
Mesamphiagrion Kennedy 1920
|
publication ID |
https://doi.org/10.5281/zenodo.182666 |
|
DOI |
https://doi.org/10.5281/zenodo.5691611 |
|
persistent identifier |
https://treatment.plazi.org/id/03ED87D9-FFB6-BE5A-A9D1-A559FCD46723 |
|
treatment provided by |
Plazi |
|
scientific name |
Mesamphiagrion Kennedy 1920 |
| status |
|
Mesamphiagrion Kennedy 1920 View in CoL stat. rev.
Figures 1 View FIGURE 1 b; 2b–c; 3c–f; 4b–e; 7; 15–24; 38–48; 60–84; 98–100; 103; 107–109
Mesamphiagrion Kennedy 1920: 87 View in CoL (diagnosis; designation of Enallagma occultum Ris 1918 View in CoL as type species).
St. Quentin 1960: 52 (key).
Davies 1981: 6 (generic listing with type species listed).
Davies & Tobin 1984: 75 (synonymic list).
De Marmels 1989 (redefinition).
Bridges 1994: III.29 (synonymic list).
Steinmann 1997: 278 (synonymic list).
Tsuda 2000: 39 (synonymic list).
Archaeallagma Kennedy 1920: 87 (diagnosis; designation of Enallagma ovigerum Calvert 1909 as type species). — syn. nov.
Lieftinck 1949: 204 (summary of similarities among Protallagma / Archaeallagma / Oreagrion ).
Davies 1981: 5 (as synonym of Enallagma ).
Davies & Tobin 1984: 65 (synonymic list).
De Marmels 1988: 100 (synonymy with Cyanallagma ).
De Marmels 1989: 250 (discussion of synonymy with Cyanallagma ).
Bridges 1994: III.5 (as synonym of Cyanallagma ).
Steinmann 1997: 245 (synonymic list as valid genus).
Tsuda 2000: 202 (as synonym of Cyanallagma ).
Cyanallagma Kennedy 1920: 87 View in CoL (in part; inclusion of Acanthagrion laterale Selys 1876 View in CoL ).
Davies & Tobin 1984: 66 (in part, synonymic list).
De Marmels 1989: 246–248 (generic characterization).
Bridges 1994: III.13 (in part, synonymic list).
Steinmann 1997: 247 (in part, synonymic list).
De Marmels 1997: 135–156, figs. 1–85 (in part, generic diagnosis, keys, maps, and illustrations for northern species, description of the larva of ' C. ' gaianii ).
Tsuda 2000: 31 (in part, synonymic list).
De Marmels 2007: 96–110, figs. 111–122 (description of larvae of ' C. ' laterale and ' C. ' tamaense ).
Type species: Enallagma occultum Ris 1918 by original designation ( Kennedy 1920: 87).
Other species included: M. demarmelsi ( Cruz 1986) comb. nov., M. dunklei sp. nov., M. ecuatoriale sp. nov., M. gaianii ( De Marmels 1997) comb. nov., M. laterale ( Selys 1876) comb. nov. [syn Argia ternaria Navás 1934 , syn Argia trina Navás 1934 ], M. ovigerum ( Calvert 1909) comb. nov. [ Argia hebdomatica Navás 1934 , syn. nov.], M. risi ( De Marmels 1997) comb. nov., M. tamaense ( De Marmels 1988) comb. nov., and M. tepuianum ( De Marmels 1997) comb. nov..
Specimens examined. M. demarmelsi (1 ɗ). COLOMBIA. Cundinamarca Dept.: 11 km N of Bogotá, 8.iii.1969, P.P. Spangler leg., 1 ɗ (RWG).
M. dunklei (31 ɗ, 11 Ψ). ECUADOR. Napo Prov. See details under species account.
M. ecuatoriale (4 ɗ, 2 Ψ). ECUADOR. Napo Prov. See details under species account.
M. gaianii (2 ɗ). VENEZUELA. Trujillo State: Ancient road Boconó-Trujillo, Páramo La Cristalina, 29.viii.1991, J. De Marmels leg., 2 ɗ paratypes (RWG).
M. laterale View in CoL (26 ɗ, 8 Ψ). VENEZUELA. Mérida State: 17 km E of La Azulita, 26.vii.1989, T.W. Donnelly leg., 3 ɗ (TWD); same data but 1 ɗ (RWG); same data but 3 ɗ (FSCA); Carbonera, 31.xii.1960, J. Rácenis leg., 1 ɗ (FSCA). COLOMBIA. Boyacá Dept.: La Pica, 13.ii / 3.iii.1917, M.A. Carriker leg., 12 ɗ, 3 Ψ (RWG); Santuario de Fauna y Flora de IGUAQUE, near Arcabuco, 1.viii. 1991, J. Delgado leg., 2 Ψ (TWD, RWG); same data but 1.ix.1991, 1 Ψ (TWD); same data but 1.x.1991 (C. Rodrigues), 1 Ψ (TWD); Cauca Dept.: Nueva Granada, M. Mac Lachlan leg., 1 ɗ lectotype (BMNH); same data 2 ɗ paralectotypes (IRSNB). Santander Dept.: La Unión, 1 ɗ holotype of Argia trina Navás 1934 View in CoL (FSCA). Cundinamarca Dept.: Quetame, vii.1912, 1 ɗ syntype of Argia ternaria Navás 1934 View in CoL (UMMZ); Pensilvania, 1913, 1 ɗ syntype of Argia ternaria Navás 1934 View in CoL (NHMP); Choachí, viii.1916, 1 Ψ syntype of Argia ternaria Navás 1934 View in CoL (NHMP).
M. occultum View in CoL (3 ɗ). COLOMBIA. Cundinamarca Dept.: Parque Nacional Chingaza, Quebrada La Playa, 9.xi.2003, O. Realpe & M. Beltrán leg., 1 ɗ (RWG); same data but Churca Ludeck, 20.ix.2003, 2 ɗ (RWG). M. ovigerum View in CoL (3 ɗ). COLOMBIA. Cundinamarca Dept.: Santa Fe de Bogotá, 1863, Lindig leg., 1 ɗ holotype (MCZ); Pensilvania, 1913, 1 ɗ lectotype A. hebdomatica View in CoL (MNHP); Boyacá Dept.: Arcabuco, Santuario de Flora y Fauna de Iguaque, 2400-3600 m, 16.xi.1991, M. Roa leg., 1 ɗ (DRP).
M. tamaense (11 ɗ, 1 Ψ). COLOMBIA. Boyacá Dept.: La Pica, 9-14.ii.1917, M.A. Carriker leg., 11 ɗ, 1 Ψ (RWG).
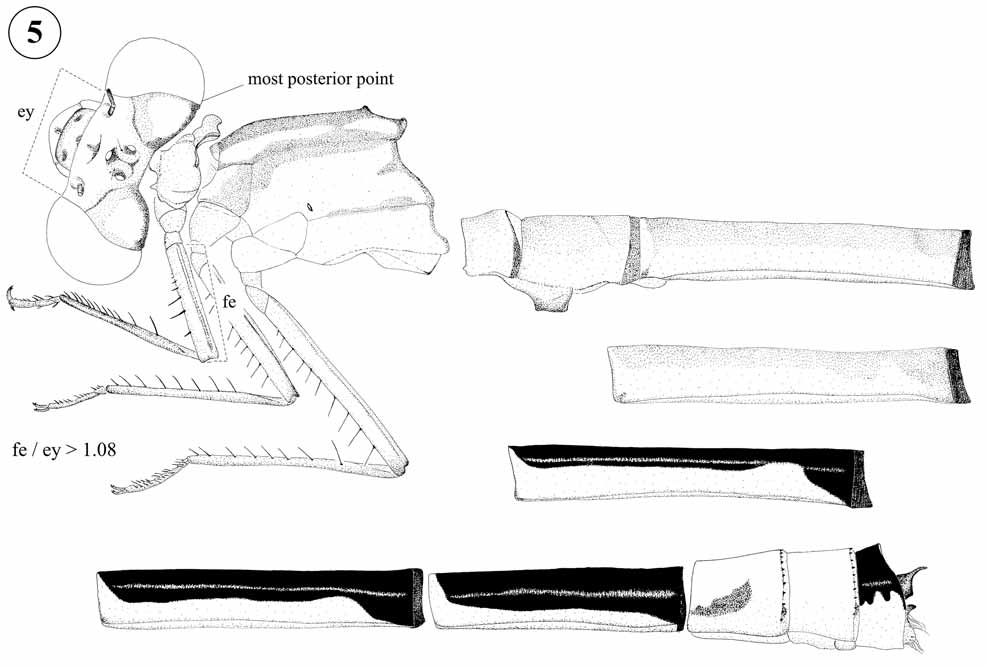
Generic characterization. Head. Color of dorsum dark reddish brown to black with pale blue to olive postocular spots, usually no pale postocular bar but present in some specimens of M. laterale ; rear of head surrounding occipital foramen pale ( Fig. 1 View FIGURE 1 b). Frons rounded, occipital lobes protruding posteriorly so that most posterior point of head is at their level ( Figs. 3 View FIGURE 3 c–e). Thorax. Posterior lobe of prothorax trilobate, usually with medial lobe developed into a caudally projected plate ( Figs. 15 View FIGURES 15 – 24 ; 16a; 17–18; 20a, b–22a, b; 23a; 24a– b) especially in males, but only slightly projected in male M. tepuianum ( Fig. 19 View FIGURES 15 – 24 ) and female M. ecuatoriale ( Fig. 16 View FIGURES 15 – 24 b) or not projected in females of M. dunklei and M. gaianii ( Figs. 22 View FIGURES 15 – 24 c; 23b). Female mesostigmal plates broadly triangular and wide, each with ratio of maximum width/length of less than 0.5; mesanepisternal carinae arising between mesostigmal plates anteriorly to their postero-medial edges ( Figs. 16 View FIGURES 15 – 24 c; 20c–21c; 22d; 23c–24c). Pterothorax (Figs. 2b–c; 3c–e) with dark mid-dorsal and humeral stripes, usually with a dark stripe over metapleural suture (Figs. 2b; 3d–f) that is absent in M. ecuatoriale , M. occultum , M. ovigerum , and M. tepuianum (Figs. 2c–3c); with pale blue antehumeral stripe usually complete (Figs. 2b; 3c–e), but interrupted distally in M. tepuianum and some M. demarmelsi (Fig. 2c). Legs short with femur 1 usually shorter than distance between eyes at level of antennifer ( Figs. 3 View FIGURE 3 d–e; ratio = 1.03), tibial spurs shorter to slightly longer than distance between them ( Figs. 3 View FIGURE 3 d–e); pretarsal claw with well developed supplementary tooth. Wings hyaline to smoky in some tenerals ( i.e. in M. laterale and M. tamaense ), CuP reaching CuPAA proximal to hind margin of wing for a distance as long as CuP or shorter, vein descending from quadrangle not forming a straight line to wing margin ( Fig. 7 View FIGURE 7 ).
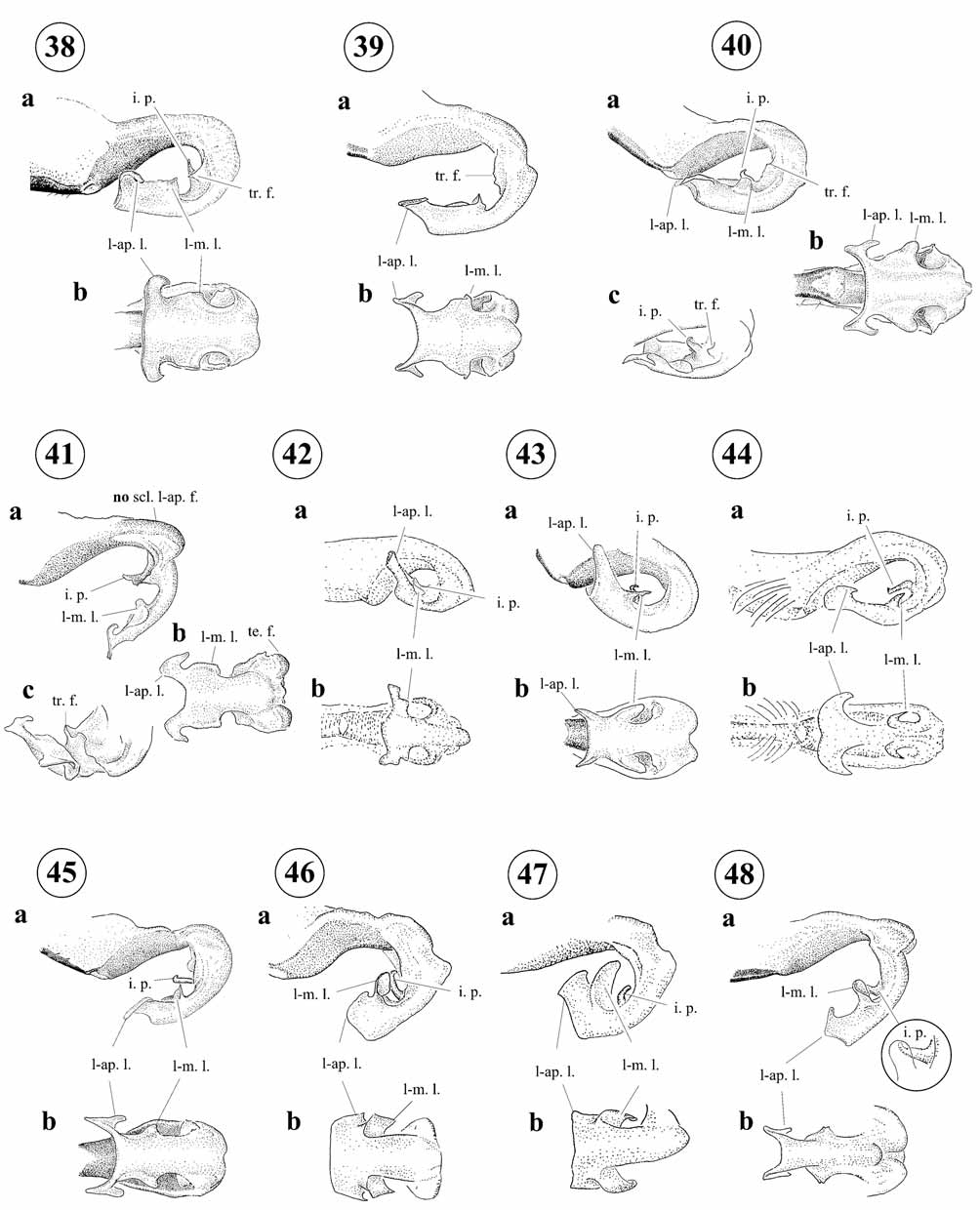
Abdomen ( Fig. 4 View FIGURE 4 b–e). Black with blue spots on base and distal segments (S6–10 or less), and reddishorange (on S1–3) on not fully mature specimens ( Figs. 108 –109); relatively short, with a ratio of 3.39–4.28 to length of head plus thorax, less in M. ecuatoriale (4.2 to 4.85). Genital ligula with distal segment lacking inner fold; with paired latero-apical lobes and paired latero-medial lobes ( Figs. 38–48 View FIGURES 38 – 48 ); with two ental membranous transverse folds distal to flexure, one between medio-lateral lobes and the second between medio-lateral lobes and flexure, this latter fold projected into a medial membranous process that may be folded and hidden in lateral view; second segment lacking wide latero-apical folds with sclerotized margins ( Figs. 38–48 View FIGURES 38 – 48 ). Posterodorsal margin of male S10 with a 'u'-shaped cleft margined by a pair of tubercles ( Figs. 60 View FIGURES 60 – 69 –70; 71a; 77b–c; 78b–79b; 80–84). Male cercus sometimes with a pale spot on its posterior surface but lacking differentiated scalariform-like cuticle in that area. Male cercus structure complex with four processes that usually can be seen simultaneously only in medio-posterior view (Figs. 70b; 71–76; 77a–79a): a dorsal process that may be apical, long, and hooked (as in M. demarmelsi , M. dunklei , M. gaianii , M. laterale , and M. tamaense ) or subapical and short (as in M. ecuatoriale , M. occultum , M. ovigerum , M. risi , and M. tepuianum ); two ventro-apical processes represented by short lobes each of which may be rounded (as in M. ecuatoriale , M. occultum , M. ovigerum , and M. risi ) or bluntly pointed (as in M. demarmelsi , M. tepuianum , M. dunklei , M. gaianii , M. laterale , and M. tamaense ); and a broadly triangular ventro-basal process that is directed antero-medially and is visible only in medio-posterior view or in dorso-external view (as in M. demarmelsi , Fig. 65 View FIGURES 60 – 69 b). Male paraproct with a dorsal branch ending on a sclerotized tip ( Figs. 60–69 View FIGURES 60 – 69 ; 70a–71a; 72; 76; 77b–c; 78b–79b).
Female with vulvar spine on S8; ovipositor slightly shorter to slightly longer than S10, not reaching tips of cerci ( Figs. 4 View FIGURE 4 c, e).
Generic diagnosis. Male cercus of Mesamphiagrion is unique among all New World Coenagrionidae by its four processes: one dorsal, two short ventro-apical and one ventro-basal (Figs. 70b; 71–76; 77a–79a). As mentioned under Cyanallagma , the combination of a rounded frons, presence of pale postocular spots, a trilobate prothoracic posterior lobe, striped pterothorax and male cerci provided with some kind of processes is shared among New World Coenagrionidae not only with Cyanallagma and Oreiallagma but also with Apanisagrion , Chrysobasis , Hesperagrion , Homeoura , some Ischnura species, Leptobasis , and Telagrion . Mesamphiagrion differs from Cyanallagma by having the rear of head surrounding occipital foramen pale ( Fig. 1 View FIGURE 1 b) and by the male cercus having a dorsal process ( Figs. 60–84 View FIGURES 60 – 69 View FIGURES 80 – 84 ). It can be distinguished from all remaining mentioned genera except Hesperagrion and some Ischnura species by its most posterior point of head located at level of postocular lobes ( Figs. 3 View FIGURE 3 c–e) rather than at level of eyes (as in Fig. 5 View FIGURE 5 ).
Mesamphiagrion is separated from Hesperagrion by the male postero-dorsal margin of S10 with a 'u'- shaped cleft margined by a pair of tubercles ( Figs. 60–84 View FIGURES 60 – 69 View FIGURES 80 – 84 ), genital ligula lacking a pair of large lateral chitinized spines ( Figs. 38–48 View FIGURES 38 – 48 ), and female mesanepisterna with well developed carinae forming distinct ridges ( Figs. 16 View FIGURES 15 – 24 c; 20c; 22d; 23c–24c). Mesamphiagrion differs from Ischnura by the presence of a ventro-basal process in male cercus ( Figs. 60–84 View FIGURES 60 – 69 View FIGURES 80 – 84 ), genital ligula distal segment lacking an inner fold, presence of paired latero-apical and latero-medial lobes and distal corners not projected into flagellae ( Figs. 38–48 View FIGURES 38 – 48 ), and female mesanepisterna with well developed carinae forming distinct ridges ( Figs. 16 View FIGURES 15 – 24 c; 20c; 22d; 23c–24c).
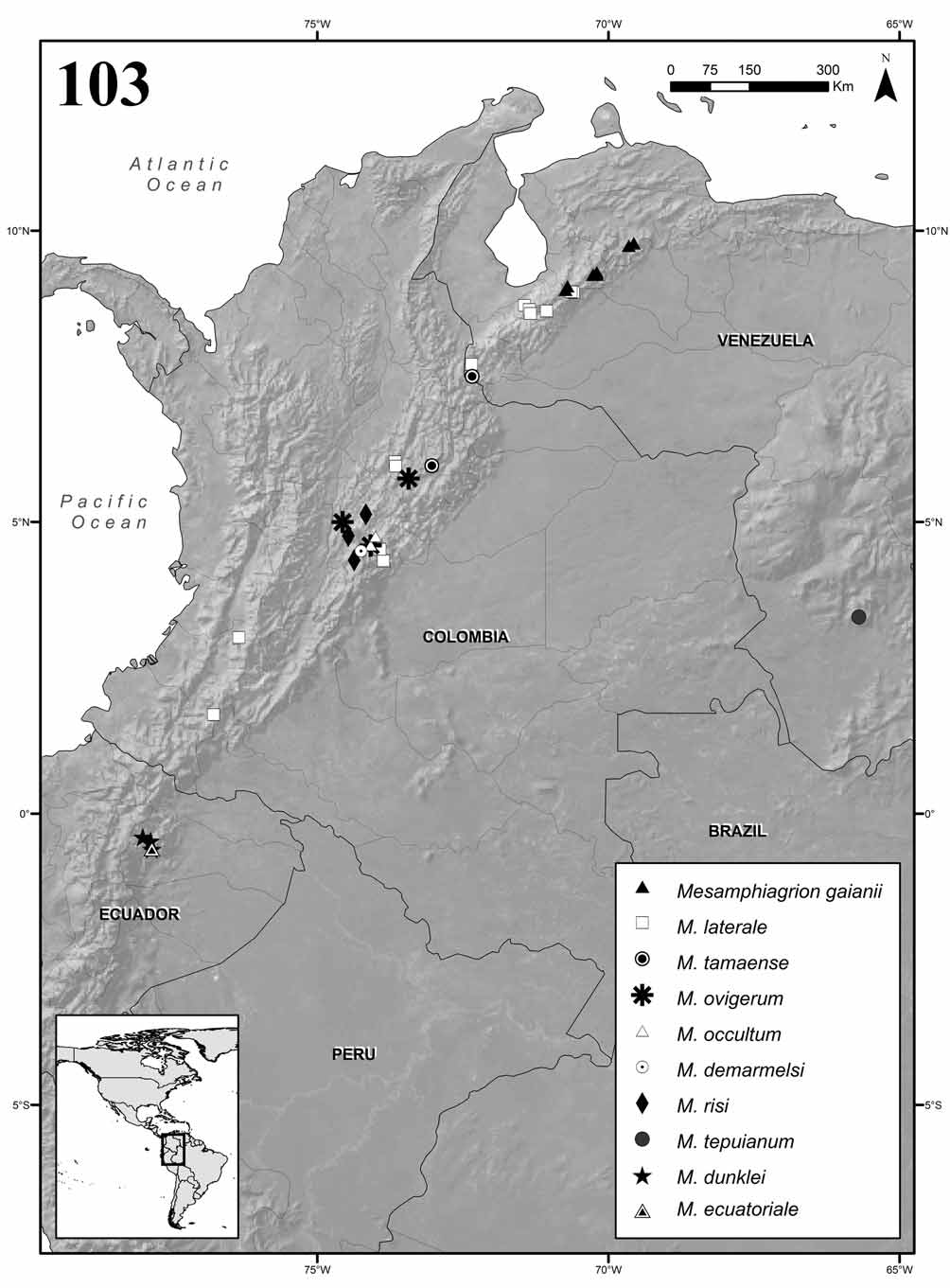
Distribution. Northwestern South America from Ecuador to Venezuela along the Andes and in the Tepuis, from 650 to 4000 m above sea level ( Fig. 103 View FIGURE 103 ).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Mesamphiagrion Kennedy 1920
| Ellenrieder, Natalia Von & Garrison, Rosser W. 2008 |
Mesamphiagrion
| Kennedy 1920: 87 |
Cyanallagma
| Kennedy 1920: 87 |