
Iridoteuthis merlini, Reid, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5005.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:A06EDB00-575C-4F6A-B47F-A9CFB4CF50B4 |
|
persistent identifier |
https://treatment.plazi.org/id/03F06B54-317D-FFB9-198A-FB363E3150A0 |
|
treatment provided by |
Plazi |
|
scientific name |
Iridoteuthis merlini |
| status |
sp. nov. |
Iridoteuthis merlini View in CoL new species
( Figures 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 , 11 View FIGURE 11 ; Tables 3, 4; Appendices 3, 4) https://zoobank.org/ urn:lsid:zoobank.org:pub:A06EDB00-575C-4F6A-B47F-A9CFB4CF50B4
Iridoteuthis sp. Lu & Dunning 1998: 513, Figure 13.7 A, F.
Type material. Holotype: New Zealand: ♂ (16.5 mm ML), W of New Plymouth , 39°2.27´S, 172°13.5´E, 530 m (over 710–732 m), coll. RV ‘ James Cook’, Stn J 07/59/80, 12 Apr. 1980 ( NMNZ M.67223) GoogleMaps . Paratypes. New Zealand: ♂ (15.0 mm ML) , ♀ (11.2 mm ML), SE of Mercury Islands , 36°46.8´S, 176°18.5´E, 114 m (over 620 m), coll. RV ‘ James Cook’, Stn J 11/38/76, 31 Jul. 1976 ( NMNZ M.91591) GoogleMaps ; ♀ (10.9 mm ML), 37°13´37.79´´S, 176°19´20.98´´E to 37°10´51.60´´S, 176°17´45.62´´E, 358–376 m, coll. Minister of Fisheries , NIWA, 21 Jan.1998 ( NIWA 50749 View Materials ) GoogleMaps ; 2♂ (14.1, 16.8 mm ML), ♀, 9.4 mm ML, 1 juv. 5.0 mm ML, E of Mayor Island , 37°16.1´S, 176°19.4´E, 243 m (over 402 m), coll. RV ‘ James Cook’, Stn J 12, 15 Aug. 1975 ( NMNZ M.287523) GoogleMaps ; ♂ (14.3 mm ML), 4♀ (10.8–13.6 mm ML), E of Mayor Island , 37°17.3´S, 176°18´E, 91–223 m, coll. RV ‘ James Cook’, Stn J 07/24/75, 6 May 1975 ( NMNZ M.287529) GoogleMaps ; ♂ (14.7 mm ML), ESE of White Island , 37°33.8´S, 177°16.6´E, 360 m, coll. RV ‘ James Cook’, Stn J 11/89/76, 3 Aug. 1976 ( NMNZ M.287527) GoogleMaps ; 2♂ (12.5, 14.1 mm ML), 3♀ (10.7–13.1 mm ML), W of New Plymouth , 39°2.27´S, 172°13.5´E, 530 m (over 710–732 m), coll. RV ‘ James Cook’, Stn J 07/59/80, 12 Apr. 1980 ( NMNZ M.330463) GoogleMaps ; 2♀ [former paratypes S. maoria ] (11.8, 12.0 mm ML), North Bay , Bay of Plenty, 238 m ( NMNZ M.11178) ; 9♂ (8.5–11.3 mm ML), 3♀ (8.5–9.2 mm ML), off White Island , 37°33.7´S, 177°14.90´E, 420–840 m, coll. RV ‘ James Cook’, Stn J 07/50/75, 8 May 1975 ( NMNZ M.329181) GoogleMaps .
Other Material. New Zealand: 3♀ (9.5–13.6 mm ML), 36°38´46.79´´S, 176°10´58.19´´E, 288 m, coll. Min- ister of Fisheries, NIWA, 24 Jan. 1988 ( NIWA 84790 View Materials ) GoogleMaps . Australia: New South Wales: 2♂ (16.1, 12.7 mm ML), off Port Kembla , 34°28´S, 159°29´E, 0–229 m (bottom 1418 m), coll. J. Paxton and K.J. Graham, 22–24 Jul. 1974 ( AM C.451294) GoogleMaps ; 13♂ (11.7–16. 9 mm ML), ♀ (13.3 mm ML), Off Jervis Bay , 35°01´S – 34°58´S, 151°07´E – 150°08´E, 457 m, coll. K.J. Graham, 4 Dec. 1980 ( AM C.451293) GoogleMaps ; ♀ (13.2 mm ML) off Jervis Bay , 35°01´S – 35°07´S, 151°05´E – 151°02´E, 0–256 m (bottom 329–347 m), coll. K.J. Graham, 9 May 1978 ( AM C.451295) GoogleMaps ; 2♂ (13.1, 11.3 mm ML), ♀ (14.2 mm ML), off Ulladulla , 35°28´S, 150°47´E, 0–300 m, (bottom 350 m), coll. J. Paxton and S. Lar- son, 27 Oct. 1977 ( AM C.451286). Tasmania GoogleMaps : ♀ (8.0 mm ML), 42°48´31.68´´S, 148°28´45.84´´E, 0–1000 m, coll. RV ‘ Investigator’, 19 Apr. 2015 ( AM C.487262) GoogleMaps . 1 ♂ (9.4 mm ML), Southern Ocean , 42°49´43.68´´S, 148°30´E, 0–400 m, coll. RV Investigator Trial Voyage, 18 Apr. 2015 ( AM C.487264) . ♀ (10.8 mm ML), 42°48´31.68´´S, 148°28´45.84´´, 0–1000 m, coll. RV ‘ Investigator’, 19 Apr. 2015 ( AM C.487263) .
Diagnosis. Male dorsal arms 1 swollen, laterally ridged; swelling most pronounced on right dorsal arm; suckers uniform in size. Arms 4 in both sexes with biserial suckers proximally, tetraserial distally; remaining arm suckers biserial. Some suckers on dorsal and ventral sides of arms 2, and ventral side of arms 3 of males enlarged: suckers largest on arms 2, larger in ventral than dorsal row. Tips of arms 1 in females devoid of suckers. Males with welldeveloped keel on arms 3; arms 4 with narrow keel. Females with keels on all arms; well developed on arms 2 and 3, keel only on distal tips of arms 1. Club suckers in 5–7 series proximally, ~9 series on distal end of club.
Description. Counts and indices for a subset of individual specimens are given in Appendices 3 and 4. Measurements and counts refer to eight mature males and ten mature females.
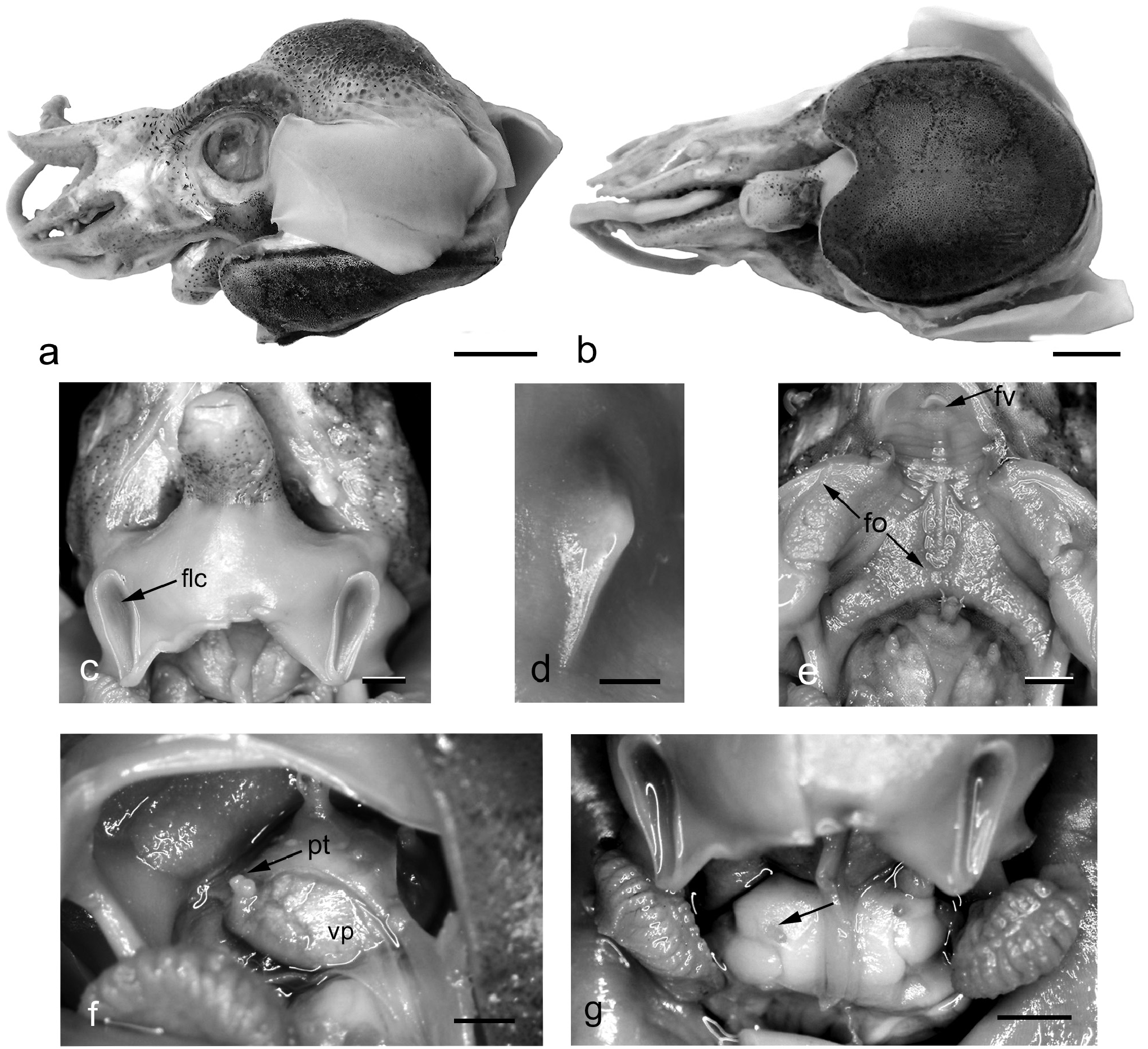
Species relatively small: ML males examined 14.1– 14.8 – 16.5 mm (SD, 0.9), females 11.2– 12.4 – 13.6 mm (SD, 0.8). Mantle short, deep, dome-shaped posteriorly ( Figure 5a, b View FIGURE 5 ), MWI males 64.3– 81.8 –98.8 (SD, 13.8), females 10.0– 11.1 –12.8 (SD, 0.9). Body deep dorso-ventrally with distinct ‘hump’ arch in dorsal mantle that extends well above head when seen in lateral view ( Figure 5a View FIGURE 5 ). Dorsal mantle contiguous with head, joined by wide occipital band that extends to level of junction with fins, OBL males 95.2– 133.1 –160.8 (SD, 24.5), females 69.2– 119.7 –178.6 (SD, 36.9). Ventral mantle convex and covers base of funnel; anterior margin with deep ‘U’-shaped indentation ventral to funnel ( Figure 5b View FIGURE 5 ). VMLI males 102.4– 115.2 –127.0 (SD, 9.4), females 94.9– 108.7 –147.3 (SD, 16.7). Ventral shield clearly demarcated, with densely packed chromatophores and darker outline ( Figure 5b View FIGURE 5 ); covers almost entire ventral surface of body VSLI males 65.0– 90.9 –113.3 (SD, 19.9), females 83.3– 96.5 –115.2 (SD, 12.5). Fins oval; maximum length approximately equal to mantle length, with blunt point posteriorly; positioned close to anterior mantle margin; FIIa males 12.1– 16.0 –20.0 (SD, 2.7), females 13.4– 18.5 –23.1 (SD, 3.1); fin width about 65% ML; FWI males 49.0– 63.8 –76.6 (SD, 9.4), females 57.5– 66.8 –79.5 (SD, 7.7); attached dorso-laterally midway along mantle, approximately level with dorsal margin of eye; FII males 49.6– 54.9 –60.3 (SD, 4.4), 43.1– 50.4 –63.6 (SD, 6.6). Fins do not project anteriorly beyond anterior dorsal mantle margin ( Figure 5a View FIGURE 5 ).
Funnel conical, projects anteriorly beyond ventral mantle margin. FuLI males 60.0– 62.7 –68.8 (SD, 3.2), females 50.0– 60.8 –78.1 (SD, 9.9); free for most of its length, FFuI males 57.6– 63.9 –70.9 (SD, 4.6), females 38.5– 51.9 –67.0 (SD, 9.9). Dorsal side of funnel distinctly swollen. Funnel-locking cartilage ( Figure 5c View FIGURE 5 ) broadest anteriorly, tapering posteriorly with curved groove, deepest anteriorly. Mantle cartilage strongly angled corresponding to funnel groove ( Figure 5d View FIGURE 5 ). Funnel valve a small flap ( Figure 5e View FIGURE 5 ). Funnel organ ( Figure 5e View FIGURE 5 ) dorsal element broad, inverted V-shape, with ridge mid-anteriorly; ventral elements ovoid.
Head broader than long, HLI males 47.6– 68.9 –94.3 (SD, 15.2), females 53.4– 68.1 –83.3 (SD, 11.5); HWI males 53.1– 71.2 –85.1 (SD, 10.9), females 63.2– 72.5 –81.3 (SD, 6.4). Eyes large, EDI males 42.0– 60.8 –82.3 (SD, 16.1), females 46.2– 55.8 –65.0 (SD, 7.1); ventral eyelids free ( Figure 5a View FIGURE 5 ). Distinct, large photosensitive vesicle on lateroposterior surface of head, ventral to eyes.
Visceral photophore large, round, iridescent greenish-yellow in preserved specimens, underlies median mantle septum (septum divides mantle cavity and extends posteriorly from anus to join the ventro-posterior mantle). Photophore bears paired photophore tubes on each side; photophore tubes joined basally, with tiny ovoid openings ( Figure 5e, f View FIGURE 5 ). Anteriorly, a narrow band of muscular tissue connects the paired photophore tubes with base of funnel. Renal openings prominent inside mantle cavity ( Figure 5g View FIGURE 5 ).
All arms in both sexes very similar in length with arms 2 only slightly longer than other arms in males ( Table 3). Arm length index of longest arm in males (ALI2) 60.6– 76.2 –92.2 (SD, 12.3), females (ALI4) 40.0– 55.0 –80.4 (SD, 13.2). Males with well-developed keel on arms 3; arms 4 with narrow keel on distal 2/3 of arms. Aboral keel on all arms of females: keel only at distal tip of arm 1, arms 2 and 3 with long, broad flange-like keel, arms 4 with narrow keel. Arms 1–3 connected by deep webs, protective membranes absent. Arm sucker pedicels with tiny lappets. Arm suckers biserial on arms 1–3, arms 4 with biserial suckers basally for approximately eight rows, tri- and tetraserial suckers after the 8 th row in both sexes.
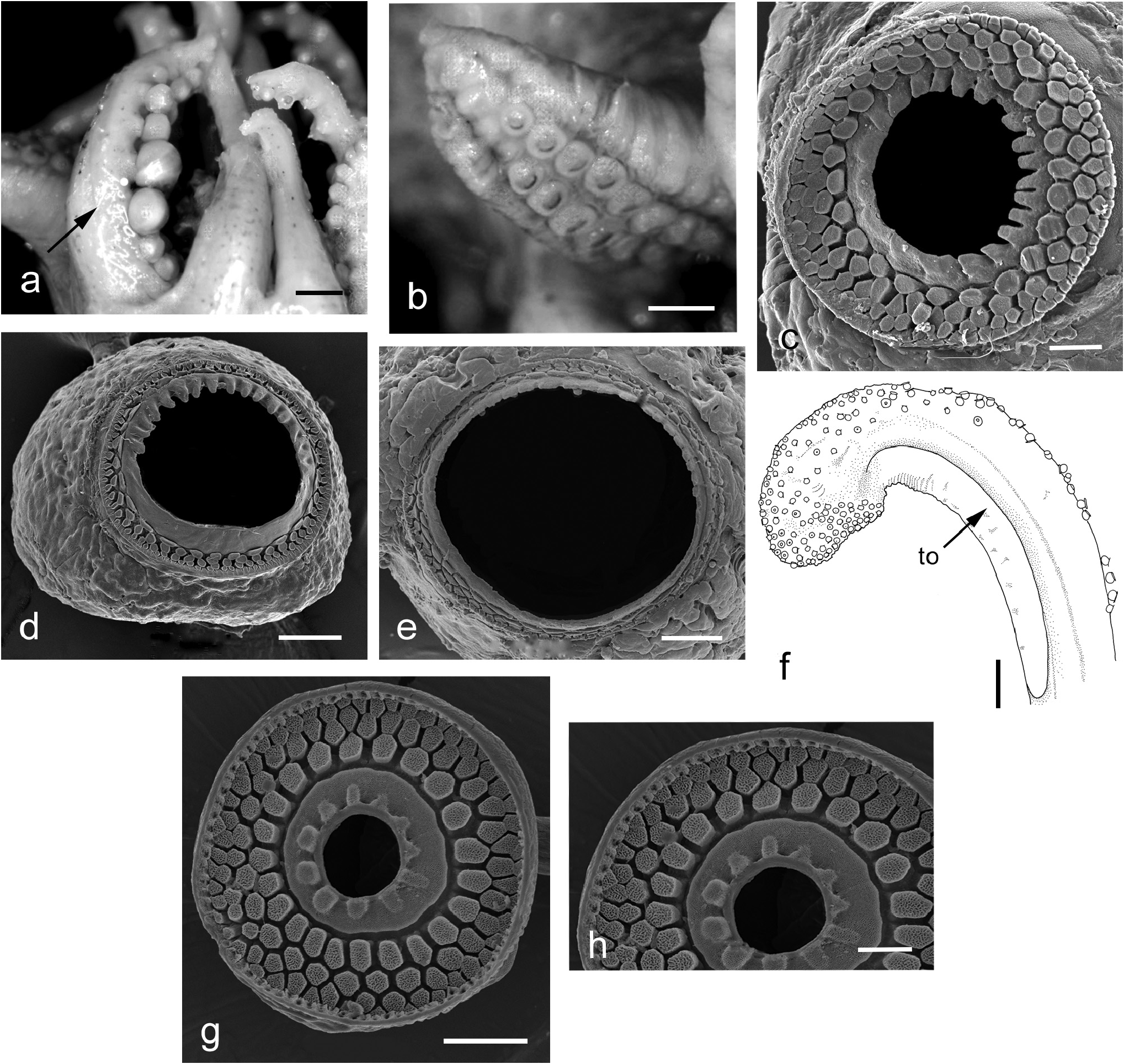
Males with enlarged suckers on arms 2 and 3 ( Figure 6a View FIGURE 6 ), largest on arms 2 ( Table 3); arms 2 with suckers in ventral rows (counting from the base) ~5–8 (usually 6–7) enlarged, suckers in rows 6 and 7 on the dorsal side enlarged, but are not as large as those on the ventral side; suckers on ventral side of arms 3 in rows ~3–6 enlarged. In both sexes distal portion of arms 4 after ~8th sucker row in 3–4 rows, suckers minute, crowded. Tips of arms 1 in females devoid of suckers. Female arm suckers slightly smaller on dorsal arm pairs than ventral arm pairs, but otherwise not markedly enlarged.
Right dorsal arm of males hectocotylised, swollen laterally, fleshy ( Figure 6b View FIGURE 6 ), more swollen on dorsal side of arm, swollen part divided transversely into ridges; left dorsal arm swollen slightly, swelling greater on ventral than dorsal side of arm. Hectocotylus suckers uniform in size. Arms 2 slightly swollen basally.
Sucker counts range from 20–36 on arms 1–3; Arms 4 with a much greater number of suckers than other arms in both sexes (50–70) ( Table 3).
Chitinous arm sucker rims: infundibulum with 3–4 rows of pavement-like processes ( Figure 6c–e View FIGURE 6 ), peripheral sucker rim processes wedge-shaped, rest irregular, roughly hexagonal. Chitinous inner rim of normal arm suckers with teeth on one side, very slightly crenulated on other side ( Figure 6c, d View FIGURE 6 ). Suckers on modified region of hectocotylus with wide, smooth openings, slightly crenulated, without teeth ( Figure 6e View FIGURE 6 ). Pavement-like processes smooth distally.
Tentacles slender, stalks naked, semicircular in section. Club relatively short; ClLI males 27.3– 34.3 –42.6 (SD, 6.4), females 29.7– 44.2 –58.3 (SD, 12.0), tapers to blunt end distally; sucker-bearing face convex, recurved and joined to stalk ( Figure 6f View FIGURE 6 ). Suckers approximately 0.1 mm in diameter in centre of club; arranged in 5–7 crowded oblique series proximally; distally, club with up to 9 transverse series of suckers, suckers slightly smaller distally than on basal half of club. Aboral keel absent; tentacle organ extends along proximal half of sucker-bearing portion of club and along tentacle stalk beyond club sucker-bearing portion; groove extends along length of club ( Figure 6f View FIGURE 6 ). Club sucker dentition ( Figure 6g, h View FIGURE 6 ): inner ring without teeth, and bearing indistinct pegs that are more prominent on one side; infundibulum with three rows of pavement-like processes; processes uniform width from base to distal margins; inner row sub-rectangular to hexagonal; middle and outer rows ovoid; irregular, with strongly pitted surfaces ( Figure 6h View FIGURE 6 ).
Gills with 21–22 lamellae per demibranch; GLI males 32.2– 38.9 –42.6 (SD, 4.2), females 31.5– 40.5 –50.0 (SD, 5.9). Buccal membrane with six lappets; suckers absent.
Upper beak ( Figure 7a View FIGURE 7 ) with pointed rostrum, hood convex, high above crest posteriorly; jaw angle approximately 90 degrees; lateral wall edge with slight indentation. Lower beak ( Figure 7b View FIGURE 7 ) with pointed, short rostrum, rostral edge obtuse, curved, with distinct inner angle; hood notch absent, wings straight. Distinct dark pigmentation restricted to rostrum and hood of upper and lower beaks.
Radula with seven transverse series of teeth ( Figure 7c View FIGURE 7 ). Rhachidian simple, without cusps, triangular, concave laterally with wide bases; bases only slightly convex. First lateral teeth with pointed cusps, strongly displaced laterally, with distinct ‘heel’ directed towards midline of radula. Second and third laterals with elongate bases, curved, scythe-like, third laterals longer than second laterals.
Gladius absent.
Male reproductive tract ( Figure 7d View FIGURE 7 ) typical for the family but with spermatophore storage sac in two distinct sections. Spermatophores approximately half mantle length SpLI 51.0– 59.6 –67.4 (SD, 6.7), SPWI 1.7– 2.5 –3.3 (SD, 0.6). Spermatophore ejaculatory apparatus ( Figure 7e View FIGURE 7 ) bipartite; cylindrical, broadest posteriorly, tapering to a nipple-like duct that connects with sperm reservoir. Oral end of ejaculatory apparatus with simple coils.
Female reproductive tract ( Figure 7f View FIGURE 7 ): Ovary occupies large proportion of posterior end of mantle cavity. Nidamental glands paired, broad, located ventral to ovary, almost completely covering ovary. Large, cream-coloured accessory nidamental glands located toward distal end of nidamental glands. Large sac-like bursa copulatrix on animals’ left side. Some specimens with spermatangia embedded in bursa. EgDI 7.4– 12.7 –16.9 (SD, 3.6).
Colour. Alcohol preserved specimens with rich coloration: maroon with large deep purple chromatophores on dorsal head and mantle; spots small and animal darker on ventral surface; ventral shield bordered by dark U-shaped margin ( Figure 5a, b View FIGURE 5 ); few scattered chromatophores at base of arms. Body with distinct metallic sheen, most prominent at base of arms, dorsal side of fins and surrounding ventral shield. Funnel with scattered chromatophores in a band covering the siphon on ventral side only; distal tip and base of funnel unpigmented, without chromatophores. Fins, arms and tentacles largely unpigmented. Club unpigmented.
Habitat. All specimens were collected in midwater trawls.
Type locality. New Zealand: west of New Plymouth , 39°2.27´S, 172°13.5´E GoogleMaps .
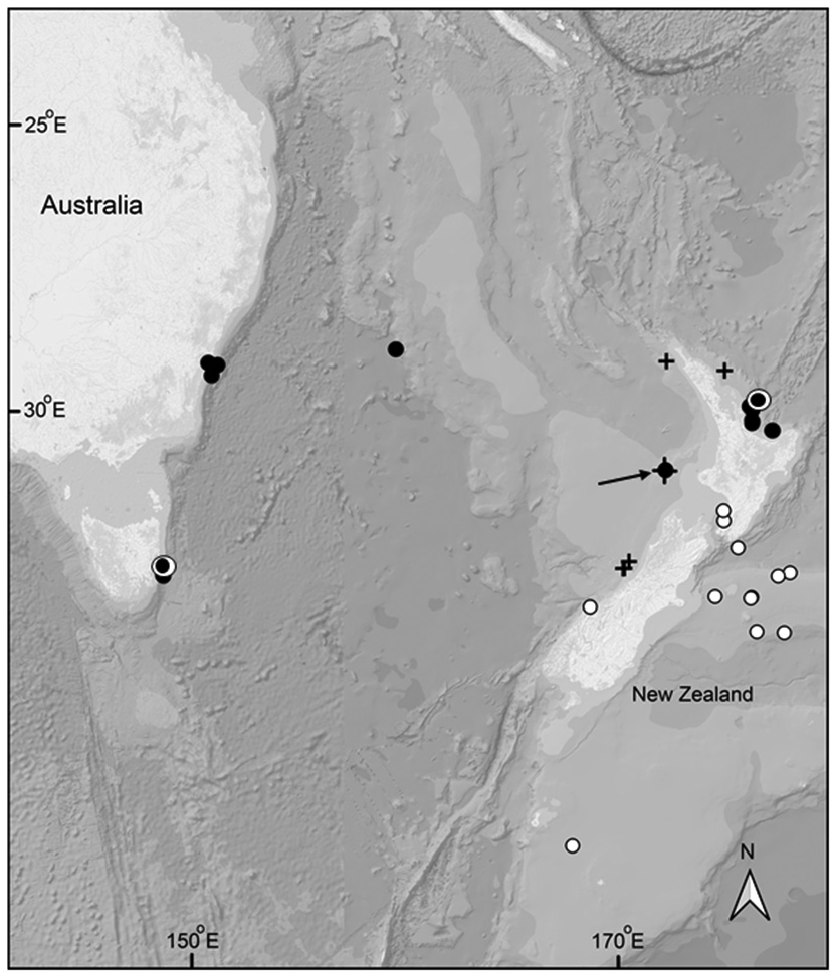
Distribution. New Zealand: from SE of Mercury Islands, 36°46.8´S, 176°18.5´E to off White Island, 37°33.7´S, 177°14.90´E. Southern and southeastern Australia from approximately 34°S where it appears to be common ( Figure 11 View FIGURE 11 ). Collection depths range from approximately 114–350 m over bottom depths ranging from 400 to 1400 m.
Etymology. This species is named in honour of Professor Merlin Crossley of the University of New South Wales, Australia, an eminent researcher in the fields of genetics and microbiology, in recognition of his highly valued contribution in serving on the Trust of the Australian Museum from 2012 to 2021. He has played a pivotal role in supporting and promoting science at the Australian Museum.
Comments. The future study of specimens held in all Australian museum collections will provide additional information on the distribution of this species. Its pelagic lifestyle suggests it is likely to be widespread. Based on its recognition by C.C. Lu among Museum Victoria (MV) collections some decades ago ( Lu & Dunning 1998), and examination of current Atlas of Living Australia Records (http://www.ala.org), future work will undoubtedly yield additional information and considerably expand the known distributional range of this species, particularly around Australia.
Future molecular studies will also provide more information on this species and enable its phylogenetic position within the Sepiolidae to be better understood. This family is currently a focus of extensive molecular research worldwide. Formal naming of this taxon, is therefore, timely and will assist in species identifications to underpin such studies.
This is only the second described species in the genus, with Iridoteuthis iris known from the Hawaiian Archipelago. Another, as yet undescribed species from off New Caledonia has also been reported as Iridoteuthis sp. A (Young & Vecchione 2007). Iridoteuthis merlini is clearly most closely related to Iridoteuthis iris , and they are likely sister species. The main differences distinguishing the currently recognised members of this genus are shown in Table 4.
If adult material from Japan becomes available, it will be important to compare I. merlini with Stoloteuthis japonica Kubodera & Okutani, 2011 . Young & Vecchione (2007) suggest that S. japonica may actually be a synonym of I. iris . It was not possible to examine the S. japonica type specimen but its description suggests it may indeed belong to Iridoteuthis due to its huge eyes, crescent-shaped mantle-locking cartilage and shallower arm webs than those seen in Stoloteuthis . As the type is immature, and the light organ morphology difficult to determine from the figures, it is hard to be sure of its correct generic placement. However, the arm sucker counts of the S. japonica holotype appear to differ from those of I. merlini , with the total number of arm suckers in S. japonica reported as Arms 1, ~30 (v. 35–36 in I. merlini ); Arms 2, ~40 (v. 22–26); Arms 3, ~40 (v. 20–22); Arms 4 ~30 (v. 56–70). In addition, the median radula teeth appear to be slenderer and more pointed in S. japonica than those of S. merlini , although the amount of intra specific variation in this character and variation due to the size specimens is unknown.

| NMNZ |
Museum of New Zealand Te Papa Tongarewa |
| AM |
Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Iridoteuthis merlini
| Reid, Amanda L. 2021 |
Iridoteuthis sp.
| Lu, C. C. & Dunning, M. C. 1998: 513 |