
Boreviulisoma barrocalense, Reboleira, Ana Sofia P. S. & Enghoff, Henrik, 2013
|
publication ID |
https://doi.org/10.11646/zootaxa.3646.5.2 |
|
publication LSID |
lsid:zoobank.org:pub:649D8424-F543-4489-B025-E8006D1EEB52 |
|
DOI |
https://doi.org/10.5281/zenodo.5691641 |
|
persistent identifier |
https://treatment.plazi.org/id/F29BAE99-2F6C-4DF2-A0D9-E7A1242D3A43 |
|
taxon LSID |
lsid:zoobank.org:act:F29BAE99-2F6C-4DF2-A0D9-E7A1242D3A43 |
|
treatment provided by |
Plazi |
|
scientific name |
Boreviulisoma barrocalense |
| status |
sp. nov. |
Boreviulisoma barrocalense View in CoL n. sp.
Figs 1–14 View FIGURE 1 View FIGURES 2 – 6 View FIGURES 11 – 14 , 27 View FIGURE 27
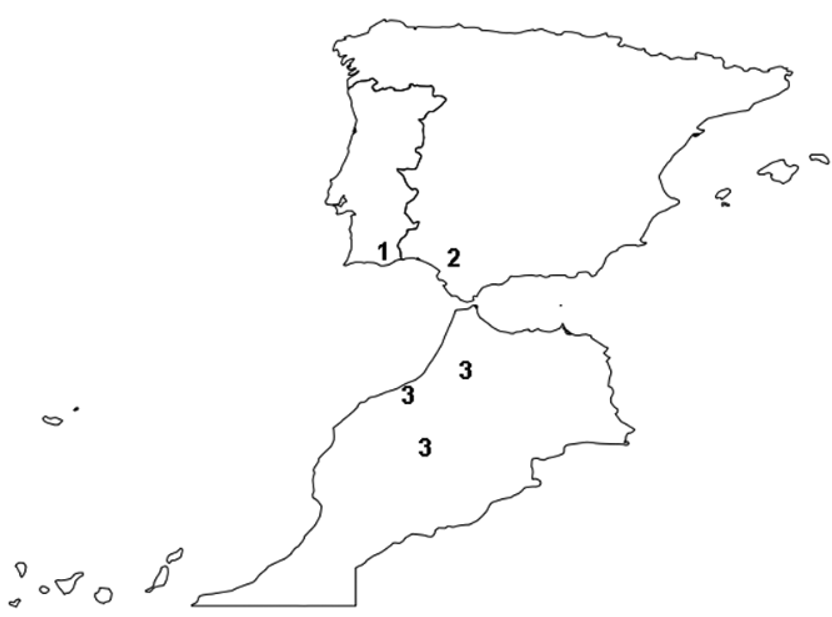
Material examined: Holotype 3, PORTUGAL, Algarve, Loulé, Gruta do Vale Telheiro, 239 m a.s.l., 30.VI.2011, S. Reboleira leg. (ZMUC). – Paratypes: 1 Ƥ, same data (ZMUC); 1 3, 1 Ƥ, same data, but 13.III.2009 (ZMUC); 1 Ƥ, same data, but 24.V.2009 (DZUL).
Name: The species epithet is a Latin adjective referring to the subregion Barrocal in the Algarve, in which the type locality is situated.
Diagnosis. Differs from congeneric species by lacking pigment, by having femoral knobs on male legs 3–7 and 9–10, and by having a hawksbill-like process on the dorsal side of the gonopod tip. Further differs from B. liouvillei by having simple, pointed metazonal setae.
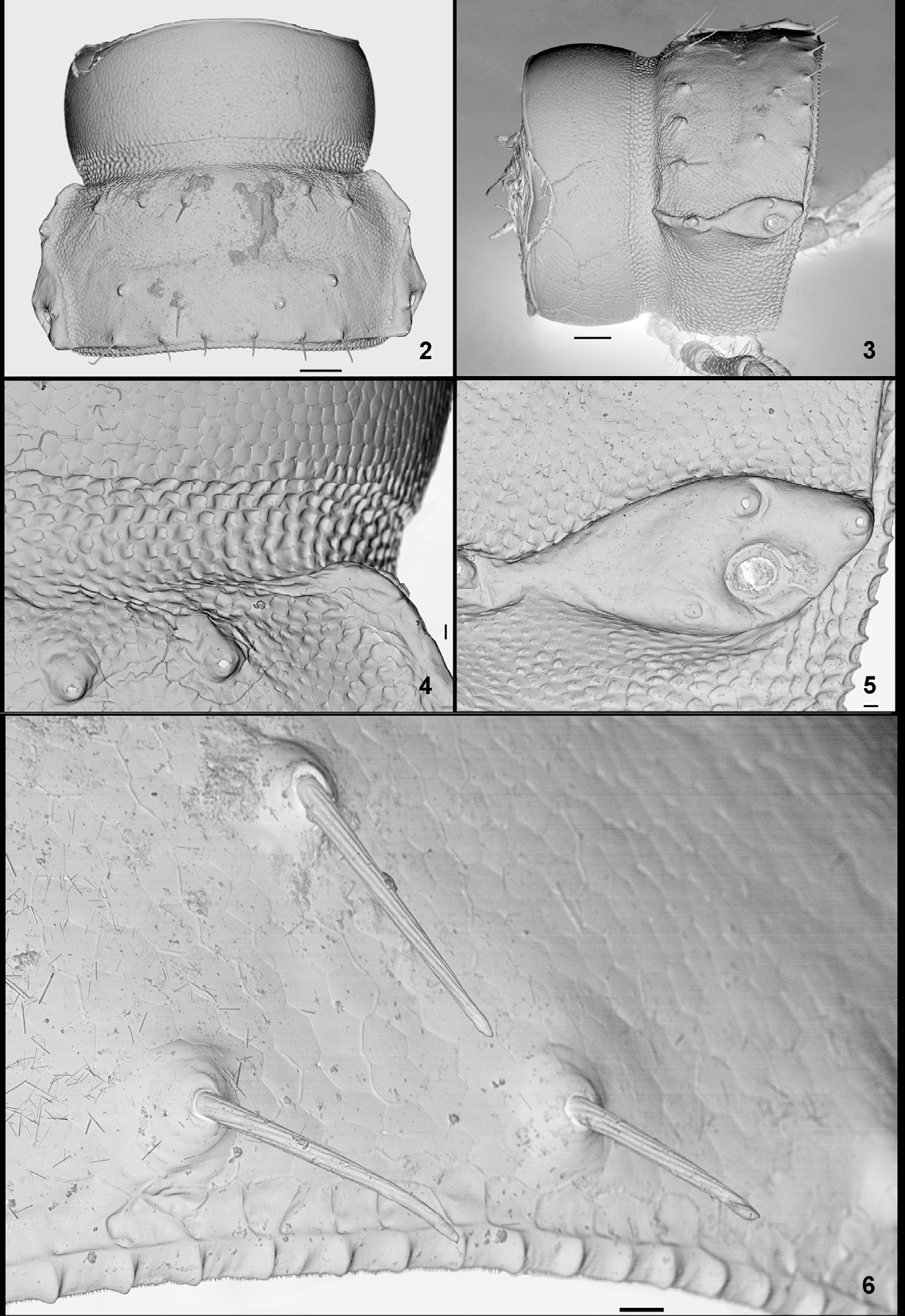
Description. Body length 9–10 mm in 3, 10–11 mm in Ƥ. Width across midbody prozona/metazona 0.67– 0.68/0.86–0.89 mm in 3, 0.73–0.87/0.99–1.04 mm in Ƥ. 20 “segments” (18 podous + 1 apodous rings + Telson). Completely depigmented ( Fig. 1 View FIGURE 1 ).
FIGURES 7–10. Boreviulisoma barrocalense n. sp., 3 paratype, 7—right 5th leg. 8—SEM image, left 7th leg, femur and postfemur. 9—do., detail of femoral tubercle. 10— holotype, right gonopod, mesal view. Scale bars: 7, 10—100µm; 8—10 µm and 9—1 µm.
Collum with small paranota, not reaching down to level of paranota of ring 2, the latter at same level as those of following rings; collum with three transverse rows of slender setae.
Body rings ( Figs 2–6 View FIGURES 2 – 6 ) regularly vaulted, slightly constricted between pro- and metazona; stricture with coarse, scalelike microsculpture sharply delimited against normal sculpture on rest of prozona. Metazona with fine punctate microscupture and three transverse rows of 8, 6 and 8 small tubercles, each with a slender, pointed seta. Limbus cellular, each cell with numerous very small denticles at posterior margin. Pleurosternal keels very poorly developed. Paranota narrow, set at ca. mid-height of rings, their margins with two blunt-angled protrusions, posterior corners barely reaching behind hind margin in posteriormost rings. Ozopores in ovoid peritremata in posterior ¼ of metazona. Sterna unmodified (cf. genus diagnosis). Telson unremarkable, preanal process triangular with bluntly rounded tip, subanal scale evenly rounded.
Male legs 5–7, as well as first and second postgonopodal legs (9th and 10th pairs of appendages) with subdistal femoral tubercles (Figs 7–9). Legs otherwise unmodified.
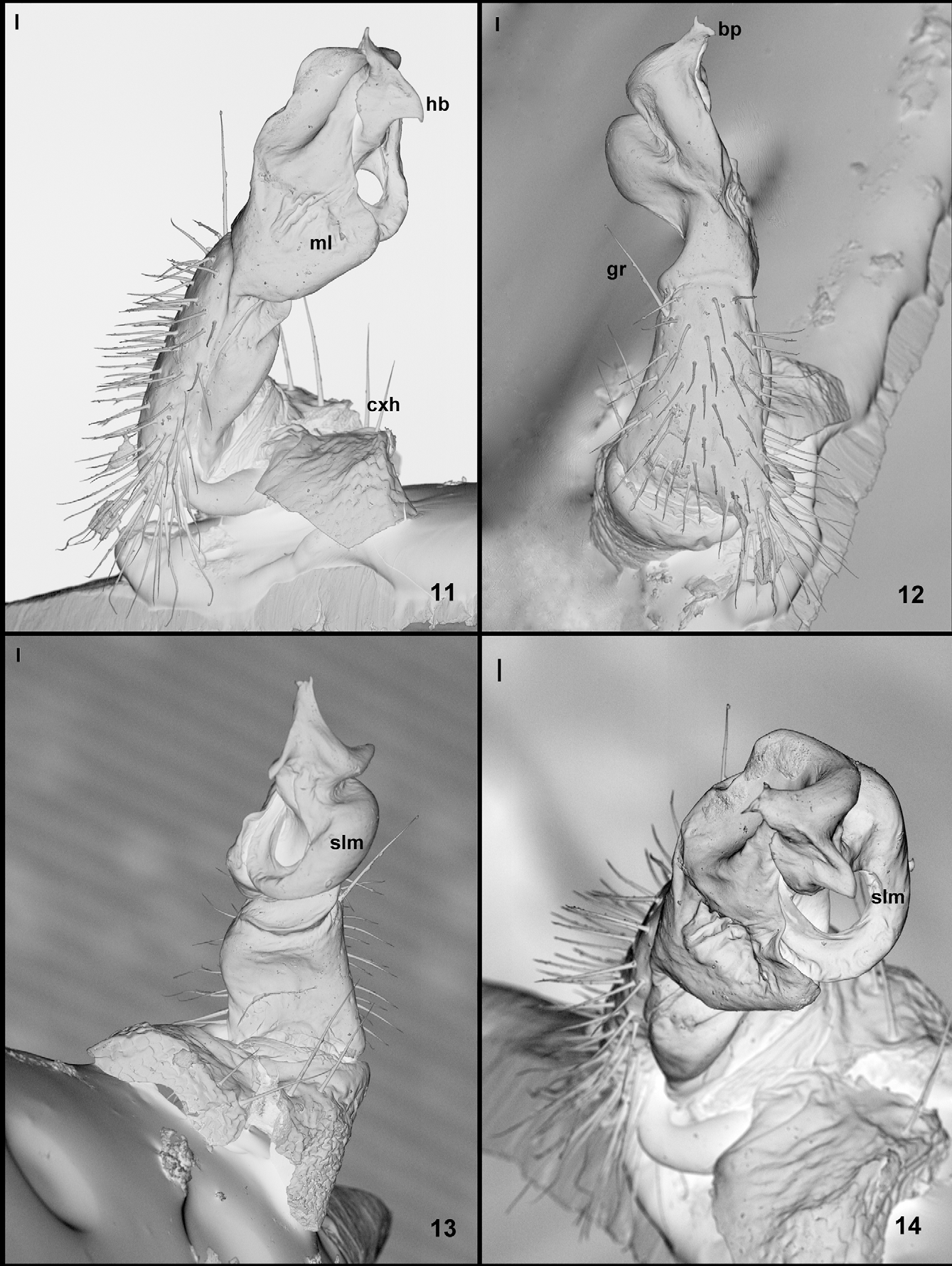
Gonopods (Figs. 10–14) very similar to those of congeneric species. Coxa distally with a large anterior hump (cxh). Telopodite at right angles to coxa; prefemoral part hypertrophied, as usual with numerous long setae on ventral side; femorite very short, distally delimited on ventral side by transverse groove (gr); distal part of telopodite consisting of a simple, sinuous solenomere (slm); solenomere originating laterally, curving posteriad, then mesad, then anteriad, its distal part hidden from view by large, mesal lamella (ml) of solenophore; solenophore complicated, torsate, with a large, rounded mesal lamella (ml), a subdistal, dorsal, hawksbill-like process (hb) and a terminal bifid process (bp). Efferent canal (= seminal groove, cf. Enghoff 2011) starting mesally, then running across telopodite’s dorsal edge to lateral side before entering the solenomere.
Habitat and ecological remarks. The Algarve is considered an area of high endemism for arthropods (Gama et al. 1999), while the subterranean ecosystem of the Algarve has been a refuge for several biological relicts over time (Reboleira et al. 2011).
From a biogeographic point of view, the Algarve has a complex history, and its troglobiont fauna reflects different colonization events, related to the isolation of the Iberian Peninsula, its relative recent connection to the north of Africa and the migration of Eastern European lineages, as well as some enigmatic faunal elements (Reboleira 2012, Reboleira et al 2010a, 2010b, 2012a, 2012b).
Boreviulisoma barrocalense n. sp. is the first troglobiont species of the genus. The type locality, Vale Telheiro cave, is currently the richest in hypogean species of Portugal (Reboleira 2012). It is located in the Barrocal area with a natural vegetation cover on the surface and it harbours a subterranean community composed of 15 caveadapted species. Boreviulisoma barrocalense n. sp. shares its habitat with another troglobiont millipede, Acipes n. sp. Enghoff & Reboleira, in press ( Blaniulidae ). Besides these two millipedes, the subterranean biocoenosis is composed of the following troglobionts: 3 pseudoscorpions ( Chthoniidae , Bochicidae , Neobisiidae ), 3 spiders ( Leptonetidae , Linyphiidae , Dysderidae ), 1 centipede ( Lithobiidae ), 1 oniscidean isopod ( Trichoniscidae ), 1 dipluran ( Campodeidae ), 1 silverfish ( Nicoletiidae ) and 3 beetles ( Staphylinidae , Leiodidae , Curculionidae ) (Reboleira 2012) and some frequent troglophiles such as gastropods, the isopod Porcellio dilatatus Brandt, 1833 and a large japygid dipluran.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |