
Calliophis haematoetron, Smith, Eric N., Manamendra-Arachchi, Kelum & Somaweera, Ruchira, 2008
|
publication ID |
https://doi.org/10.5281/zenodo.183338 |
|
DOI |
https://doi.org/10.5281/zenodo.6236000 |
|
persistent identifier |
https://treatment.plazi.org/id/03EC87B6-0F54-FF8A-B1CD-FC4DFD430D93 |
|
treatment provided by |
Plazi |
|
scientific name |
Calliophis haematoetron |
| status |
sp. nov. |
Calliophis haematoetron sp. nov.
( Figs. 1–3 View FIGURE 1 View FIGURE 3 )
Calliophis melanurus ( Taylor 1950: 583–585, in part, described specimens) ( Bahir 1999: 22–24, in part) Calliophis melanurus sinhaleyus ( Deraniyagala 1951: 147–148, in part, referred specimens of new taxon, at least specimens 12–13 on table 2) ( Das & De Silva 2005: 57, in part)
Calliophis sp. ( Somaweera 2006: 154–155)
English name: Blood-bellied coralsnake Holotype — WHT (The Wildlife Heritage Trust of Sri Lanka, Colombo, Sri Lanka) 1621, an adult female from Wasgomuwa National Park, [Central Province], Sri Lanka, ca. 90 m (ca. 7.648056° N 80.93583° E), collected 7 June 1997 by Mohomed M. Bahir and Sampath Nanayakkara (figs. 1–3).
Paratypes (2) — USNM (National Museum of Natural History, Smithsonian Institution, Washington D.C., United States of America) 120334 – 120335, young male and adult female, respectively, from Clodagh Estate, Rattota, Matale [Central Province], Sri Lanka, ca. 570 m (ca. 7.521667° N 80.68472° E), collected on 2 July 1944 by Herbert G. Deignan (fig. 2).
Diagnosis — A small ( 152–414 mm TL, young male and mature female, respectively), brownish, terrestrial coralsnake in which the tail comprises 10.5% of the TL in the known male and 8.0–8.6% in the two known females. It has no sublabial-chin-shield contact, 6 supralabials, 6 infralabials, 2 postoculars, 225–239 ventrals, a divided anal, 29–35 divided subcaudals, dorsal scale rows arranged in 13 rows along entire body, and a color pattern consisting of 13–22 lateral body blotches and 2–3 tail bands (including one band in the cloacal region).
The new species can be distinguished from the only other known Sri Lankan Calliophis , C . melanurus, in having a frontal that is about equal in length or slightly shorter (vs. longer) than the interparietal suture, a first sublabial that does not touch the second pair of chin-shields (vs. broad contact with second pair of chinshields), a relatively unpigmented head (vs. capped with black from rostrum to nuchal collar), no light spots posterolateral to the parietals, numerous bands on the dorsum of the body (13–22 vs. none; excluding nuchal collar and cloacal and tail bands), a blood-red body venter (vs. orange or yellow through most of the length), and in having red pigment lateral to blue coloration under the tail (vs. no red on tail).
The new species can be distinguished from all other coralsnakes, except C. melanurus and Calliophis maculiceps (Günther, 1858) , by its melanized tail base internal musculature, hypaxial, and associated epimysial and perimysial tissues (i.e., asulcate layer of hemipenis, m. propulsor, m. retractor penis magnus, and homologous muscles in females). The new species can be distinguished from all other Calliophis species in nearby India, C. beddomei ( Smith, 1943) , C. bibroni (Jan, 1858) , C. melanurus , and C. nigrescens (Günther, 1862) . From C. beddomei it differs in having fewer supralabials (6 vs. 7), first sublabials not touching the second pair of chin-shields, a pale head with light-brown subocular and cheek blotches (vs. head with melanic color, covering the snout and continuing caudally as a narrow interparietal line joining the dark collar and as lateral stripes from above the eye to the corner of the mouth), a body pattern of complete and incomplete bands and a few lateral anterior blotches (vs. pattern of paradorsal and lateral blotches separated by faint and broken medial stripe), a colorful venter (vs. immaculate light colored, including tail), and tail bands (vs. no bands, only small blotches). From C. bibroni it differs in having fewer supralabials (6 vs. 7), a light colored (vs. dark) snout, a preocular (vs. no preocular), two postoculars (vs. 1), 2 or 3 tail bands (vs. 4–6), thin dark bands on the body that are up to 2 scales wide (vs. usually 3–4 scales wide), a divided anal (vs. single), and higher ventral scute counts (males 225 vs. 219; females 232–239 vs. 222–227). It differs from C. nigrescens in having a first sublabial that does not contact the second pair of chin-shields (vs. broad contact), having 6 supralabials (vs. 7), dark subcaudal markings (vs. unmarked), and a dorsal pattern of bands (vs. striped, with longitudinal blotches, or unicolored). The new species differs from Calliophis gracilis Gray, 1835 in possessing fewer ventral scales (225–239 vs. 303–320), more subcaudals (29–35 vs. 21–23), 2 maxillary teeth posterior to fangs (vs. none), a dorsal pattern of narrow bands (vs. large and paired paravertebral spots and 5–7 well-defined stripes), and a venter with no bands (vs. numerous regularly spaced wide bands). From C. maculiceps it can be distinguished by its high number of ventrals (225–239 vs. 169–222), more subcaudals (29–35 vs. 20–31), lack of postocular stripes, the first sublabial not touching the second pair of chin-shields (vs. broadly touching), and a frontal that is shorter or about equal to the length of the interparietal suture (vs. longer).
From the long-glanded coralsnakes Calliophis bivirgata (Boie, 1827) and Calliophis intestinalis (Laurenti, 1768) , previously known as members of a the genus Maticora Gray, 1834 (see Slowinski et al. 2001), the new species differs in having a venom gland that is confined to the temporal region (vs. extending behind the head), a Harderian gland with a moderately developed posterior extension (vs. enlarged posterior extension, larger than the eyeball), pterygoids of moderate size and with 4 teeth (vs. reduced in size and with 2, 1, or no teeth), and a dorsal pattern of bands (vs. stripes). Additionally, the new species can be distinguished from C. bivirgata in lacking a bright red venter, head and tail dorsum, and from C. intestinalis in lacking a venter without bars and a blue (vs. red) subcaudal coloration.
From species in the genus Sinomicrurus Slowinski, Boundy & Lawson, 2001 , i.e., S. hatori (Takahashi, 1930) , S. japonicus (Günther, 1868) , S. kelloggi (Pope, 1928) , S. macclellandi (Reinhardt, 1844) , and S. sauteri (Steindachner, 1913) ( sensu Slowinski et al. 2001) , the new taxon differs in possessing no protuberant sclerified tail tip, 6 (vs. 7) supralabials, and a Harderian gland with a moderately developed posterior extension (vs. no extension). It can further be distinguished from S. hatori , S. japonicus , and S. sauteri in having no pattern of stripes, and from S. kelloggi and S. macclellandi in having no white band anterior to the nuchal band. From snakes in the genus Hemibungarus Peters, 1862 , namely Hemibungarus calligaster (Wiegmann, 1834) , that had previously been considered coralsnakes (e.g., Slowinski et al. 2001), the new species differs in having 13 dorsal scale rows (vs. 15), 1+2 temporals (vs. 2+2 or 2+3), more subcaudals (32–35 vs. <25), a divided anal plate (vs. single), and a Harderian gland with a moderately developed posterior extension (vs. no extension). Hemibungarus calligaster is now placed in the tribe Hemibungarini, and the genus Calliophis and the rest of the Asian and American coralsnakes in the tribe Calliophini, based on hemipenial and molecular data ( Castoe et al. 2007).
Etymology — The specific name, haematoetron , is derived from the Greek adjective haematodes (bloody), and the Greek noun etron (belly) meaning “bloody belly” in reference to the bright-red venter of this “beautiful snake” (= Calliophis ).
Description of holotype and variation — Features of the adult female holotype are followed in parentheses by variation of the young male and the mature female paratypes, respectively. Total length 371 mm (152, 414); tail length 32 mm (16, 33); head length 6.9 mm (4.8, 9.5) from anterior edge of rostral to posterior end of mandible; head width 5.8 mm (3.1, 5.3) at broadest point; head slightly distinct from neck; snout 2.0 mm (1.5, 2.7) from front of rostral to anterior edge of eye; eye 0.3 (0.4, 0.4) times length of snout; pupil round; rostral 1.4 (1.3, 1.8) times wider than high; internasals 2.0 (1.2, 1.3) times wider than long, contacting only the nasals laterally; length of internasal suture slightly more than half diameter of eye (half in female paratype); prefrontals as wide as long, in contact laterally with nasal, preocular, and supraocular; prefrontal suture 1.4 (1.0, 1.3) times diameter of eye; frontal 1.3 (1.5, 1.5) times longer than wide; supraoculars 1.4 (1.3, 1.8) times longer than wide; parietals twice as long as wide (2.0, 1.8); parietal suture 0.6 times length of parietals, 1.2 (1.1, 0.9) times longer than frontal; 1+0 temporals and one posttemporal, shields touching parietal laterally large and elongated; temporal 1.9 (2.2, 1.9) times longer than wide; single preocular, 1.6 (1.5, 1.4) times longer than wide, triangular, with apex rostrally, located above line between center of eye and posterior border of naris; two postoculars, upper slightly wider than lower, reaching beyond upper and lower borders of eye, respectively; no loreal, preocular and nasal in contact; 6/6 supralabials, sixth largest and longest, first in contact with anterior nasal, second in contact with both nasal plates; third in contact with posterior nasal, preocular, and one sixth of orbit; fourth below orbit and contacting lower postocular, fifth in contact with lower postocular and temporal, and sixth in contact with temporal; mental 1.7 (1.9, 1.7) times as broad as long; anterior chin-shields 2.1 (2.1, 2.0) times longer than wide; posterior chin-shields 2.3 (1.9, 2.0) times longer than wide; 6/6 infralabials, first pair in contact behind mental, second small, second and third touching anterior chin-shields, fourth largest and contacting anterior and posterior chin-shields, fifth and sixth contacting sublabials; first sublabial not touching chin-shields; 2 (2, 1) gulars and 2 (3, 4) preventrals at midline between posterior chin-shields and first ventral; with few tubercles on head scales, concentrated anteriorly; dorsals in 13 rows, smooth, unreduced; apical pits absent; ventrals 239 (225, 232); anal divided; preanal single; subcaudals 32 (35, 29), paired; tail complete, tip round; no anal ridges or tubercles; male with umbilical scar on ventrals 161–163.
Dentition of paratypes examined, characteristics of male followed by variation of female, in parentheses: maxillae bearing one fang 0.50 mm long ( 1.1 mm), arising below suture of supralabials 2 and 3, slanted backward; two posterior maxillary teeth on each side, first largest, 0.13 mm long ( 0.15 mm), close to each other (0.13, 0.10 mm), slanted backward, below suture of supralabials 4 and 5, first at 1 mm from base of fang ( 1.2 mm); 6/6 palatine teeth (7/6); 4/4 pterygoid teeth (6/5); 9/9 dentary teeth, decreasing in size from front to rear. Head glands visible through transparent head shields of young male and when reflecting head skin of female paratype (skin detached prior to our examination; characteristics of male followed by variation of female, in parentheses): salivary gland developed under supralabials 1–3; nasal gland occupying area below prefrontal shields (prefrontal shields, posterior nasal, and preocular); Harderian gland under anterolateral portion of parietal (and posterior supraocular and upper postocular), 0.68 mm long (1.00 mm), 0.60 mm wide ( 0.90 mm), triangular, apex caudal, with a moderate posterior extension; venom gland triangular (rounded posteriorly), corners at middle of fourth supralabial (middle of third supralabial), middle of sixth supralabial (tip of temporal), and middle of temporal at border with parietal, 0.78 mm wide ( 1.95 mm), 1.88 mm long ( 4.85 mm), not inflected ventrally and confined to head; infralabial gland bordering mouth under lateral tips of mental to fifth infralabial, area differentiated, darker and more granular under fourth infralabial; in male, salivary, nasal, Harderian and infralabial glands yellowish and of irregular texture (granular), venom gland whitish and smooth; m. adductor mandibulae externus superficialis (AES) forming continuous loop, from upper parietal surface above Harderian gland to insertion on compound bone; in both specimens, granular glands situated under the rostral; in female, slender gland evident under lower edge of supralabials five and six.
Hemipenes of male exposed in situ but not dissected or cut, single (unbifurcated), relatively smooth, reaching level of subcaudal 10; each hemipenis and associated m. retractor penis magnus covered in a melanic epimysium, asulcate layer; m. retractor penis magnus inserts first to vertebrae at level of subcaudal 32; m. propulsor also with external melanized epymisium; no spines or papillae evident through transluscent tissue (after removing the melanic sheath); subvertebral and medial hypaxial musculature in tail with melanized internal epymisial covering; cloacal scent glands oval, ending at levels of subcaudals 3 (left) and 4 (right). In female paratype, tail subvertebral and medial hypaxial musculature and associated epimysial and perimysial connective tissues melanized; external epymisial tissue covering m. propulsor and basal and medial section of m. constrictor sacculi ani melanized.
Color ( Fig. 1 View FIGURE 1 ) — Holotype in life: Dorsum of head and body Burnt Sienna (132), turning Drab-Gray (119D) towards venter; Jet Black (89) dorsal bands and blotches, those anterior finely edged with Pale Pinkish Buff (121D); Jet Black irregular diffuse reticulation on top of head and suborbital and cheek markings; dark cheek marking covering almost all sixth supralabial and continuing through lower edge of temporal and posttemporal towards black collar; eye surrounded by dark pigment; 22 primary bands on body (crossing midline or not), about 1.5–2 scales wide; five small middorsal blotches anteriorly and small diffuse and scattered blotches posteriorly; three dorsal bands on tail; scales on second dorsolateral row with dark centers; tip of tail light colored; no spots on ventral scales; venter of body to anal plate and first row of dorsal scales on tail Gem Ruby (110) red; underside of tail True Blue (168A), except for last two subcaudals which are Gem Ruby and Pearl Gray (81); 19 subcaudals blotched with Jet Black.
Holotype in preservative: Drab Gray dorsally and ventrally; head markings Warm Sepia (221A) to Sepia (119); nuchal band and dorsal body and tail bands and blotches Sepia; faint middorsal stripe Warm Sepia; dark subcaudals Jet Black.
Male paratype in preservative similar to holotype, differs in having 13 body dorsal bands, two tail dorsal bands, a Light Drab (119C) overall coloration, Dark Drab (119B) chin, Sepia (219) bands and blotches, 12 subcaudals with dark lateral coloration (tail ventrally almost immaculate), and only faint suborbital and cheek blotches.
Female paratype in preservative more similar in color and pattern to preserved holotype than to male paratype; differs from holotype in possessing 17 body dorsal bands and two tail dorsal bands.
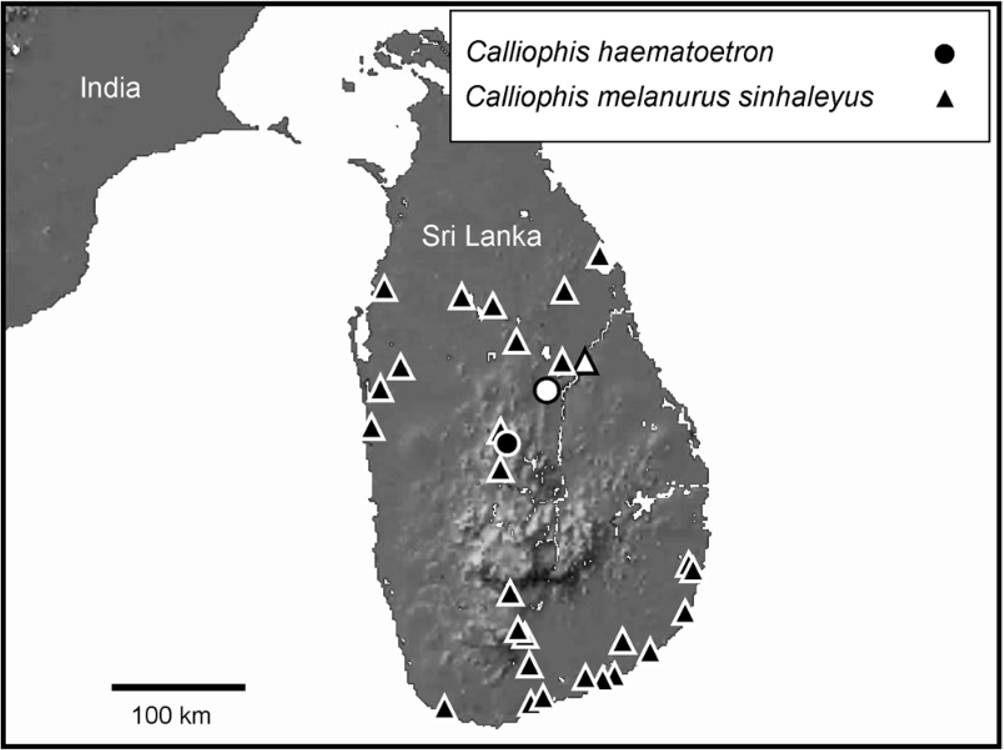
Habitat, distribution and natural history ( Figs. 4–7 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 ) — The two known localities for Calliophis haematoetron lie in two different types of forest. The holotype was collected at Wasgomuwa National Park, a tropical dry (mixed) evergreen forest, whereas the paratypes from Clodagh Estate, Rattota, come from an originally tropical moist semi-evergreen forest locality. Nowadays, however, no significant forest remnants of the latter formation remain ( Gunatilleke & Gunatilleke, 1990). The recorded altitudinal range of the new species is ca. 90– 570 m. The moment of capture and associated defensive behavior of the holotype was described by Bahir (1999): “Running my hand under the thick layer of brown leaves on the forest floor, I uncovered a long, brown snake, almost 40 cm (rather more than a foot) long. Immediately I disturbed it, it turned the underside of its tail up displaying a stunningly spectacular blue which instinctively had me letting go. As I tried to grasp it in my hand again, it turned its belly up at me all the while keeping its head concealed and the right way up, displaying a bright red underside.” Bahir (1999) also stated that “it moves with incredible stealth and speed through leaf litter.” The female paratype, collected 2 July, has yolked oviductal eggs with soft shells, two anteriorly in the left oviduct (anterior 22 mm, posterior 27 mm), and one posterior, in the right oviduct ( 30 mm). Egg deposition presumably happens later during the month of July.
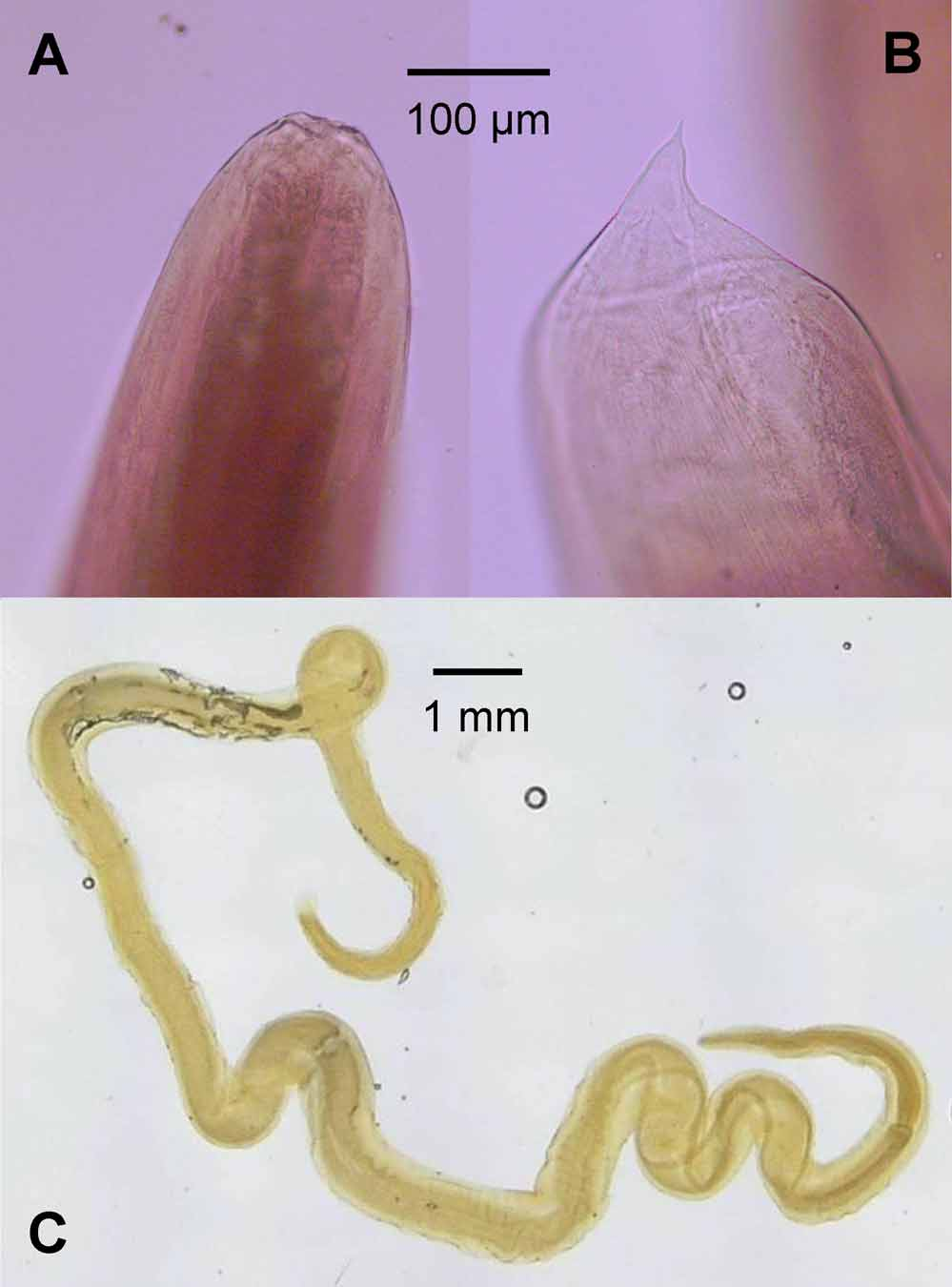
Endoparasite ( Fig. 8 View FIGURE 8 ) — The adult female paratype contained a single encysted nematode in the abdominal cavity; this larval individual endoparasite appears to belong to the family Spiruridae , a member of the superfamily Dracunculoidea Cameron, 1934. It possesses a long and relatively thin body, a rounded cephalic end with a cuticular shield and cephalic papillae, a reduced buccal capsule, an esophagus consisting of short muscular and long glandular parts, and an intestine that is straight and tubular ( Skryabin 1984). Numerous species of Spiruridae infect the mesentery, coelomic cavity, and blood vessels of snakes. This nematode could not be assigned to a lower taxonomic rank because it is in a larval stage and diagnostic features have not yet developed. The specific identity of this Spiruridae found in Calliophis haematoetron remains to be corroborated.
Relationships — Calliophis haematoetron is probably closely related to the lowland Calliophis with blue tail venters namely, C. melanurus from India and Sri Lanka and C. maculiceps from southern Myanmar, Thailand, Cambodia, and northern peninsular Malaysia. These species share melanized musculature and tissues at the base of the tail, blotching in some of the subcaudals, tail bands (usually two), and a body venter with red or orange, particularly towards the anal plate.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.