
Copionodon elysium, Pinna & Burger & Zanata, 2018
|
publication ID |
https://doi.org/ 10.1590/1982-0224-20170146 |
|
publication LSID |
lsid:zoobank.org:pub:84A16FA5-9A0B-45BC-A8A5-19BF11B9B9FD |
|
DOI |
https://doi.org/10.5281/zenodo.3716980 |
|
persistent identifier |
https://treatment.plazi.org/id/2D15386B-101C-43E0-9108-47076966C4CD |
|
taxon LSID |
lsid:zoobank.org:act:2D15386B-101C-43E0-9108-47076966C4CD |
|
treatment provided by |
Carolina |
|
scientific name |
Copionodon elysium |
| status |
sp. nov. |
Copionodon elysium , new species
u r n:l s i d:z o o b a n k.o rg:a c t: 2 D1 5 3 8 6B -1 0 1C - 4 3 E 0-9 1 0 8- 47076966C4CD
( Figs. 1-2 View Fig View Fig ; Tab. 1 View Table 1 )
Holotype. MZUSP 120631 View Materials , 45.0 mm SL, Brazil, State of Bahia, Chapada Diamantina, Município de Palmeiras, Vale do Capão near Morrão outcrop, Riacho Águas Claras , tributary of Rio Mucugezinho , itself tributary of Rio Santo Antonio , Rio Paraguaçu drainage, 12º31’19”S 41º28’52”, M. de Pinna, V. Abrahão, V. Carvalho, A. Zanata, 6 Mar 2017.
Paratypes. All from same locality as holotype. MZUSP 121654 View Materials , 52 View Materials , 12.0- 71.5 mm SL, collected with holotype ; UFBA 8100 , 1 , 34.3 mm SL, R. Burger , 15 Jan 2016 ; UFBA 8163 , 47 (1 c&s), 10.3-46.5 mm SL, A. M. Zanata, R. Burger, P. Camelier, R. Freire , 26 Jun 2016 ; MZUSP 120630 View Materials , 15 View Materials (3 c&s), 21.7-59.7 mm SL and UFBA 8115 , 6 , 32.8 -47.0 mm SL, R. Burger , 11 Apr 2016.
Diagnosis. Distinguished from all other copionodontine catfishes by the large eye without a free orbital rim ( Figs. 1 View Fig , 2 View Fig ; vs. large eyes and free orbital rim in Copionodon spp. or with small eyes but without a free orbital rim in Glaphyropoma spp.). Also, uniquely diagnosed in the subfamily by the second hypobranchial entirely cartilaginous ( Fig. 3 View Fig ; vs. presence of a conical ossification cap in the anterolateral corner of the second hypobranchial in other copionodontines) and the reduced or vestigial adipose fin in adults, resulting in a dorsal profile of the caudal peduncle almost straight ( Fig. 1 View Fig ; vs. caudal peduncle dorsally convex as a result of the well-developed adipose fin). Further distinguished from congeners by the thin upper and lower lips, with lower lip not flattened ventrally, continuous and not split in two halves by medial depression ( Fig. 2b View Fig ; vs. broader lips and presence of median constriction on lower lip), mouth inferior, located close to anterior margin of snout (vs. mouth distinctly ventral, located posterior to anterior margin of snout), and by the posteriorly-positioned dorsal fin (predorsal distance 56.7-61.7 % of SL vs. 46.0-55.0 % of SL). The new species is also distinguished by its color pattern, with an uniformly broad midlateral dark ( Fig. 1 View Fig ; vs. narrow dark band in C. pecten , somewhat broad and asymmetrical dark band in C. lianae , or absence of dark lateral band in remaining copionodontines). Copionodon elysium further differs from C. pecten by the smaller interopercular patch of odontodes, distant from lower lip anteriorly and not reaching opercular aperture ( Fig. 2b View Fig ; vs. interopercular patch of odontodes large, extending from near lower lip anteriorly to beyond pectoral-fin insertion posteriorly) and longer barbels, with maxillary barbel usually reaching pectoral-fin origin and rictal one extending beyond midlength of interopercular patch of odontodes (vs. maxillary barbel reaching posterior ¼ of length of interopercular patch of odontodes and rictal reaching anterior ¼ of length of interopercular patch of odontodes). From the species of Glaphyropoma , the new taxon further differs by a series of characters, including the broad hypobranchial ( Fig. 3 View Fig ; vs. slender), the caudal fin bilobed ( Fig. 1 View Fig ; vs. truncate), the toothed portion on dentary not reaching the coronoid process ( Fig. 4 View Fig ; vs. reaching coronoid process), and the presence of asymmetrical spatulate teeth in jaws ( Fig. 4 View Fig ; vs. all teeth symmetrically spatulate).
Description. Body elongate, trunk roughly round in crosssection, gradually more compressed posterior to dorsal fin. Caudal peduncle compressed, but less strongly than in congeners. Anterior part of body and head ( Fig. 1 View Fig ), from snout to dorsal-fin origin, slightly convex. Posterior region of body, from endpoint of dorsal fin to caudal-fin origin, straight to slightly convex. Ventral profile straight along head and gular regions, nearly parallel to longitudinal axis of body; convex or slightly convex from that point to pelvic-fin origin and nearly straight or slightly concave from pelvic-fin base to caudal-fin base. Dorsal profile of adipose fin nearly straight; fin barely developed, somewhat more visible posterior to vertical through end of anal-fin base. Limit between adipose and caudal fins not welldefined.
Head ( Fig. 2 View Fig ) slightly longer than wide or as long as wide in larger specimens, deep and not as depressed as in C. pecten . Mouth inferior, subterminal, located close to anterior margin of snout. Upper lip narrow in ventral view, not flattened ventrally. Ventral margin of upper lip partly continuous laterally with rictal barbel. Lower lip with about same thickness as upper lip; continuous, not divided in lateral halves by median constriction, only slightly concave mesially.Angle of mouth with fleshy outgrowth just posterior and adpressed to rictal barbel.
Dentary and premaxillary teeth somewhat similar to each other in shape and arrangement. Teeth gradually larger towards symphysis ( Fig. 4 View Fig ). First row of teeth spatulate and darkly-pigmented distally in dentary and premaxilla, with spatulate portion in most teeth asymmetrical ( Fig. 4 View Fig ). Asymmetric structure of teeth evident even in teeth only incipiently spatulate. Spatulate portion of teeth not overlapping in lower jaw ( Fig. 4 View Fig ). Distal border of teeth straight. In both jaws, second row of teeth also slightly expanded distally; third row conical. Teeth on lower jaw distributed over anterior two thirds of ossification and not reaching coronoid process. Outer series of lower jaw with about 6-8 spatulate larger anterior teeth followed by 3-5 conical teeth.
Eye large, conspicuous, without free orbital rim and covered with skin ( Fig. 2a View Fig ). Rim of dark pigment along dorsal margin of eye (see below) creates false impression of free rim under cursory examination. Eye located at half of head length, closer to lateral border of head than to midline in dorsal view. Infraorbital latero-sensory canal complete, extending from sphenotic posteriorly to first infraorbital (lacrimal) anteriorly. Three infraorbital pores, one shortly posterior to vertical through posterior margin of eye, another ventral to ventralmost point of eye, and third approximately at vertical through posterior naris. Anterior naris surrounded by short anterolateral tube, continuous with nasal barbel. Posterior naris located between eyes and somewhat aligned longitudinally with anterior naris, not tubular and partly closed by two thin flaps of skin. Three pairs of flat barbels; barbels wide at base and gradually narrowing distally. Maxillary barbel reaching posterior margin of interopercle, or base of pectoral fin. Rictal barbel located ventral to maxillary, rictal extending beyond interopercular patch of odontodes and almost reaching posterior border of interopercle. Nasal barbel originating on posterolateral region of anterior naris, reaching up to opercular opening when completely and straightly extended. Barbels extension ontogenetically variable, longer in large juveniles and mature specimens. Interopercle with well-developed odontodes visible in ventral aspect of head ( Fig. 2b View Fig ). Interopercular patch of odontodes extending from vertical through posterior border of eye anteriorly and somewhat distant from border of opercular membrane posteriorly. Odontodes arranged in two series, medial series well-defined and lateral series somewhat disorganized; odontodes gradually larger posteriorly in all rows, those on inner row largest. Number of interopercular odontodes increasing with size: 5-6 odontodes in single row in small specimens (around 20.0 mm SL) and up to 21 odontodes distributed in two rows in larger specimens. Opercular region smooth, without opercular odontodes.
Pectoralfinlarge(notaslargeasinothercopionodontines), round in distal profile, its base immediately ventral to posterior tip of interopercle. Pectoral-fin rays i7i*(4), i8(4), i8i(10), i9(8), i9i(2), or i10(1). Pelvic fin with round distal profile, its origin anterior to vertical through dorsal-fin origin. Pelvic-fin rays ii4i(1), i4i(2), i5(2), i5i*(23), or i6(2), first ray (unbranched) shorter than others; when adpressed tip of pelvic-fin rays reaching or almost reaching anal-fin origin. Dorsal fin smaller than anal fin and similar in size to pectoral fin, its dorsal profile round. Dorsal-fin origin closer to base of caudal fin than to tip of snout. Dorsalfin rays ii5i*(4), ii6(3), ii6i(13), ii7(6), iii4i(1), or iii5i(1). Anal fin with base longer than that of dorsal, pectoral, and pelvic fins, its distal profile round. Anal-fin origin located aligned or shortly anterior to end of dorsal-fin base. Analfin rays ii9(3), ii10(1), iii9*(5), iii9i(2), iii10(11), iii10i(1), iv8i(1), iv9(1), or iv9i(2). Caudal fin bilobed, with i6/8i(1), i7/8i*(26), or i8/7i(2) principal rays. Upper lobe slightly longer than lower lobe in some specimens. Dorsal procurrent rays 6(2), or 7(1) and ventral 8(1), or 9(2). Adipose poorly developed or vestigial in adults, lacking clear anterior and posterior limits and extending between vertical at base of posterior anal-fin rays and caudal-fin origin. Adipose fin larger and more well-defined in juveniles, clearly derived ontogenetically from larval fin-fold ( Fig. 5 View Fig ).
Lateral line extending from posterior part of head to hypural plate, composed of interrupted series of short soft-tissue tubules lacking ossifications. Vertebrae 34(2), or 35(1) (exclusive of Weberian complex; ural complex counted as one), first dorsal-fin pterygiophore immediately posterior to neural spine of 12 th (10) or 13 th (9*) vertebra, first anal-fin pterygiophore immediately posterior to neural spine of 14th (1), 15th (9), 16th (9*), or 17th (1) vertebra. Pleural ribs 10 (1), 11 (1) or 12 (2). Branchiostegal rays 7 (3). Dorsal-fin pterygiophores 7(1), 8(1) or 9(1), including anterior rudimentary element when present. Anal-fin pterygionphores 12(3).
Coloration in alcohol. Sides of body with large partly coalescent black blotches (not coalescent in few specimens), forming broad midlateral dark band with approximately constant depth from rear of opercle to base of caudal fin ( Fig. 1 View Fig ). Dorsal and ventral borders of band deeply sinuous, resulting in a zig-zag aspect in most the specimens. Lateral band symmetrical along midlateral line, with dark pigmentation somewhat equally distributed dorsally and ventrally to midalteral line. Additional, dark markings positioned dorsally to midlateral band, forming poorlydefined row on each side of middorsal line, restricted to dorsum anterior to dorsal fin origin. Middorsal line not outlined by dark pigment. Dark blotches below midlateral band small, usually barely fused forming inconspicuous narrow band variable in length, initiating in the rear of pectoral fin in some specimens or posterior to pelvic fin in others. Abdominal region white.
Overall color of head ( Fig. 2a View Fig ) similar to dorsal coloration of body or slightly darker, usually without dark blotches; specimens up to 30.0 mm SL may have small and inconspicuous dark markings. Mid-dorsal region of head, corresponding to central portion of skull roof, usually darker and more uniformly colored than surroundings; head dorsum homogeneously dark by concentration of minute melanophores in specimens up to 22.0 mm SL. Dorsal edge of eye outlined as thin dark line. Contour of laterosensory pores on head dorsum distinctly black. Small teardrop-shaped white area posterior to base of nasal barbel ( Fig. 2a View Fig ). Distal edge of skin flap on posterior naris very dark. Sides of head usually homogeneously tan, except in specimens up to 22.0 mm SL, with concentration of dark melanophores forming inconspicuous sinuous strip close to ventral margins of head and opercle. Ventral surface of head mostly white and with scattered minute melanophores along its contour. Anterior narrow border of lower lip and fleshy outgrowth at angle of mouth sparsely pigmented ( Fig. 2b View Fig ); upper lip heavily darkly-pigmented anteriorly, abruptly lighter posteriorly. Skin on region of interopercular patch of odontodes lacking melanophores. Nasal barbel uniformly dark. Maxillary barbel lighter than nasal barbel; dorsal surface darker than ventral. Rictal barbel with melanophores concentrated along its border and leaving central portion white, forming two lateral dark lines; ventral portion lighter and without dark lines. All fins with melanophores alongside individual rays and segment limits. Pelvic and anal fins somewhat lighter than remaining fins. Adipose fin scattered with melanophores, in pattern continuous to dorsal portion of body.
Coloration ontogenetically variable ( Fig. 5 View Fig ). Specimens up to 17 mm SL strongly dark, nearly uniformly black, but with midlateral stripe faintly visible against dark background ( Fig. 5a, b View Fig ). Somewhat larger juvenile specimens with midlateral dark stripe continuous and very prominent ( Fig. 5 c View Fig ). Larger specimens with midlateral stripe less continuous and gradually masked by additional dark markings dorsal and ventral to it ( Fig. 5d View Fig ), approaching adult pattern ( Figs. 1 View Fig , 5e View Fig ). Very large specimens (not shown in Fig. 5 View Fig ) more uniformly dark than smaller ones, with extra pigment partly masking midlateral stripe and other features.
Coloration in life. General color pattern similar to preserved specimens but uniformily darker, without dark blotches or with barely visible small ones. Mid-dorsal region of head with distinctly golden, roughly rectangular blotch. Lateral band not visible or barely visible on caudal peduncle. Fin-rays tan and intervening membranes yellowtinted. Distal portion of caudal peduncle and base of caudal fin light yellow. Ventral portion of body white.
Geographical distribution. Copionodon elysium is known exclusively from riacho Águas Claras, a stream of approximately 6 km, tributary of rio Mucugezinho, itself a tributary of rio Santo Antonio, upper rio Paraguaçu basin, Chapada Diamantina, Bahia State, Brazil ( Fig. 6 View Fig ).
Conservation status. Copionodon elysium is known exclusively from a stream ca. 6 km long, tributary of the Upper rio Paraguaçu basin, within the Chapada Diamantina Domain. The environment of the species has well oxygenated fast-running water running over rocky bottom. Because no specific threats have been detected, the species can be categorized as Least Concern (LC) according to IUCN criteria (IUCN, 2016), however, we emphasize the apparently narrowly endemic distribution of the species, which makes it (and most remaining copionodontines) dependent on the preservation of the delicate headwaters they inhabit.
Etymology. The specific epithet comes from the Greek elysium, in reference to the mythological Elysean Fields: a place or condition of ideal happiness; a state or place of perfect bliss. This alludes to the locality inhabited by the new species, a scenic pristine place ( Fig. 7 View Fig ) shared with a single other fish species and no fish predators. A noun in apposition.
Ecological notes. The Águas Claras stream is perennial and torrential in the rainy season. Its headwaters are around 1000 m above sea level and it runs on the western slopes of the Sincorá Range, partly within the Chapada Diamantina National Park. The population has a patchy distribution in the stream, with individuals concentrating in certain stretches and entirely absent in others. Copionodon elysium was sampled in slowflowing stretches over solid rock bottom or boulders, with riffles and shallow pools 10-30 centimeters deep and 1-3 meters wide ( Fig. 7 View Fig ), with water transparent and tea-stained, though not to the same darkness as is typical in the Chapada Diamantina. Shallow pools have accumulations of plant debris along the margin. Marginal vegetation is typical “Campo Rupestre”, an herbaceousshrub vegetation type that develops at elevations of about 900 m on sandstones and quartzite. Shrubs of Bonnetia stricta (Nees) Nees, Mart. (Bonnetiaceae) dominate the margins along the stretches sampled. The only other fish species collected with C. elysium was Astyanax sp.
The analysis of stomach contents of three specimens of C. elysium includes allochthonous and autochthonous items, composed mainly of aquatic insects ( Trichoptera : larvae of Hydroptilidae and Hydropsychidae ; Diptera : larvae of Simuliidae and Chironomidae ; adult coleopterans; Plecoptera eggs), filamentous algae, and fragments of adult terrestrial insects ( Hymenoptera : Formicidae ), and numerous sand grains.
Considering the pristine condition of the type locality and its location inside an official preservation area, there are no evident threats to the survival of C. elysium , despite its narrowly endemic distribution and probably small population size.


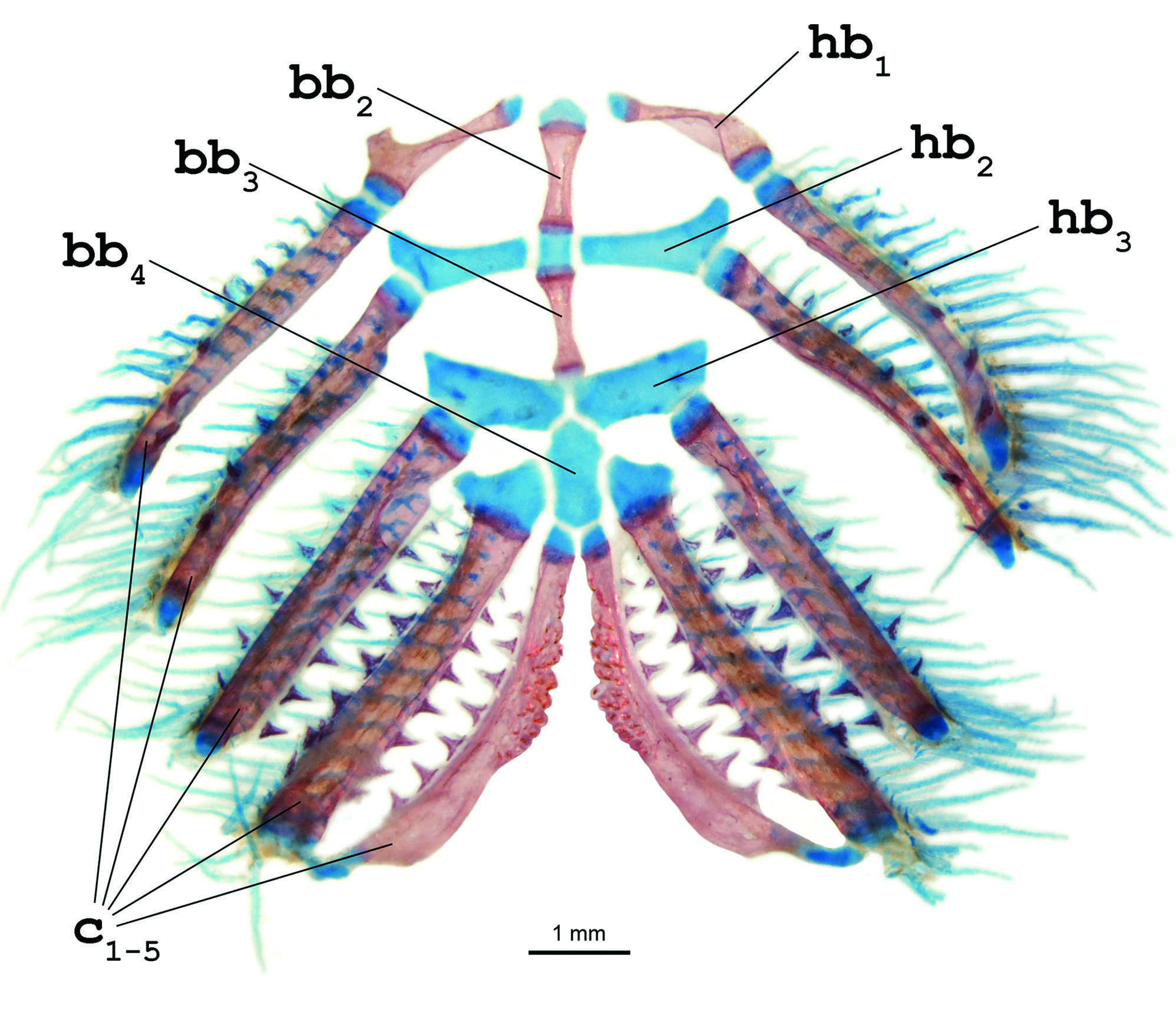
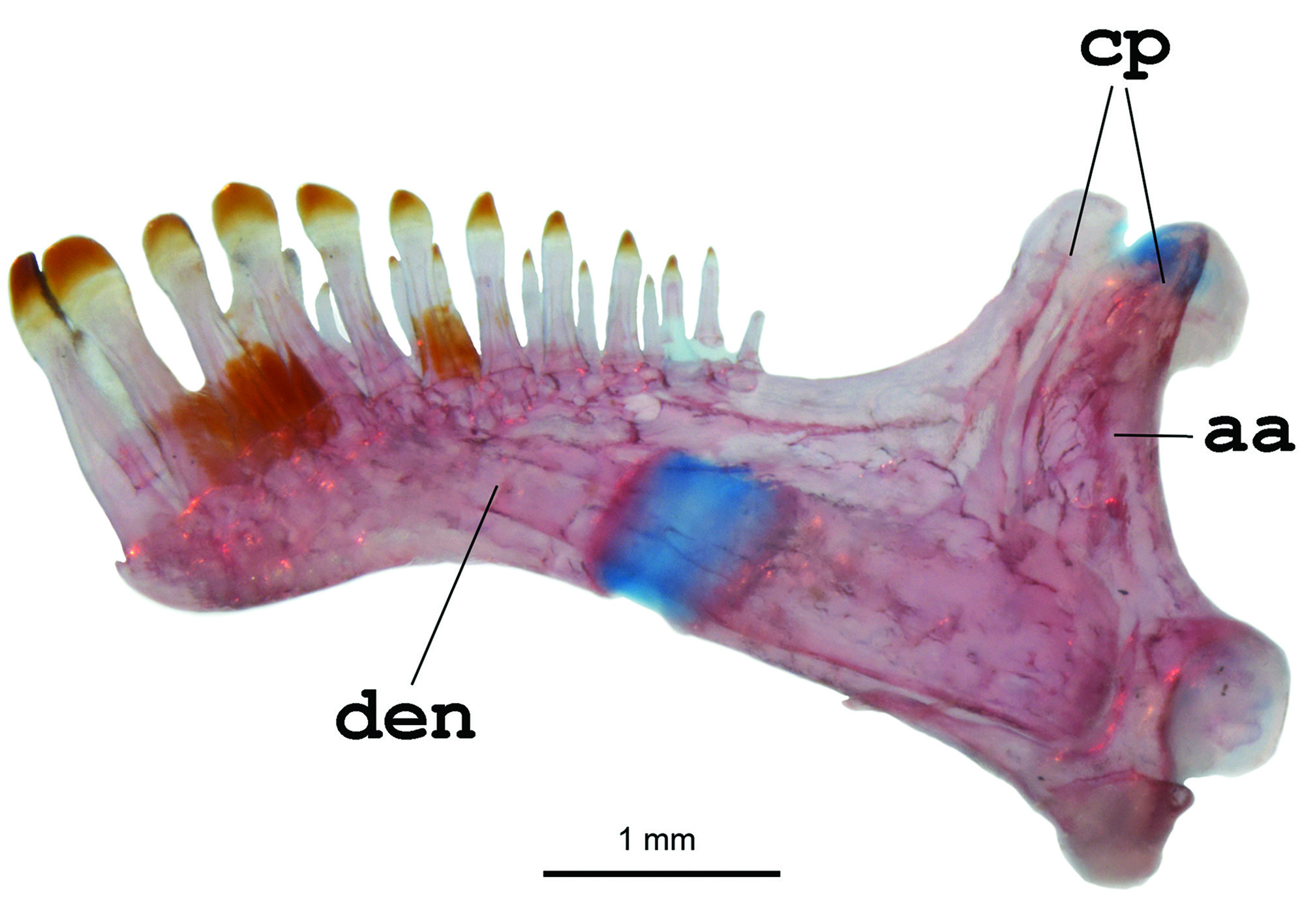



Table 1. Morphometric data for Copionodon elysium, new species. N = 30; SD = standard deviation.
| Holotype | Range | Mean | SD | |
|---|---|---|---|---|
| Standard length in mm | 45.0 | 32.5-71.5 | 44.5 | - |
| Percents of standard length | ||||
| Total length | 114.4 | 114.4-122.5 | 118.9 | 1.8 |
| Body depth | 17.3 | 13.5-18.4 | 16.0 | 1.4 |
| Predorsal length | 56.9 | 56.7-61.7 | 59.0 | 1.2 |
| Preanal length | 65.8 | 61.5-67.0 | 65.1 | 1.3 |
| Prepelvic length | 53.3 | 50.3-55.1 | 52.4 | 1.2 |
| Caudal-peduncle length | 19.8 | 16.5-21.0 | 18.9 | 1.0 |
| Caudal-peduncle depth | 14.9 | 11.3-14.9 | 13.0 | 0.8 |
| Dorsal-fin base length | 10.7 | 8.0-10.7 | 9.5 | 0.7 |
| Anal-fin base length | 16.4 | 14.4-18.4 | 16.3 | 1.0 |
| Head length | 22.2 | 21.3-23.7 | 22.4 | 0.6 |
| Percents of head length | ||||
| Head width | 98.0 | 89.6-100.0 | 92.9 | 3.4 |
| Head depth | 54.0 | 46.1-54.6 | 49.3 | 2.4 |
| Snout length | 56.0 | 48.0-57.6 | 52.5 | 2.3 |
| Mouth width | 40.0 | 34.3-44.8 | 36.3 | 2.2 |
| Orbital diameter | 17.0 | 12.2-19.7 | 16.5 | 1.9 |
| Interorbital width | 50.0 | 45.1-53.0 | 49.3 | 1.9 |
| Length of interopercular patch of odontodes | 34.0 | 31.7-39.6 | 35.2 | 2.0 |
| SL |
University of Sierra Leone, Njala University College |
| V |
Royal British Columbia Museum - Herbarium |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.