
Polacanthoderes Sørensen, 2008a
|
publication ID |
https://doi.org/ 10.1016/j.jcz.2022.09.003 |
|
publication LSID |
lsid:zoobank.org:pub:A6CE656D-6409-4DA4-9A2A-A0AA63490C81 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE8791-857B-9755-9A7E-1253FAEFFAB8 |
|
treatment provided by |
Felipe |
|
scientific name |
Polacanthoderes Sørensen, 2008a |
| status |
|
Genus Polacanthoderes Sørensen, 2008a View in CoL .
3.1.1. Emended diagnosis for Polacanthoderes
Cuticle of first trunk segment forms complete ring; cuticle of segments 2–11 with midventral and lateroventral articulations resulting in one tergal and two sternal plates. Cuticular hairs and perforation sites absent on all segments. Primary pectinate fringe of ventral side on segment 1 with very large and conspicuous fringe tips; primary pectinate fringes of other areas on segment 1 and those on other segments with very small fringe tips. Middorsal acicular spines on segments 4–8; lateroventral acicular spines on segments 6–9; additional small acicular spines present in various positions on segments 4–10, at least in subdorsal position on segments 6 and 7, laterodorsal to midlateral position on segments 5–9, sublateral or lateral accessory position on segment 7, lateral accessory position on segments 6, 8, and 9, ventrolateral position on segments 8–10, and ventromedial position on segments 4–7. Tubes in midlateral position on segment 4; lateroventral position on segment 5; laterodorsal position on segment 10. Ventromedial setae present on segment 11. Females with ventromedial papillae on segment 6 and ventrolateral papillae on segment 7.
3.1.2. Polacanthoderes martinezi S ø rensen, 2008a
( Figs. 2–4 View Fig View Fig View Fig ; Table 4)
Polacanthoderes martinezi Sørensen 2008a: 230 View in CoL e3–230e11, Figs. 1–5 View Fig View Fig View Fig View Fig View Fig , Tables 1 and 2
3.1.2.1. Emended diagnosis. Polacanthoderes with small lateral accessory acicular spines on segment 7; lateroventral acicular spines on segments 8 and 9 similar in width to those on segments 6 and 7.
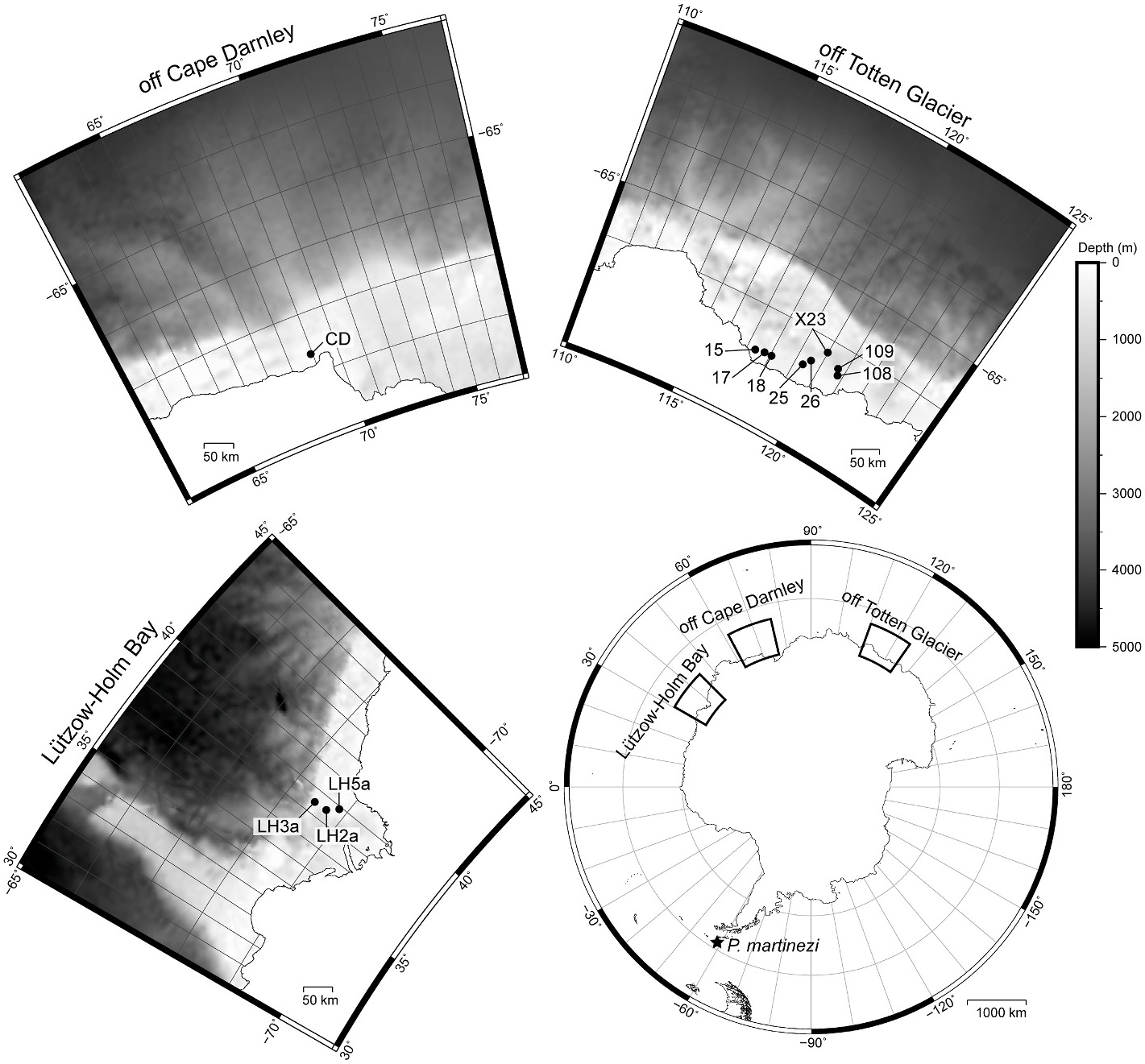
3.1.2.2. Material examined. Holotype: Adult female, collected 14 February 2002 at north of South Shetland Islands (61 ◦ 23.73 ′ S, 58 ◦ 50.26 ′ W, 2274 m depth), mounted in Fluoromount G ® ( ZMB 11237 View Materials ) GoogleMaps . Allotype: Adult male, collected 14 February 2002 at north of South Shetland Islands (61 ◦ 24.14 ′ S, 58 ◦ 51.15 ′ W, 2290 m depth), mounted in Fluoromount G ® ( ZMB 11238 View Materials a) GoogleMaps . Paratypes: Three adult females and one adult male, collected at the same time as holotype or allotype, mounted in Fluoromount G ® ( ZMB 11238 View Materials b–d and 11238f). Non-types : three females and one male, collected at the same localities, mounted on aluminium stubs for SEM.
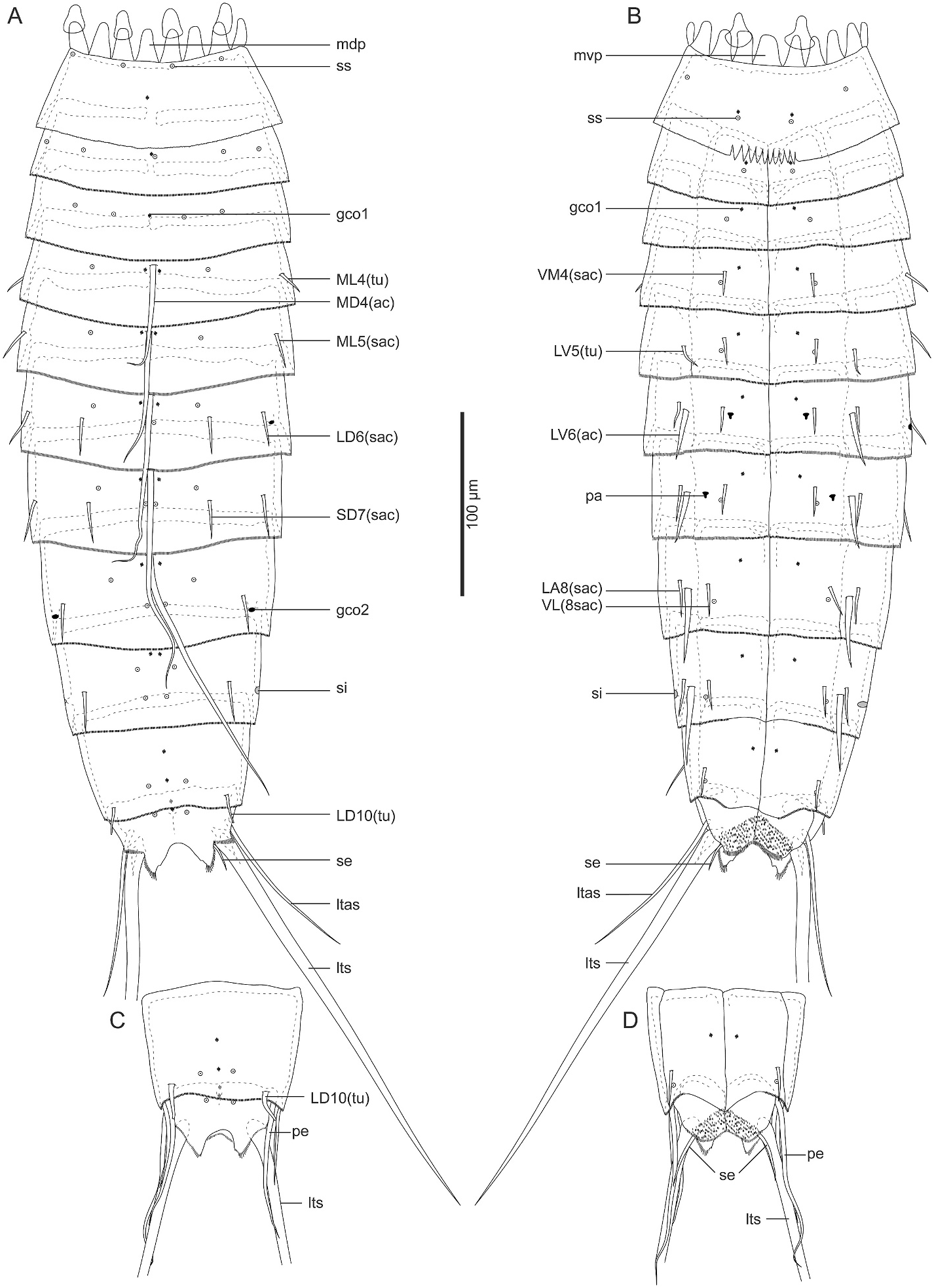
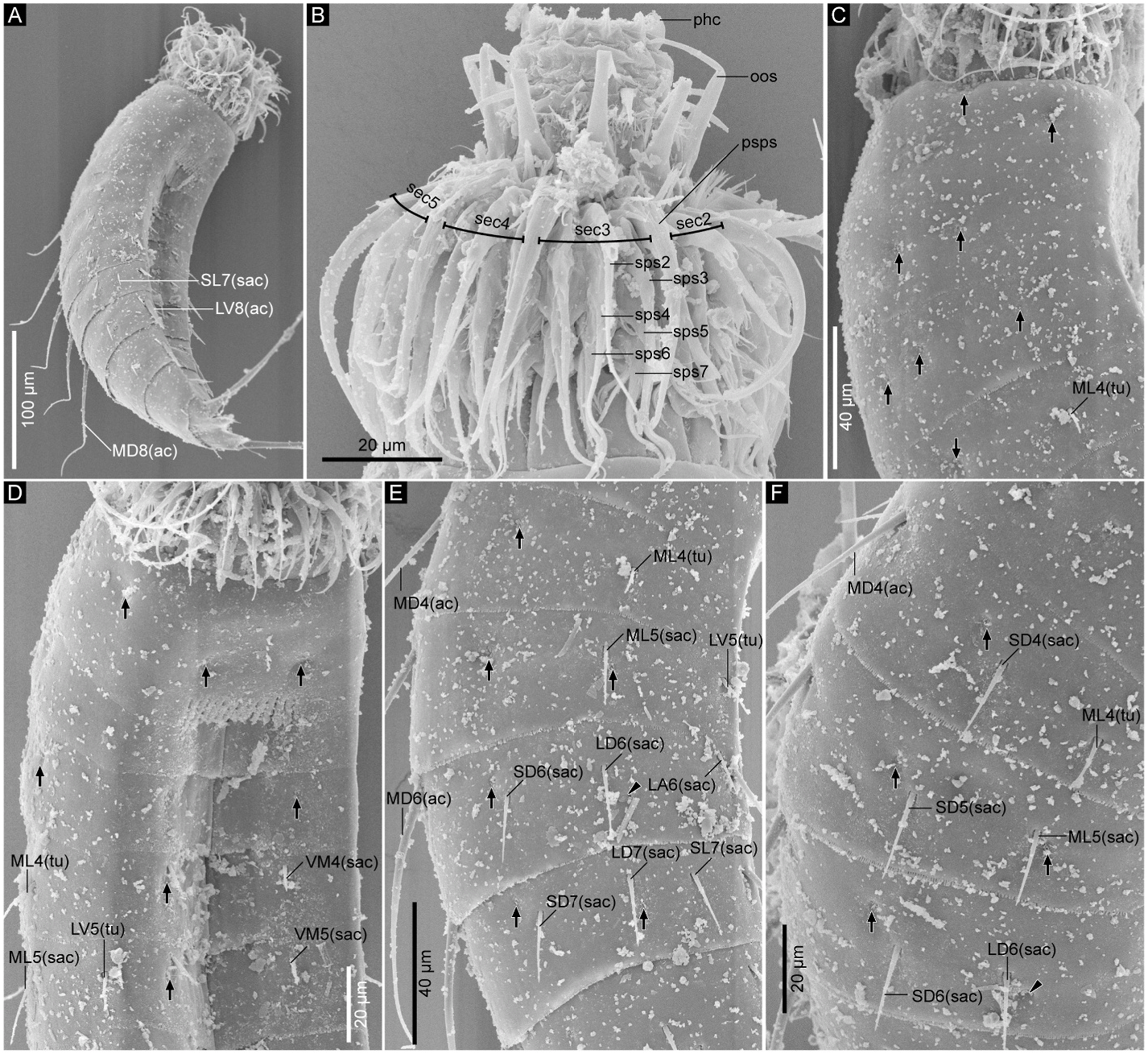
3.1.2.3. Redescription and remarks. Adult with head, neck, and eleven trunk segments ( Figs. 2A–D View Fig and 3A–F View Fig ). See Table 1 in Sørensen (2008a) for measurements. Cuticular structures are summarized in Table 4. The distribution of cuticular structures generally follows the original description in Sørensen (2008a) ( Figs. 2A–D View Fig , 3A–F View Fig , 4A–D View Fig ), hence the following notes provide additional morphological data not mentioned in the original description.
Our observation confirmed the presence of all sensory spots and glandular cell outlets mentioned in Sørensen (2008a). In addition to the original description, the following previously unnoticed features were observed: the presence of subdorsal and laterodorsal sensory spots as well as ventromedial type-1 glandular cell outlets on segment 1 ( Figs. 2A and B View Fig , and 3A–C View Fig ); ventromedial sensory spots and middorsal type-1 glandular cell outlet on segment 2 ( Fig. 2A and B View Fig ); ventromedial sensory spots on segments 3 and 4 ( Figs. 2B View Fig and 4A View Fig ); paradorsal sensory spots on segments 6–8 ( Fig. 2A View Fig ); two pairs of subdorsal sensory spots on segment 9 ( Figs. 2A View Fig and 3D View Fig ); and subdorsal sensory spots on segments 10 and 11 ( Fig. 2A, C View Fig , 3D View Fig ). Segment 9 also possesses small-rounded sieve plates in the sublateral position ( Fig. 2A and B View Fig ). Females with ventromedial papillae on segment 6 and ventrolateral papillae on segment 7 ( Figs. 2B View Fig , 3E View Fig and 4C, D View Fig compared to Fig. 3F View Fig ). We also confirmed the presence of long setae on segment 11 in both females and males ( Fig. 2A–D View Fig ).
Among the tubes and acicular spines on the trunk segments, the midlateral ones on segment 4 were referred to as “spines” in the original description. However, our observation confirmed they are thinner than the following small acicular spines and most likely not spines but tubes ( Figs. 2A View Fig , 3C View Fig and 4A View Fig ). The original description also mentioned the variable occurrence of some spines on segments 8 and 9. We additionally found the presence of subdorsal tubes on segment 2 only in one specimen (ZMB 11238a) ( Fig. 3C View Fig ); subdorsal small acicular spines or tubes on segment 5 in two specimens (ZMB 11238a and a male SEM specimen); and the absence of midlateral tubes on segment 4 in one specimen (ZMB 11238d).
3.1.3. Polacanthoderes shiraseae Yamasaki , sp. nov
Zoobank ID: urn:lsid:zoobank.org:act:B91EF876-5D41-4ABE-91C5-E150CF92BC31
( Figs. 5–9 View Fig View Fig View Fig View Fig View Fig ; Tables 5 and 6).
3.1.3.1. Diagnosis. Polacanthoderes with sublateral small acicular spines on segment 7; lateroventral acicular spines on segments 8 and 9 being conspicuously thicker than those on segments 6 and 7.
3.1.3.2. Etymology. The specific name “ shiraseae ” is a noun in the genitive case, named after the Japanese icebreaker Shirase, which was used for collecting the sediment sample containing Polacanthoderes shiraseae sp. nov.
3.1.3.3. Material examined. Holotype: Adult female (catalogue No. ICHUM 8348 View Materials ), collected 19 December 2019 from off Totten Glacier, Antarctica (66 ◦ 47 ′ 31 ′′ S, 117 ◦ 21 ′ 20 ′′ E, 691 m depth), mounted in Fluoromount G ® on H–S slides GoogleMaps . Paratypes: 30 adult males and 36 adult females (catalogue no. ICHUM 8349–8404 View Materials and NHMD 1174924–1174933 ), detailed sampling data shown in Table 1, all individually mounted in Fluoromount G ® on H–S slides. Non-type : nine adult males and eight adult females, detailed sampling data shown in Table 1, mounted on aluminium stubs for SEM.
3.1.3.4. Type locality. Off Totten Glacier , Antarctica (66 ◦ 47 ′ 31.20 ′′ S, 117 ◦ 21 ′ 19.80 ′′ E), 691 m depth GoogleMaps .
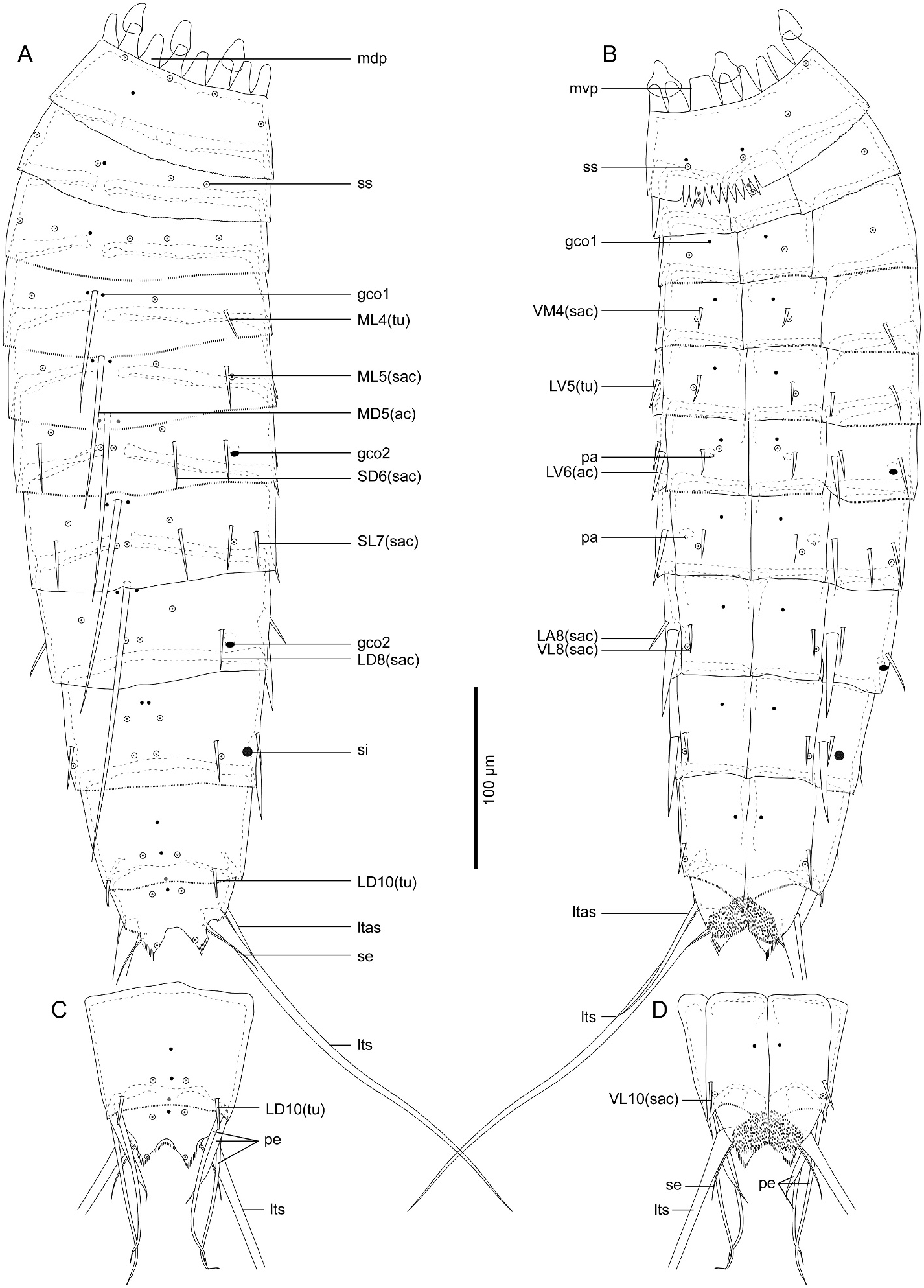
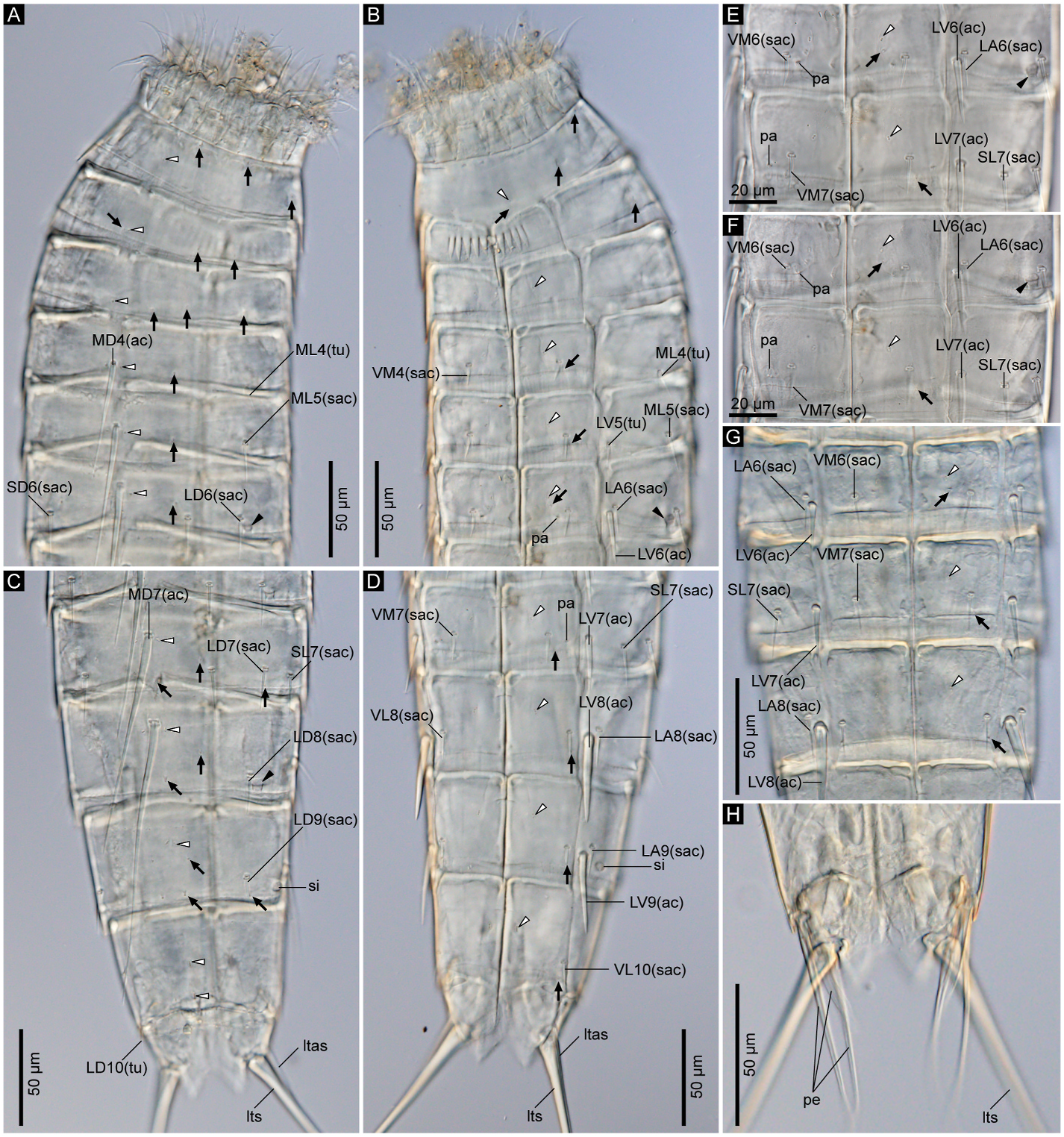
3.1.3.5. Description. Adult with head, neck, and eleven trunk segments ( Figs. 5 View Fig , 6A–D View Fig , and 7A View Fig ). See Table 5 for measurements. Table 6 indicates the positions of cuticular structures.
Head consists of retractable mouth cone and introvert ( Fig. 7B View Fig ). Distal part of mouth cone with pharynx crown, three rings of inner oral styles, and one ring of nine outer oral styles. Each of ring-03 and ring-02 bears five inner oral styles, whereas details of inner oral styles in ring-01 not observable. Outer oral styles alternating in length: slightly longer in odd sectors than in even sectors. Outer oral style in sector 6 missing. Introvert with seven rings of spinoscalids and one ring of trichoscalids. Ring 01 with ten primary spinoscalids composed of one basal sheath and one distal end piece. Each basal sheath with two overlapping fringes: proximal fringe composed of several short projections between two long and slightly thicker lateral ones; distal fringe with several long projections. Distal end pieces of primary spinoscalids longest within all spinoscalids. Rings 02 and 04 with 10 spinoscalids; rings 03 and 05 with 20 spinoscalids. Each spinoscalid of rings 02–05 with basal sheath and end piece. Rings 06 and 07 not examined in detail, but with at least seven and four spinoscalids, respectively. Six trichoscalids covered with long hairs on entire surface and each attached to trichoscalid plates. Number and arrangement of inner oral styles, outer oral styles, and spinoscalids summarized in Fig. 8 View Fig .
Neck with 16 placids ( Fig. 5A and B View Fig , 6A, B View Fig , and 8 View Fig ). Midventral placid wider than others, and others similar in width.
Trunk with eleven segments; segment 1 consists of complete cuticular ring; segments 2–11 consist of one tergal and two sternal plates ( Figs. 5 View Fig and 6A–D View Fig ). Thickened cuticle forms pachycyclus at anterior margin and along midventral and tergosternal articulations of segments 2–10 ( Fig. 6 View Fig ). Cuticular hairs and perforation sites absent on all segments ( Figs. 6 View Fig , 7 View Fig and 9 View Fig ).
Segment 1 ( Fig. 5A and B View Fig , 6A, B View Fig , and 7C, D View Fig ) with type-1 glandular cell outlet in middorsal position. Additional pair of type-1 glandular cell outlets in ventromedial position. Sensory spots present subdorsally, laterodorsally, sublaterally, and ventromedially. Posterior edge of this and following nine segments with primary pectinate fringe. Fringe tips of primary pectinate fringe in ventromedial area on segment 1 conspicuously large, whereas those in other areas and on other segments small and recognizable only in SEM.
Segment 2 ( Fig. 5A and B View Fig , 6A, B View Fig , and 7C, D View Fig ) with type-1 glandular cell outlets in middorsal and ventromedial positions. Two pairs of sensory spots present laterodorsally. Additional sensory spots present middorsally and ventromedially.
Segment 3 ( Fig. 5A and B View Fig , 6A, B View Fig , and 7C, D View Fig ) with middorsal and ventromedial type-1 glandular cell outlets. Sensory spots present in subdorsal, laterodorsal, midlateral, and ventromedial positions.
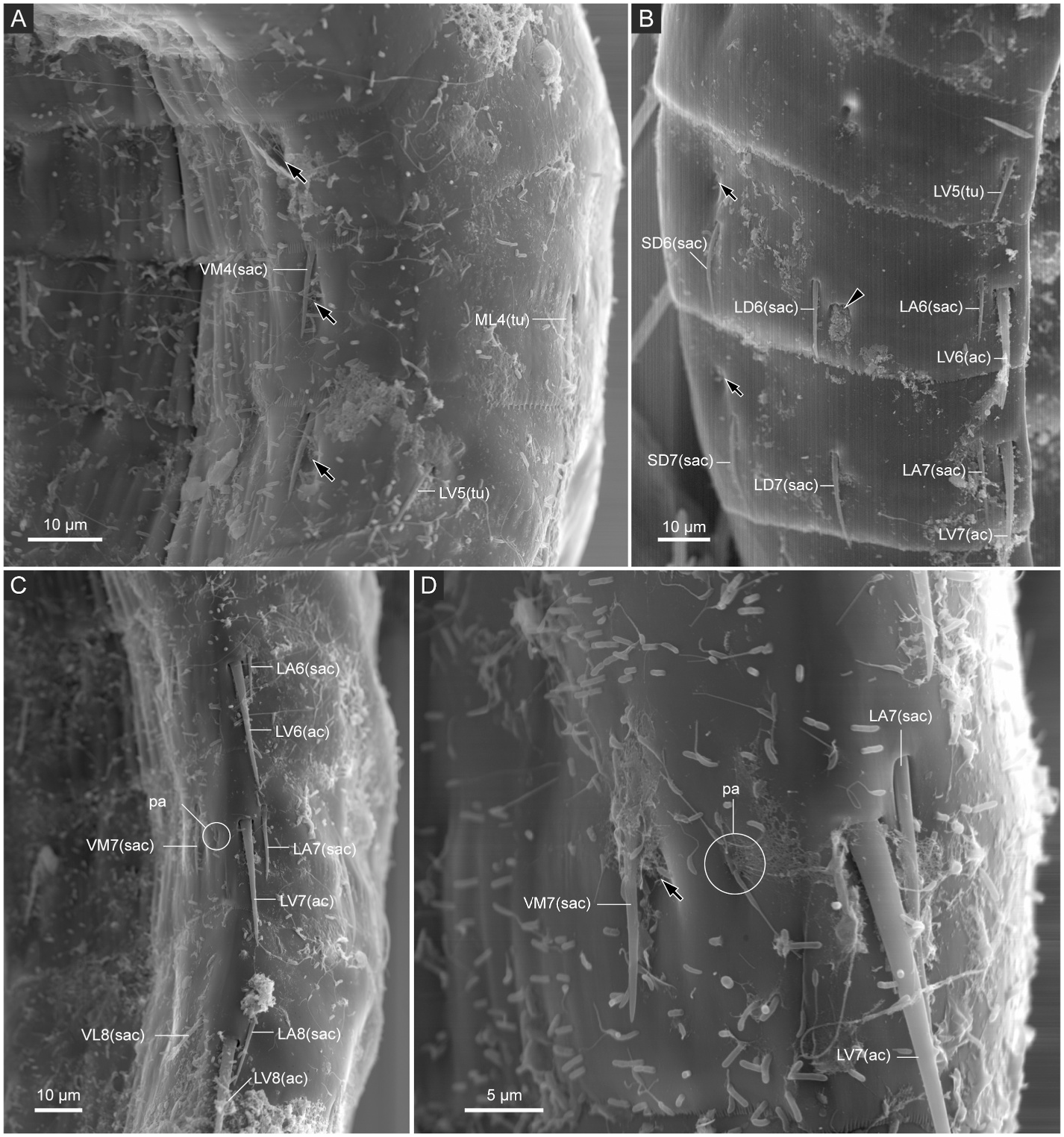
Segment 4 ( Fig. 5A and B View Fig , 6A, B View Fig , and 7C–F View Fig ) with middorsal acicular spine. Small acicular spines present ventromedially and tubes present midlaterally. Nineteen out of 84 examined specimens showed irregular spine patterns on this segment: 17 specimens with additional pair of subdorsal small acicular spines (e.g., compare Fig. 7E and F View Fig ); one specimen with additional midlateral small acicular spine/tube only on left side; one specimen lacks midlateral tubes. Type-1 glandular cell outlets present in paradorsal and ventromedial positions. Sensory spots present subdorsally and ventromedially.
Segment 5 ( Fig. 5A and B View Fig , 6A, B View Fig , 7D–F View Fig , and 9A View Fig ) with middorsal acicular spine. Small acicular spines present in midlateral and ventromedial positions, and tubes present in lateroventral position. Twentyfour out of 84 examined specimens showed irregular spine patterns on this segment: 23 specimens possess additional pair of subdorsal small acicular spines (e.g., compare Fig. 7E and F View Fig ); one specimen with an additional subdorsal small acicular spine only on left side. Type-1 glandular cell outlets present in paradorsal and ventromedial positions. Sensory spots present subdorsally, midlaterally, and ventromedially.
Segment 6 ( Fig. 5A and B View Fig , 6A, B, E–G View Fig , 7E, F View Fig , and 9A, B View Fig ) with middorsal and lateroventral acicular spines. Small acicular spines present in subdorsal, laterodorsal, lateral accessory, and ventromedial positions. Three out of 84 examined specimens showed irregular spine patterns on this segment: lateral accessory small acicular spines moved to sublateral position in 2 specimens; one specimen with an additional sublateral small acicular spine only on right side. Type-1 glandular cell outlets present in paradorsal and ventromedial positions. Type-2 glandular cell outlets present in midlateral position. Sensory spots present paradorsal, subdorsal, and ventromedial positions. Ventromedial papillae present in females.
Segment 7 ( Fig. 5A and B View Fig , 6C–G View Fig , 7A, E View Fig , and 9A–C View Fig ) with middorsal and lateroventral acicular spines. Small acicular spines present subdorsally, laterodorsally, sublaterally, and ventromedially. Eight out of 84 examined specimens showed irregular spine pattern on this segment: sublateral small acicular spines moved to lateral accessory position in four specimens, of which one specimen also lacks lateroventral acicular spine on left side; one specimen with a non-paired sublateral small acicular spines on right side and paired lateral accessory small acicular spines; one specimen lacks one laterodorsal small acicular spine on right side; two specimens with one additional lateral accessory small acicular spines only on one side. Type-1 glandular cell outlets present in paradorsal and ventromedial positions. Sensory spots present paradorsally, subdorsally, midlaterally, and ventromedially. Papillae present in ventrolateral position in females.
Segment 8 ( Fig. 5A and B View Fig , 6C, D, G View Fig , 7A View Fig , and 9B, C, E View Fig ) with middorsal and lateroventral acicular spines. Lateroventral acicular spines conspicuously thicker and longer than those on previous two segments. Small acicular spines present laterodorsal, lateral accessory, and ventrolateral positions. One out of 84 examined specimens with one subdorsal small acicular spine only on right side. Type-1 glandular cell outlets present paradorsally and ventromedially. Type-2 glandular cell outlets present in midlateral position. Sensory spots present in paradorsal, subdorsal, midlateral, and ventrolateral positions.
Segment 9 ( Fig. 5A and B View Fig , 6C, D View Fig , and 9C–F View Fig ) with lateroventral acicular spines, which are thicker than those on segments 6 and 7, but slightly shorter than those on segment 8. Small acicular spines present in laterodorsal, lateral accessory, and ventrolateral positions. Type-1 glandular cell outlets present paradorsally and ventromedially. Two pairs of sensory spots present in subdorsal position, and pair of sensory spots present in laterodorsal and ventrolateral positions. Small rounded sieve plate present sublaterally.
Segment 10 ( Figs. 5 View Fig and 6C, D View Fig , and 9E, F View Fig ) with laterodorsal tubes and ventrolateral small acicular spines. Two type-1 glandular cell outlets present in middorsal position, and one pair in ventromedial position. Sensory spots present subdorsally and ventrolaterally.
Segment 11 ( Figs. 5 View Fig and 6C, D, H View Fig , and 9E, F View Fig ) with lateral terminal spines. Males with three pairs of penile spines: most dorsal and most ventral penile spines quite slender and long, whereas middle penile spines much thicker and short. Females with lateral terminal accessory spines. Two type-1 glandular cell outlets present middorsally, and sensory spots present subdorsally. Surface of ventromedial area of sternal plates furry. Sternal plates triangle. Posterior edge of tergal plate protruding subdorsally forming pointed tergal extensions. Thin and long setae present next to tergal extensions in both males and females.
3.1.3.6. Remarks. Polacanthoderes shiraseae sp. nov. is similar to P. martinezi in many characters: the presence of (1) very strong and conspicuous pectinate fringe in the ventral side on segment 1, (2) middorsal acicular spines on segments 4–8, (3) lateroventral acicular spines on segments 6–9, (4) small acicular spines present at least in subdorsal on segments 6 and 7, laterodorsal on segments 5–9, lateral accessory on segments 6, 8, and 9, ventrolateral on segments 8–10, and ventromedial on segments 4–7, (5) tubes in midlateral on segment 4, lateroventral on segment 5, and laterodorsal on segment 10, (6) type-1 glandular cell outlets in middorsal on segments 1–3, 10, and 11, paradorsal on segments 4–9, ventromedial on segments 1–10, (7) type-2 glandular cell outlets in midlateral on segments 6 and 8, and (8) ventromedial setae on segment 11, and (9) females with ventromedial papillae on segment 6 and ventrolateral papillae on segment 7 ( Sørensen, 2008a). Furthermore, both of the two species sometimes possess irregular small acicular spines, especially on segments 4–8.
Although the two Polacanthoderes species share many characters, they can be distinguished clearly in the shapes of lateroventral acicular spines: the lateroventral acicular spines on segments 8 and 9 are conspicuously thicker than those on segments 6 and 7 in P. shiraseae sp. nov., whereas those on segments 6–9 are all similar in width in P. martinezi . In addition, P. shiraseae sp. nov. can be distinguished from P. martinezi in the position of the small acicular spines on segment 7: the spines are in the sublateral position in the former, whereas they are in the lateral accessory position in the latter species. Although the spine position may be irregularly moved from sublateral to lateral accessory position in P. shiraseae sp. nov., such a mutation seldom occurs (less than 10% of the examined specimens of P. shiraseae sp. nov. have small acicular spines in the lateral accessory position in segment 7). Furthermore, even if the spine position on segment 7 is inapplicable for some specimens, the width of the lateroventral acicular spines on segments 8 and 9 can be used for the identification of the species.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Polacanthoderes Sørensen, 2008a
| Yamasaki, Hiroshi, Fujimoto, Shinta, Tanaka, Hayato, Shimada, Daisuke, Ito, Masato, Tokuda, Yuki & Tsujimoto, Megumu 2022 |
Polacanthoderes martinezi Sørensen 2008a: 230
| Sorensen, M. V. 2008: 230 |