
Drymarchon couperi
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4695.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:80E01C99-D9F2-49B6-9B4E-0929F8723E81 |
|
DOI |
https://doi.org/10.5281/zenodo.4323917 |
|
persistent identifier |
https://treatment.plazi.org/id/03ED606B-8B19-FFC9-FF7E-FB3EB305FC34 |
|
treatment provided by |
Felipe |
|
scientific name |
Drymarchon couperi |
| status |
|
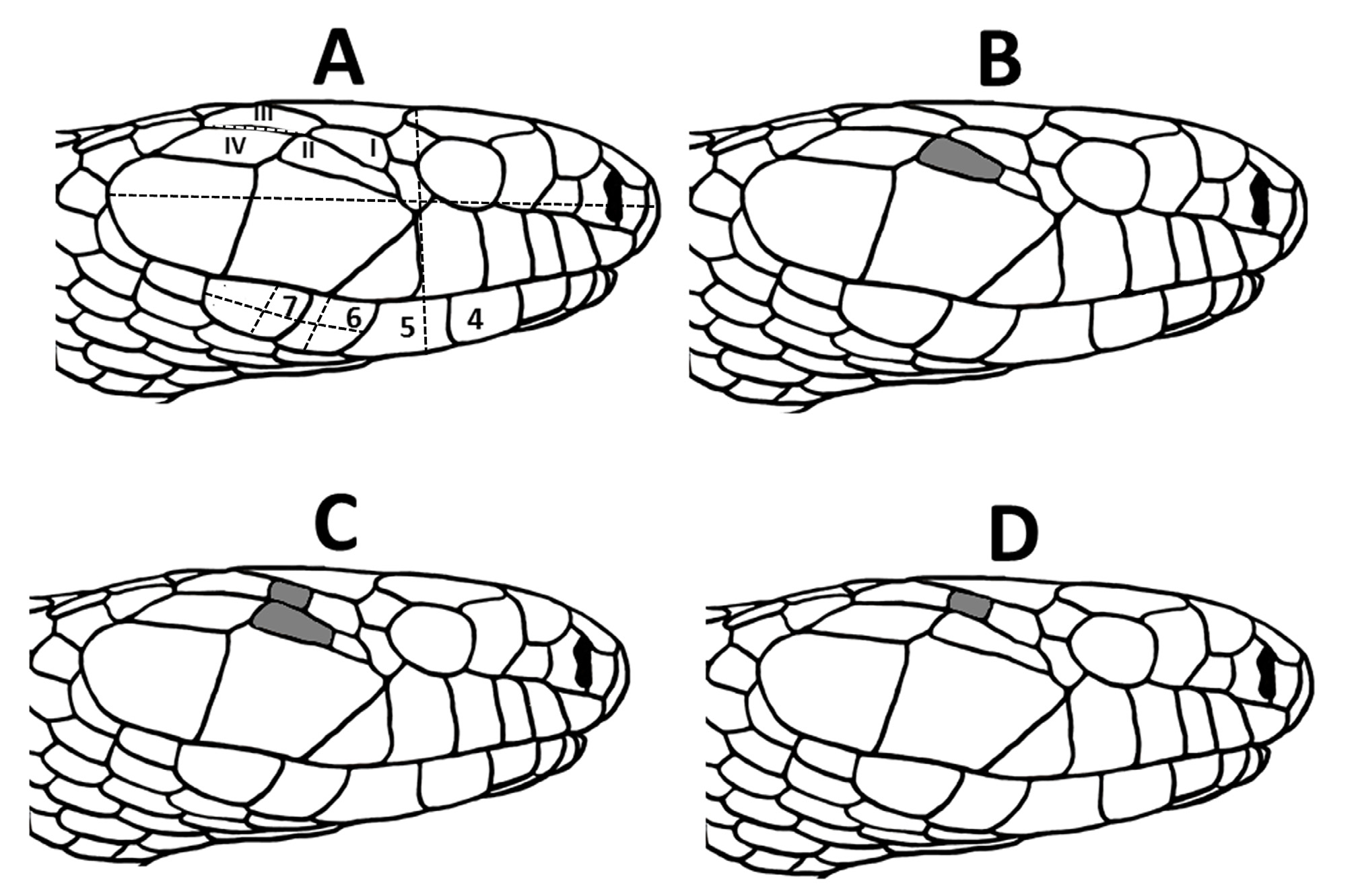
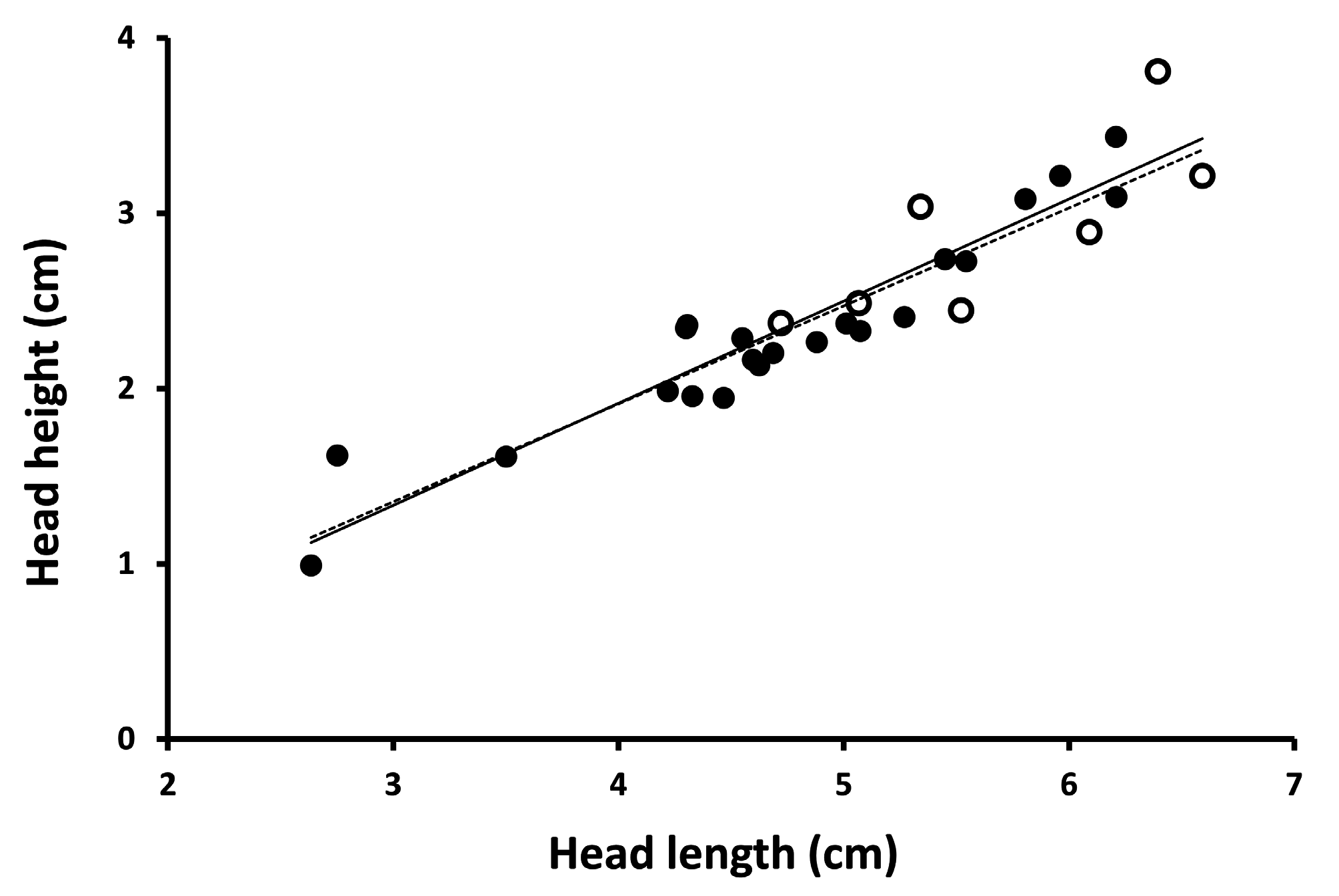
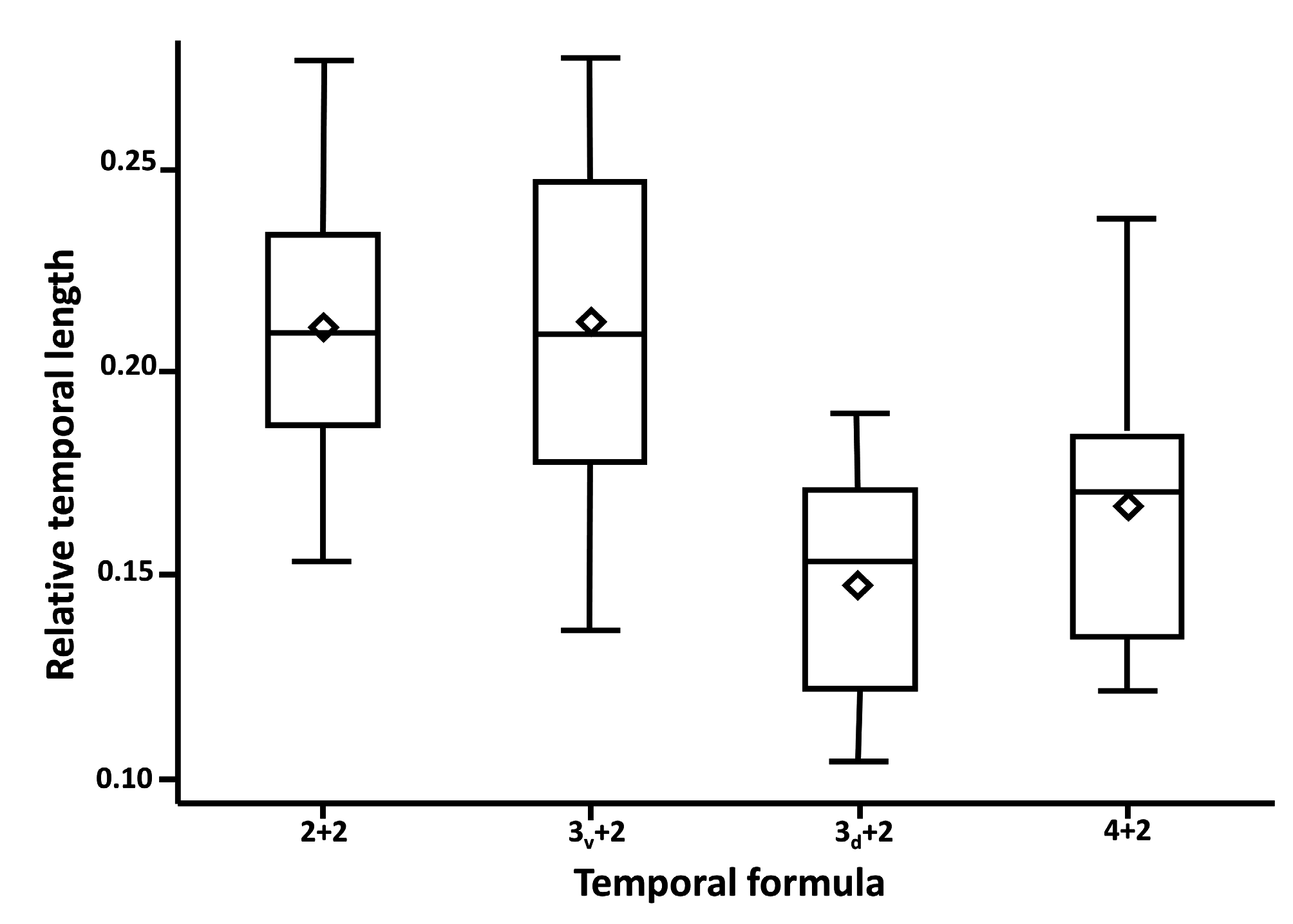
examined snakes housed at the Orianne Center for Indigo Conservation (OCIC) that were raised as stock for repatriation efforts. Our sample included 58 individuals likely representing all three genetic populations associated with the Atlantic lineage (Atkinson, Bryan, Evans, Long, Telfair, Wayne, and Wheeler counties, Georgia, USA) and 16 individuals likely representing two of three genetic populations of the Gulf Coast lineage (Citrus, Highlands, and Marion counties, Florida, USA; lineages from Krysko et al. 2016a; genetic populations from Folt et al. 2019b). We photographed lateral or dorsolateral aspects of the head of each specimen including a millimeter ruler for scale. From the images, we categorized the condition of the temporal scales for each specimen based on four character states created by the number and position of the scales ( Fig. 1 View FIGURE 1 ). We generated a contingency table providing counts of specimens in each of the four categories for each lineage. We used Chi-square analysis to determine whether the relative proportions of temporal scale categories differed between the Atlantic and Gulf Coast lineages. Additionally, we measured total head length (posterior-most point of 8th supralabial to anterior tip of rostral; n = 74), head height (only for photos in lateral aspect; at level of anterior-most point of parietal suture; n = 35), and length of the dorsal posterior-most temporal (intersection of ventral posterior-most temporal, dorsal posterior-most temporal, and adjacent first dorsal scale to intersection of ventral posterior-most temporal, dorsal posterior-most temporal, and adjacent ventral temporal; n = 74; Fig. 1A View FIGURE 1 ). We measured all distances using Adobe Photoshop 6.0 with reference to the photographed ruler. We used analysis of covariance (ANCOVA) to test whether the linear relationship between head length and head height differed between Atlantic and Gulf Coast lineages. We divided the length of the dorsal posterior temporal by head length to control for effects of body size and used an analysis of variance to test whether adjusted temporal length differed among the four categories of temporal scales.
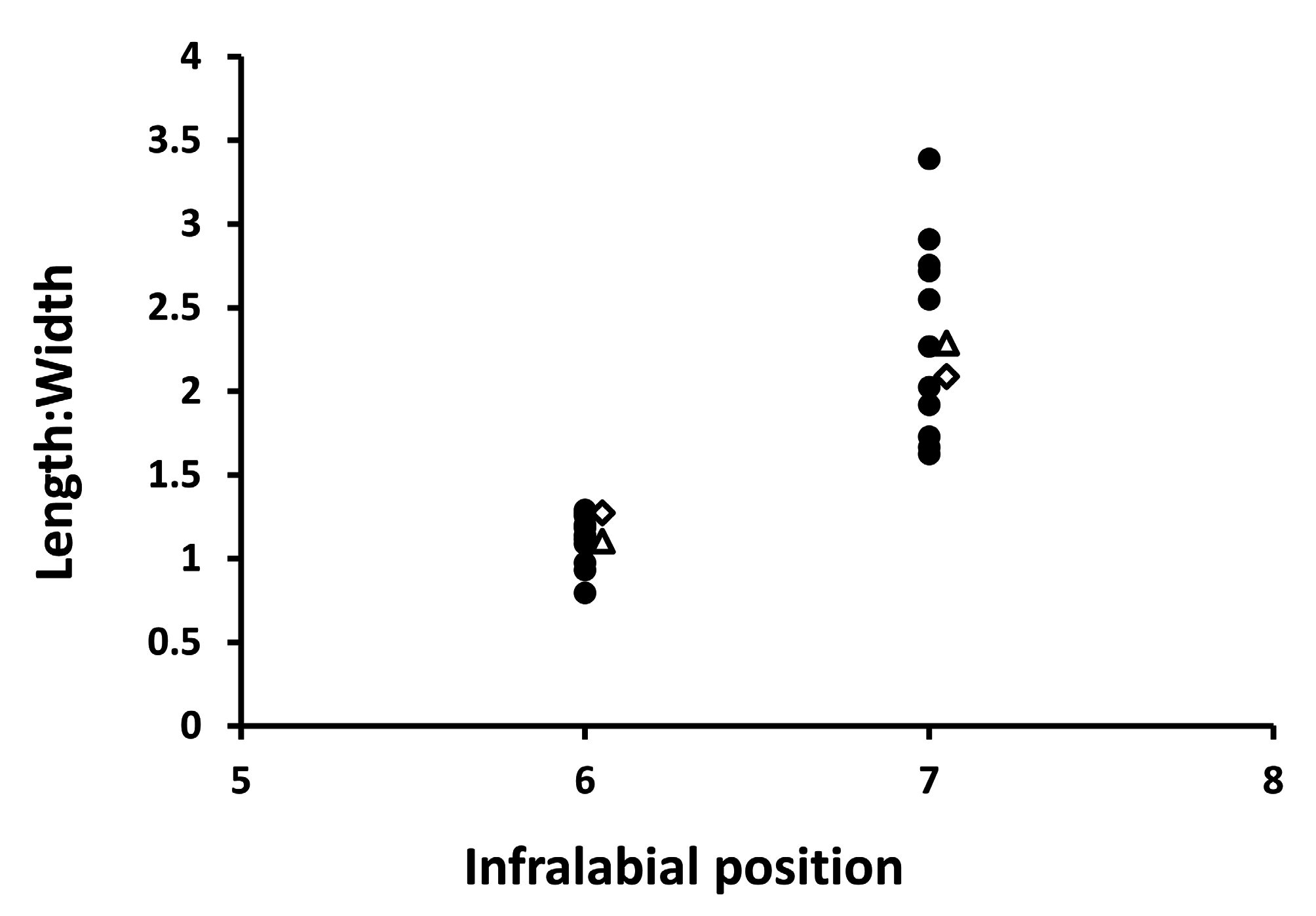
We also examined 11 preserved specimens in the Auburn University Museum collections (AUM 18437, 18444, 34209, 34216, 38868–69, 40744, 40750, 40752, 40815, and 42334). Because these snakes were from southeastern Georgia, we assumed them to belong to the Atlantic lineage. For these specimens, we measured length and width of the 6th and 7th infralabial scales with dial calipers ( Fig. 1A View FIGURE 1 ). We measured both scales because it was not clear which of these was measured by Krysko et al. (2016b). Additionally , we used photos of the type specimens presented in Krysko et al. (2016b) to determine length and width of the 6th and 7th infralabial scales using Adobe Photoshop 6.0. A length-to-width ratio was then calculated for each specimen. Mean differences between 6th and 7th infralabials were tested as a paired t-test. Differences between our sample of Atlantic lineage snakes and the type specimens was determined by visual inspection. We used SAS v.9.4 for all morphological analyses ( SAS Institute, Inc 2013) with α = 0.05 .
Our sample of OCIC specimens recovered four categories of temporal scales from both Atlantic and Gulf Coast lineages ( Table 1). In 24% of specimens, temporals conformed to the 2+2 formula that Krysko et al. (2016b) described as being invariant ( Fig. 1A View FIGURE 1 ), with 38% of specimens exhibiting an extra ventral temporal ( Fig. 1B View FIGURE 1 ), 23% of specimens having extra dorsal and ventral temporals ( Fig. 1C View FIGURE 1 ), and 15% of specimens exhibiting an extra dorsal temporal ( Fig. 1D View FIGURE 1 ). The frequency with which these four categories occurred differed between Atlantic and Gulf Coast lineage specimens ( Table 1; χ2 = 12.11.; df = 3; p <0.01), with Atlantic lineage snakes tending to have conditions with two dorsal temporals and Gulf Coast lineage snakes tending to have conditions with three dorsal temporals. Head shape, based on ANCOVA of head height on head length, did not differ between Atlantic and Gulf Coast lineages in either slope ( Fig. 2 View FIGURE 2 ; F =0.07; df = 1; p = 0.79) or elevation (F = 0.48; df = 1; p = 0.49). Length of the dorsal posterior-most temporal, expressed as a proportion of head length, differed significantly among temporal categories (F = 18.34; df = 3; p <0.0001), with the dorsal posterior-most temporal being proportionately shorter when three dorsal temporal scales are present relative to when two dorsal temporal scales are present ( Fig. 3 View FIGURE 3 ). When the length and width of the 6th and 7th infralabial scales were converted to a length-to-width ratio, the distribution of our sample of scales from Atlantic lineage snakes encompassed values for both type specimens for each scale ( Fig. 4 View FIGURE 4 ). Length-to-width ratios differed between 6th and 7th infralabials (t = 8.07; df = 12, p <0.0001), with 7th infralabials being more elongate than 6th infralabials.
Our results reject the hypothesis that the Atlantic and Gulf Coast lineages of D. couperi ( Krysko et al. 2016a) are identifiable on the basis of aforementioned phenotypic characters. We reach this conclusion after examining the variables used by Krysko et al. (2016b) to diagnose each lineage. Of the disparities that emerge between our analyses and theirs, the conformation of the infralabials is the most problematic. The figures presented by Krysko et al. (2016b) and Enge and Krysko (2019) for the 6th infralabial show great promise for diagnosing lineages. However, we were struck by how dissimilar the 6th infralabial of Atlantic specimens appeared to be from the long and thin scale shape ascribed to them by Krysko et al. (2016b, fig. 5A). Our analyses demonstrate that the 6th and 7th infralabials differ in shape, that the shape of the 7th infralabial conforms to the shape ascribed to the Atlantic lineage, and that the shape of the 6th infralabial conforms to that ascribed to the Gulf Coast lineage. Given that it is unclear which of these scales was measured by Krysko et al. (2016b) and that the range of variation of each scale within a sample of Atlantic lineage snakes encompasses both type specimens, we raise the possibility that Krysko et al. (2016b) intended to measure the 7th infralabial but inadvertently measured the 6th for Gulf Coast lineage specimens and the 7th for Atlantic lineage specimens. If the mental scale was included in the count for one lineage but not the other, this would provide a plausible explanation for their strong separation of the two lineages based on this scale and an inability for this to translate into a useful difference in character states in our analysis.
Our results for the temporal scale reveal great variation in the number of these scales present in Eastern Indigo Snakes. The four categories that characterize this variation are found in both Atlantic and Gulf Coast lineage snakes, in- dicating that this feature is not diagnostic. Nevertheless, Atlantic lineage snakes tend to have two dorsal temporals while Gulf Coast lineage snakes tend to have three. We assume that Krysko et al. (2016b) intended to measure the dorsal posterior-most temporal and, therefore, we focused our attention on this scale. Our data indicate that the length of the dorsal posterior-most temporal, relative to head length, becomes shortened if three dorsal temporals are present and becomes elongate if two dorsal temporals are present. This finding indicates that the scale shapes revealed by Krysko et al. (2016b) represent distinguishable groups, but that the groups are created by numbers of temporal scales present rather than representing two species. We speculate that the different morphologies of the dorsal posterior-most temporal result because, during embryonic development of some individuals, the dorsal anterior temporal divides, limiting space for development of the dorsal posterior-most temporal. Because these two conditions of the dorsal temporals are present in both Atlantic and Gulf Coast lineages the two lineages cannot be diagnosed by relative lengths of temporal scales.
Krysko et al. (2016b) also used head shape to diagnose the two lineages, with Atlantic lineage snakes having an elongate deep head and Gulf Coast lineage snakes having a short narrow head. Our bivariate examination of head length and height revealed no difference in head shape between the two lineages. We have no convenient explanation for this disparity, except to note that head height is difficult to measure consistently. Specimens preserved with mouths open are likely to have larger values for head height than those with mouths closed. If the relative frequency of open-mouthed versus closed-mouthed specimens (or any other preservation artifact) differed between lineages, this might yield a spurious association of head shape with lineage. Our measurements were made from live specimens with closed mouths, which we infer reduces measurement error. If the lineages differed in a way that would allow separation of them, then our ANCOVA should have revealed this difference.
Along with evidence of extensive population genetic admixture and gene flow between the Atlantic and Gulf Coast lineages (see Folt et al. 2019b), our evaluation of morphological features finds further support for not recognizing D. kolpobasileus as a distinct species. Therefore, we formally propose here Drymarchon kolpobasileus to be placed in the synonymy of Drymarchon couperi .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |