum is short and wide and the anterior borders of the nasals are nearly straight, not strongly rounded. The interorbital region is narrower than the postorbital region and the frontal depression is deep and well developed anteriorly; there is a pair of large swellings near the midline at the posterior end of the frontal. The postorbital processes are large but relatively short, with a mean length of 7.9 mm. Temporal ridges are present but do not connect to each other.
The squamosal is in broad contact with the anterior, upper part of each bulla, where it forms a broad shelf roofing the dorsal region of the bulla. The mastoid regions are very small and mastoid process is undeveloped. The tympanic (auditory) bullae are very large, thick and opaque. Within each bulla, the septae form a complex, honeycomb pattern. The anterior palatine foramina are well developed. The anterior border of the pterygoid fossa is posterior to the posterior borders of M3-M3. The pterygoids are large, robust, and curve towards the anterior base of the bullae. The basioccipital is relatively narrow with a prominent ridge in the midline.
The coronoid process of each half mandible is well developed; the condylar process has a large dorsal articular surface; the post-condylar process is absent. The angular process is large and downwardly projecting.
In the dentition, the upper incisors are almost proodont. The anterior surface of each tooth is yellow, not pigmented with orange, and there is a deep groove on the inner margin. The small upper premolar (P3) is relatively well developed and is situated in a notch in the anterior border of the second upper premolar (P4). This latter tooth is large and slightly exceeds M 1 in size. P4 has three well defined cusps on the labial side and one large cusp on the lingual side; there is an additional small cusp present in the mid-part of the posterior transverse ridge. Both the first and second upper molars (M1 and M2) have two well-developed cusps on the labial side and one large and one smaller cusp on the lingual side; they also have an additional smaller cusp on the posterior transverse ridge. The third upper molar (M3) has one cusp on the labial side and a well-developed cusp on the lingual; the posterior part of the tooth is sharply laminated.
In the lower dentition, the incisor is yellow (similar to the upper incisor). The first premolar (P4) is subequal in size to the first molar (M1). It has two cusps on both the labial and lingual sides; the anterior lingual cusp is the largest and there is an indistinct cusplet attached to it posteriorly; there is also a small cusplet on the labial side set between, and slightly internal to, the anterior and posterior cusps. The first and second lower molars (M1 and M2) are essentially similar to P4 but are larger and M2 has a less well developed posterior lingual cusp. The third lower molar (M3) is more elongated and larger than M1 and M2 and without a posterior cusp on the lingual side.
Comparisons. Biswamoyopterus laoensis n. sp. is the second species described for the genus. It is distinguished from all other flying squirrels, with the exception of B. biswasi , by its distinct combination of generic characters.
Superficially, it is most similar in size, colouration and morphology to Petaurista and particularly P. philippensis in Lao PDR. However, externally it differs (inter alia) by having ears denuded of long hairs and the ventral surface of the patagium being marked with numerous, clearly defined, discontinuous black lines. In the skull, it has relatively much larger tympanic bullae and a distinctly narrower basiocciput compared to all species of Petaurista . The internal structure of the bullae is also different. In B. laoensis n. sp., the septae within the bullae form a complex honeycomb pattern whilst those in Petaurista have four or fewer septae. In the dentition, the upper and lower incisors are yellow rather than the typical orange-red found in Petaurista and the cheekteeth are simplified and strongly cuspidate, not wrinkled and sculptured as in Petaurista . However, in order to rule out any possibility that this species had been described before under a name applied incorrectly to Petaurista , the characters of B. laoensis n. sp. were compared with the type descriptions of the sixteen taxa currently referred to P. petaurista and P. philippensis ( Corbet and Hill 1992; Thorington and Hoffmann 2005), which were named from the Indochinese Division (sensu Corbet and Hill 1992, page 4). P. petaurista and P. philippensis were selected as they are essentially similar in size to B. laoensis n. sp.; a third species, P. elegans was omitted since it is significantly smaller. For P. petaurista , descriptions of the following taxa were compared with B. laoensis n. sp.: barroni Kloss from Thailand; candidula Wroughton and taylori Thomas from Myanmar. For P. philippensis , the taxa included: hainana Allen, rufipes Allen and yunanensis Anderson from China; cineraceus Blyth, mergulus Thomas, primrosei Thomas, reguli Thomas, and venningi Thomas from Myanmar; lylei Bonhote and stockleyi Carter from Thailand; and annamensis Thomas, badiatus Thomas and miloni Bourret from Vietnam. In all cases, B. laoensis n. sp. could be distinguished by one or more characters.
The structure of the cheekteeth of Biswamoyopterus also differentiates the genus from Trogopterus Heude , which has a highly complex cusp pattern as well as a greatly enlarged P4; this latter character is shared with the genus Eupetaurus Thomas , the pelage of which is extraordinarily woolly. The genus Pteromyscus Thomas shares with Biswamoyopterus the character of larger tympanic bullae but the cusp pattern of the cheekteeth is once again more complex, P3 is small or sometimes absent, the tail is distichous and not cylindrical, and there is no interfemoral membrane. These latter two characters are shared with the genus Aeretes Allen , which additionally has upper incisors that are orange, cheekteeth that are complex, and tympanic bullae with three septae, not numerous septae in a honeycomb pattern as observed in B. laoensis n. sp. The tails of Iomys Thomas , Hylopetes Thomas , Petinomys Thomas , and Petaurillus Thomas are distinctly distichous and there is no interfemoral membrane. However, the cheekteeth are more cuspidate and share some of the characters observed in Biswamoyopterus . The tympanic bullae of Iomys and Hylopetes in particular are significantly smaller than those of Biswamoyopterus , a character shared with Aeromys Robinson and Kloss , which like the other two genera has a significantly broader basiocciput than that seen in Biswamoyopterus and a much less complex structure of septae within the bullae. However, Aeromys , like Biswamoyopterus , has yellow, not orange incisors.
In comparison to Biswamoyopterus biswasi , the only other species known within the genus, B. laoensis n. sp. is essentially similar in size externally (HB: 455 mm, Tail: 620 mm, and Ear: 52 mm in B. laoensis versus HB: 405, Tail: 605 mm and Ear: 46 mm in B. biswasi ) (Table 1) but differs in most aspects of pelage colour. This is most apparent in a comparison of the ventral pelage, which is illustrated and described in Saha (1981, p. 333 and Plate VI B). According to Saha (1981), the underside of the body of B. biswasi is white with the hairs having pearly grey bases. Each patagium is also white below but washed with a faint orange-rufous; the underside of the interfemoral membrane is white with a band of pale Morocco red near the margin; the margin is grizzled with grey and white, more so near the tail root. This compares to the essentially orange, intermixed with white/greyish hairs found on the ventral surface of B. laoensis n. sp., which also has a very well developed fringe of dark/black hairs on the border of the interfemoral membrane. In addition, the patagia and interfemoral membranes of B. laoensis n. sp. are marked with numerous, black, discontinuous lines, which are not apparent in B. biswasi .
The tail of B. biswasi is cylindrical and on the ventral side is pale grey-white for most of its length, darkening towards a dark brown/black tip. In B. laoensis n. sp., it is also cylindrical but black throughout its length on the ventral side except for a small patch of grey near the border of the interfemoral membrane and orange-grey for that portion of the tail that lies within the interfemoral membrane. On the dorsal surface in B. laoensis n. sp., the tail is also black for its entire length except at its base where it is covered in dark reddish-brown hairs, grizzled with whitish-grey. However in B. biswasi , the tail beyond the interfemoral base is initially pale smoky grey changing to rufous red, then russet and finally to dark brown near the tip; within the interfemoral membrane the tail is grey washed with red.
The upper body of B. biswasi is Morocco red grizzled with white. This is similar to B. laoensis n. sp. but a little paler. The patagia in B. biswasi also appear a little paler than those of B. laoensis n. sp. The crown of the head in both B. biswasi and B. laoensis is characteristically pale but this is more extreme in B. biswasi . Both species have ears denuded of long hairs. However, B. biswasi has a well-developed tuft of pale hairs at the posterior base of each ear; in B. laoensis n. sp. this is black.
In the skull and teeth, the occipitonasal length and zygomatic breadth of 74.4 mm and 47.7 mm respectively in B. laoensis n. sp. are essentially similar to those reported by Saha (1981) for B. biswasi , which were 72.4 mm and 47.5 mm. However, the length of the tympanic bullae and upper cheekteeth of B. laoensis n. sp., 17.3 mm and 16.3 mm respectively, considerably exceed those of B. biswasi , 15.5 mm and 15.5 mm respectively. Both species have a deeply depressed frontal, a relatively short palate, well developed palatal foramina, and tympanic bullae with a honeycomb pattern of septae. The structure of the teeth is also similar.
Habitat. Although the exact habitat of Biswamoyopterus laoensis is not known, the village market at Thongnami, where the holotype was found, is approximately 5 km Southwest of Nam Kading NBCA and it is reported that extensive hunting occurs within at least some of this NBCA ( Evans et al. 2000). The area comprises limestone karst with mostly pristine dry evergreen forest or semi-evergreen forest with an elevation of 160–1,600 m. The village of Thongnami is also approximately 25 km Northwest of Khammouan Limestone NBCA, which is also a karst area with a dry open xerophytic flora on the limestone and small pockets of semi-evergreen forest in the valleys and low lying areas ( Thewlis et al. 1998). It is possible that B. laoensis n. sp. may be present in at least one or possibly both of these NBCAs.
Distribution and Conservation. To date, Biswamoyopterus laoensis is only known from Thongnami in Bolikhamxai Province in central Lao PDR. From the locality of the market and the behaviour of the local hunters, it is probable that the specimen was either collected in Nam Kading or Khammouan Limestone NBCAs ( Fig. 1 View FIGURE 1 ). Further investigations should be carried out in these areas as well as Nakai-Nam Theun and Hin Nam No NBCAs, where limestone karst and evergreen forest are also present, to determine the distribution and abundance of this new species.
Currently, the major threats for the wildlife in Lao PDR are habitat loss and harvesting for food ( Duckworth et al. 1999; Timmins & Duckworth 2008). In squirrels, harvesting is the most serious threat as they rank as the most commonly traded mammalian ‘bush-meat’ in local markets ( Srikosamatara et al. 1992). This is a serious conservation concern for B. laoensis n. sp.
Remarks. The discovery of this new species extends the known range of the genus Biswamoyopterus by some 1,250 km. Biswamoyopterus biswasi was previously collected at an elevation of 350 metres in Deban (26 km east of Miao) and until now was only known from the western slope of the Patkai Range in the catchment area of the Noa Dihing River in Namdapha, India ( Saha 1981). Like B. biswasi , the extent of the geographical range of B. laoensis n. sp. is currently not known. However, it is of particular interest that it was collected from a region of central Lao PDR, which is home to other recently discovered rare and endemic rodent taxa such as the Kha-nyou, Laonastes aenigmamus Jenkins et al. (2005) and the Lao limestone rat, Saxatilomys paulinae Musser et al. (2005) . Furthermore, endemism is not restricted to rodents or even mammals: the area is also home to the endemic Lao langur, Trachypithecus laotum (Thomas 1921) and in birds to the recently discovered bare-faced bulbul, Pycnonotus hualon Woxvold et al. (2009) .
It is strongly recommended that further research of this new species be carried out urgently to help determine its range, population status, and its threats.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.