
Pholcophora Banks, 1896
|
publication ID |
https://doi.org/ 10.5852/ejt.2023.880.2173 |
|
publication LSID |
lsid:zoobank.org:pub:3F806FD6-2EB3-456A-AFD7-780A0FBEB2DA |
|
DOI |
https://doi.org/10.5281/zenodo.8160844 |
|
persistent identifier |
https://treatment.plazi.org/id/03F087F1-AC7E-FFDB-0941-62B7E16AFE94 |
|
treatment provided by |
Felipe |
|
scientific name |
Pholcophora Banks, 1896 |
| status |
|
Genus Pholcophora Banks, 1896 View in CoL View at ENA
Pholcophora Banks, 1896: 57 View in CoL .
Type species: P. americana Banks, 1896 View in CoL .
Pholcophora View in CoL – Gertsch 1971: 76; 1977: 112; 1982: 96. — Huber 2000: 113 View Cited Treatment .
Diagnosis
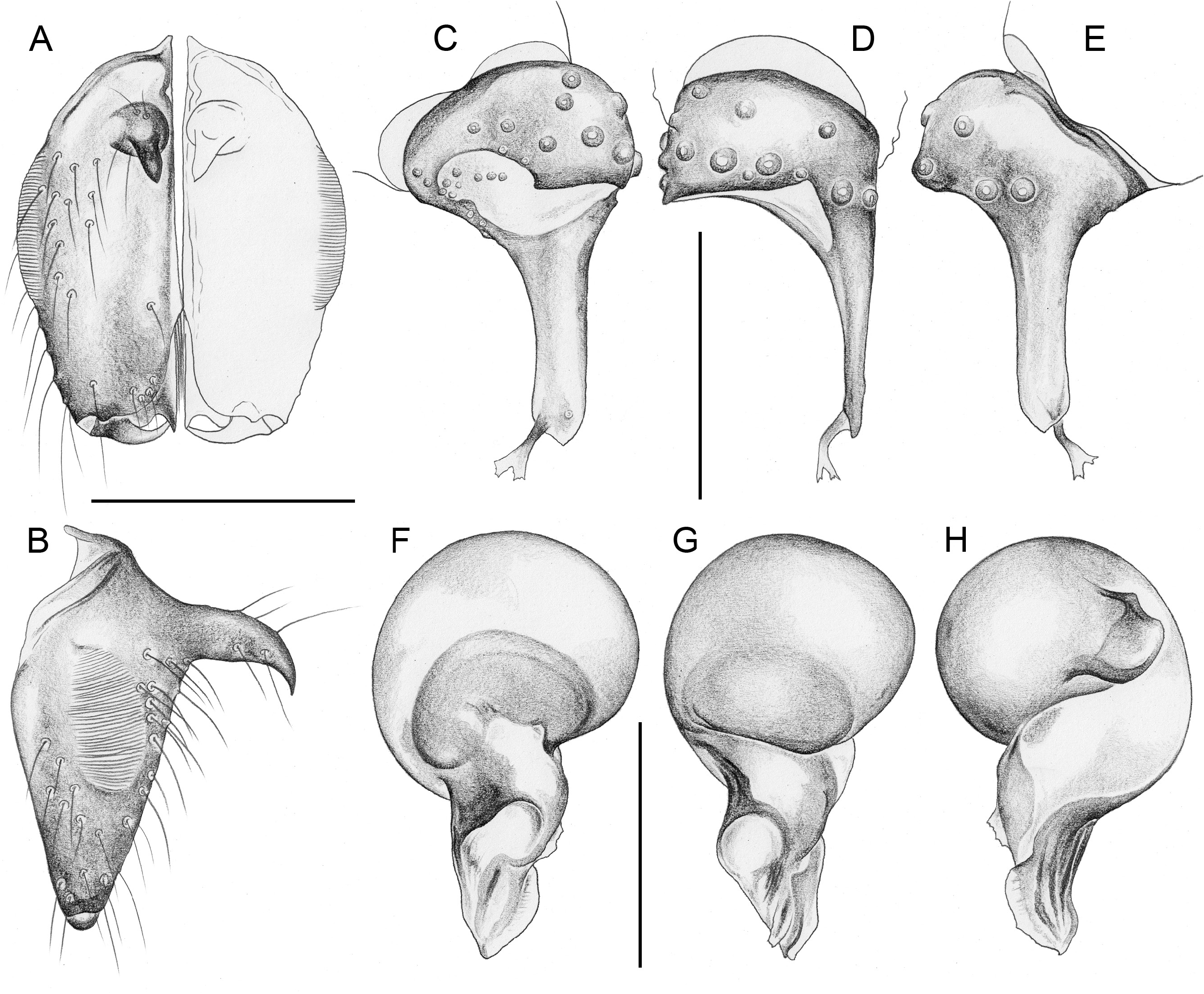
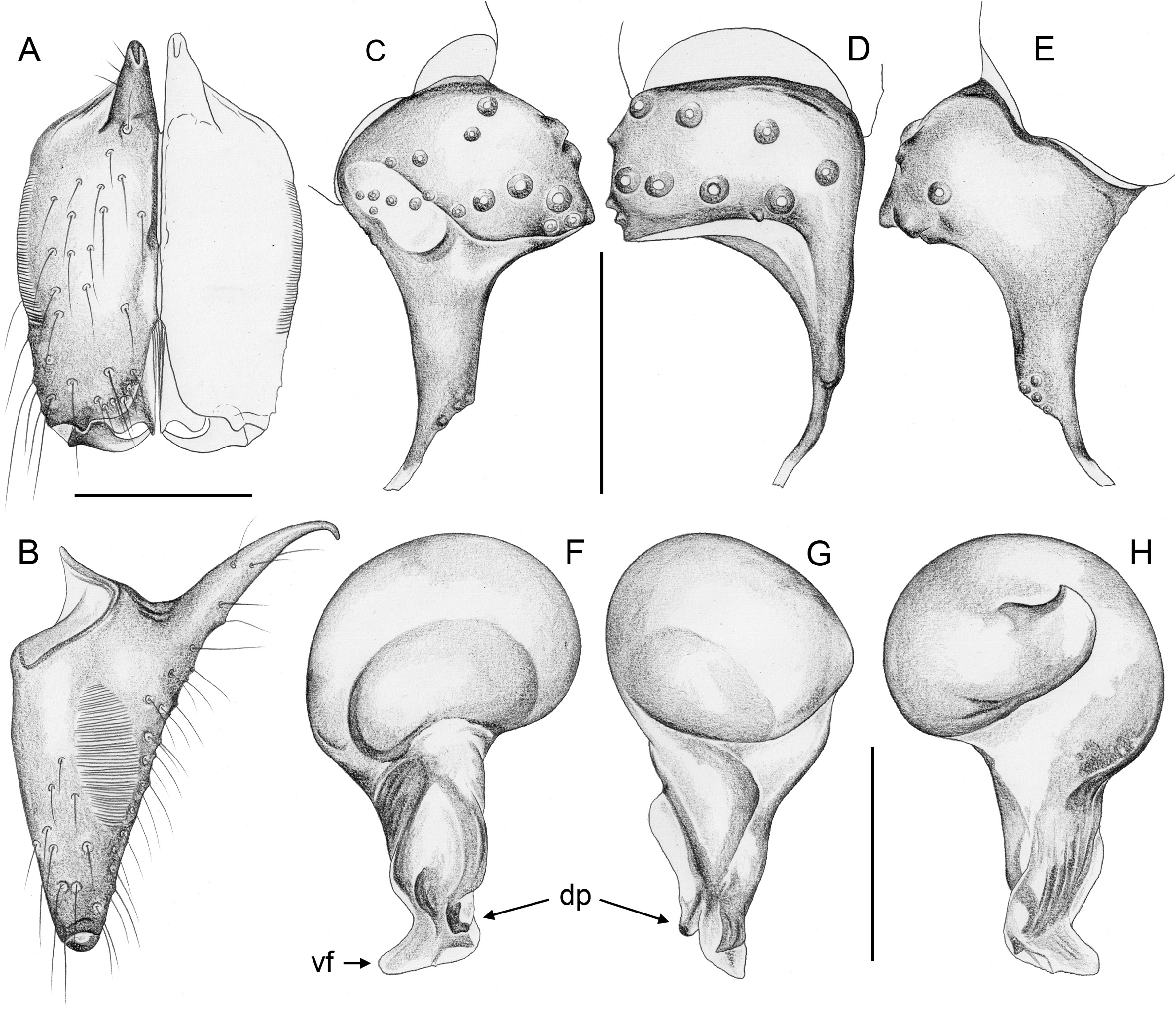
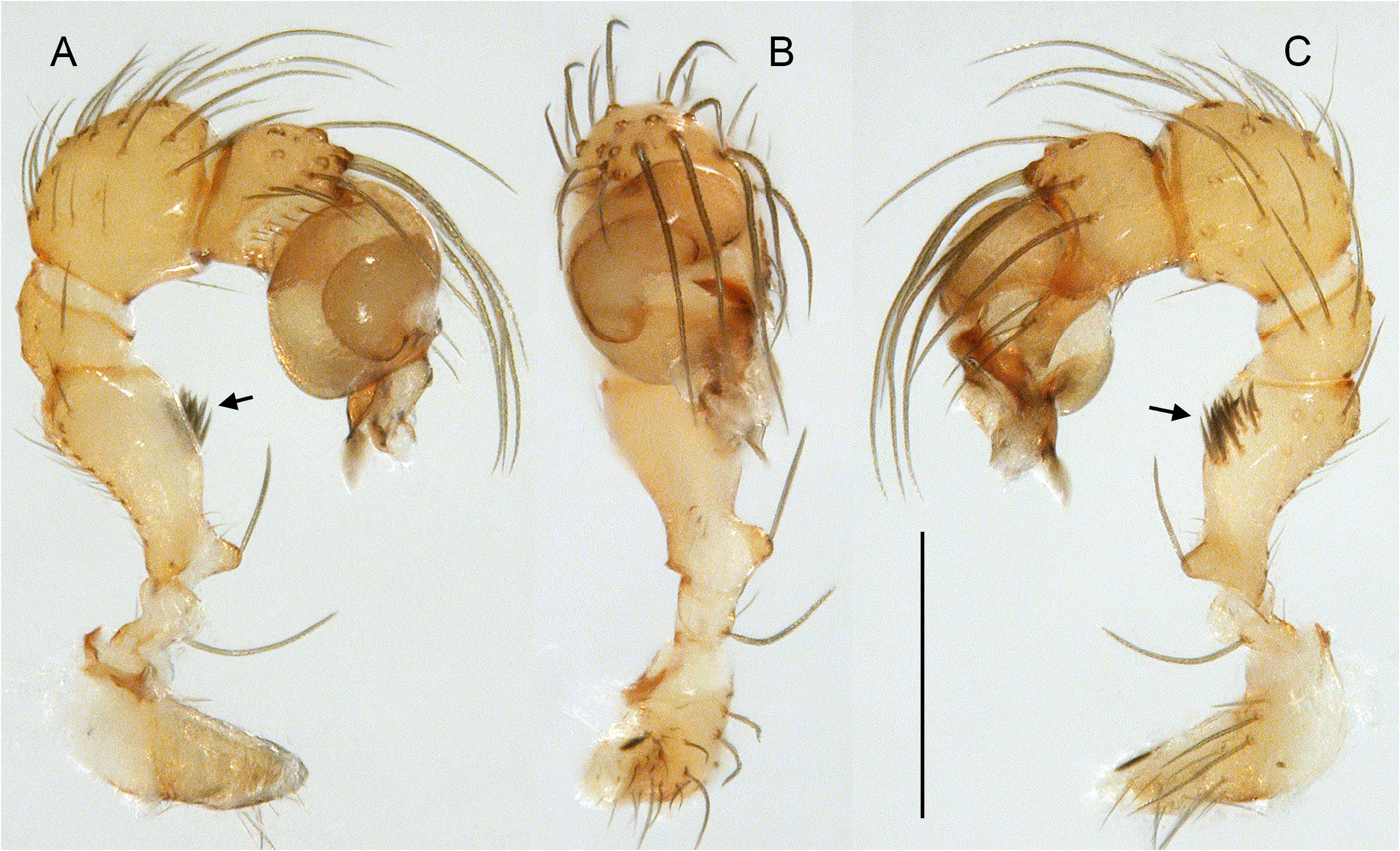
Easily distinguished from only other North American Ninetinae genus Tolteca by strong male cheliceral apophyses originating proximally ( Figs 5A–B View Fig , 10A–B View Fig ; in Tolteca small and originating distally); also by presence of stridulatory ridges on male chelicerae; most species (except P. tehuacan Huber sp. nov.) also by larger size (body length ~1.7–3.1; in Tolteca ~1.1–1.4) and longer legs (tibia 1>1.0, in Tolteca <0.7); from most species of Tolteca also by absence of knob-shaped structure between epigynum and pedicel (also absent in Tolteca sinnombre Huber sp. nov.). From other geographically close genera ( Papiamenta , Galapa ) also by simple rod-shaped procursus ( Figs 5C–E View Fig , 10C–E View Fig ; much shorter in Papiamenta ; with dorsal process in Galapa ); by presence of humps on male sternum (absent in Papiamenta ); and by unmodified male cheliceral fangs (with processes in Galapa ).
Description
Male
MEASUREMENTS. Total body length 1.2–3.1, carapace width 0.55–1.40. Legs relatively short, tibia 1: 0.65–1.85, i.e., 1.2–2.0 × carapace width; tibia 1 L/d 8–16; tibia 2 much shorter than tibia 4 (tibia 2 / tibia 4: 0.75–0.85).
COLOUR. Live specimens ( Fig. 3 View Fig ) ochre to brown, prosoma sometimes reddish (which is lost in ethanol); carapace monochromous, sometimes with indistinct darker median Y-mark; abdomen colour slightly variable, usually monochromous, sometimes with indistinct dorsal marks (bluish in ethanol); legs without dark or light bands, femora sometimes distally darkened.
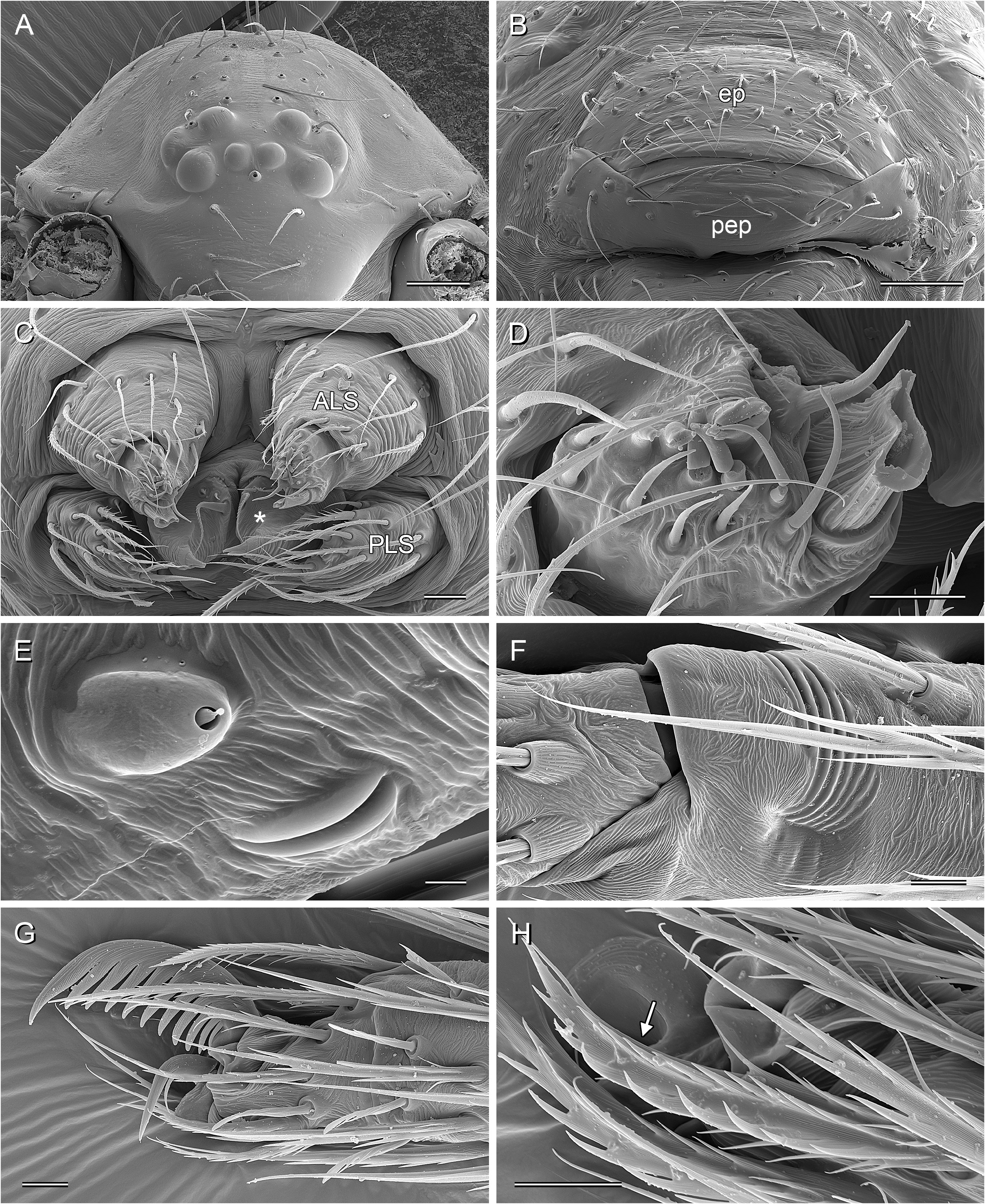
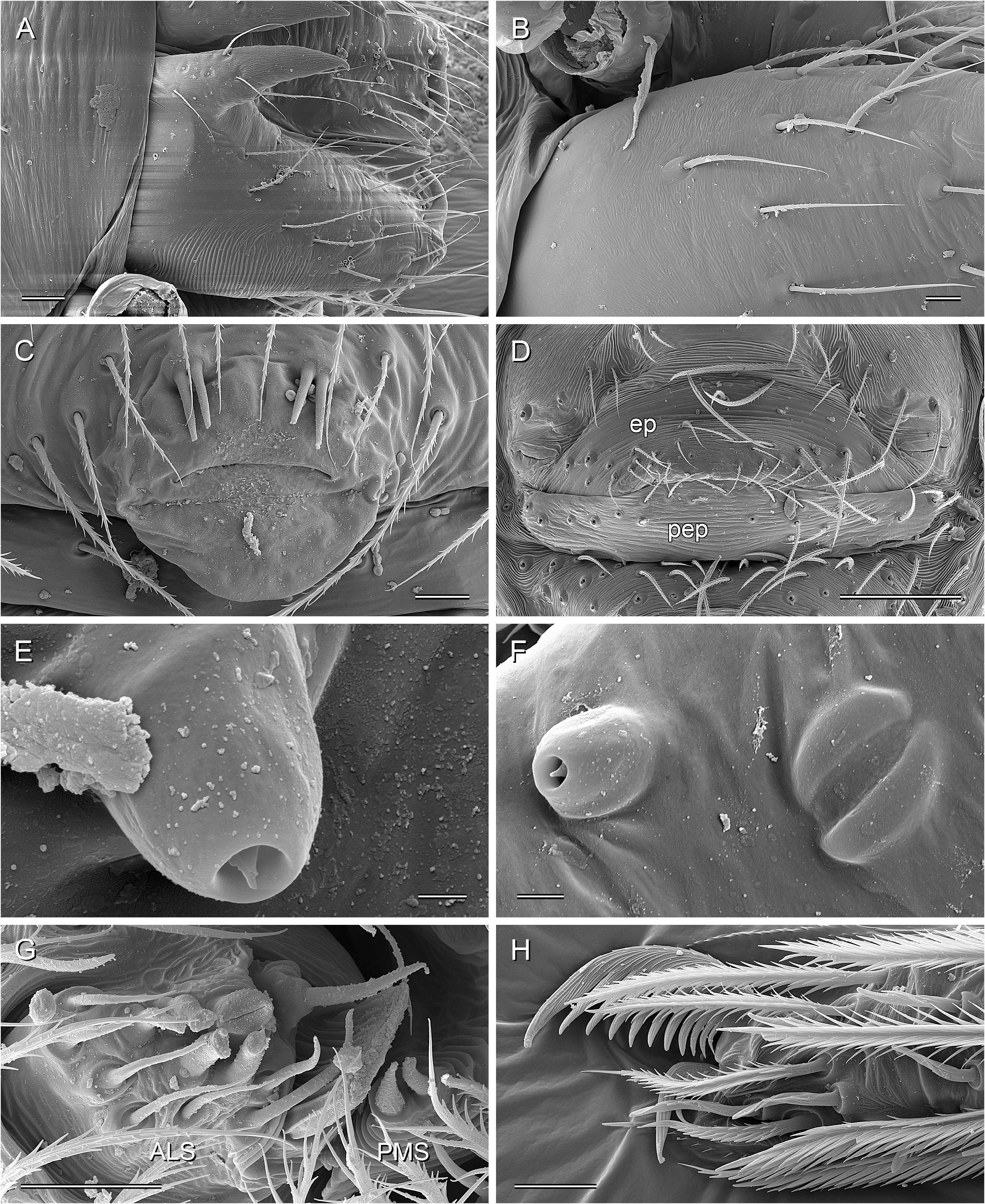
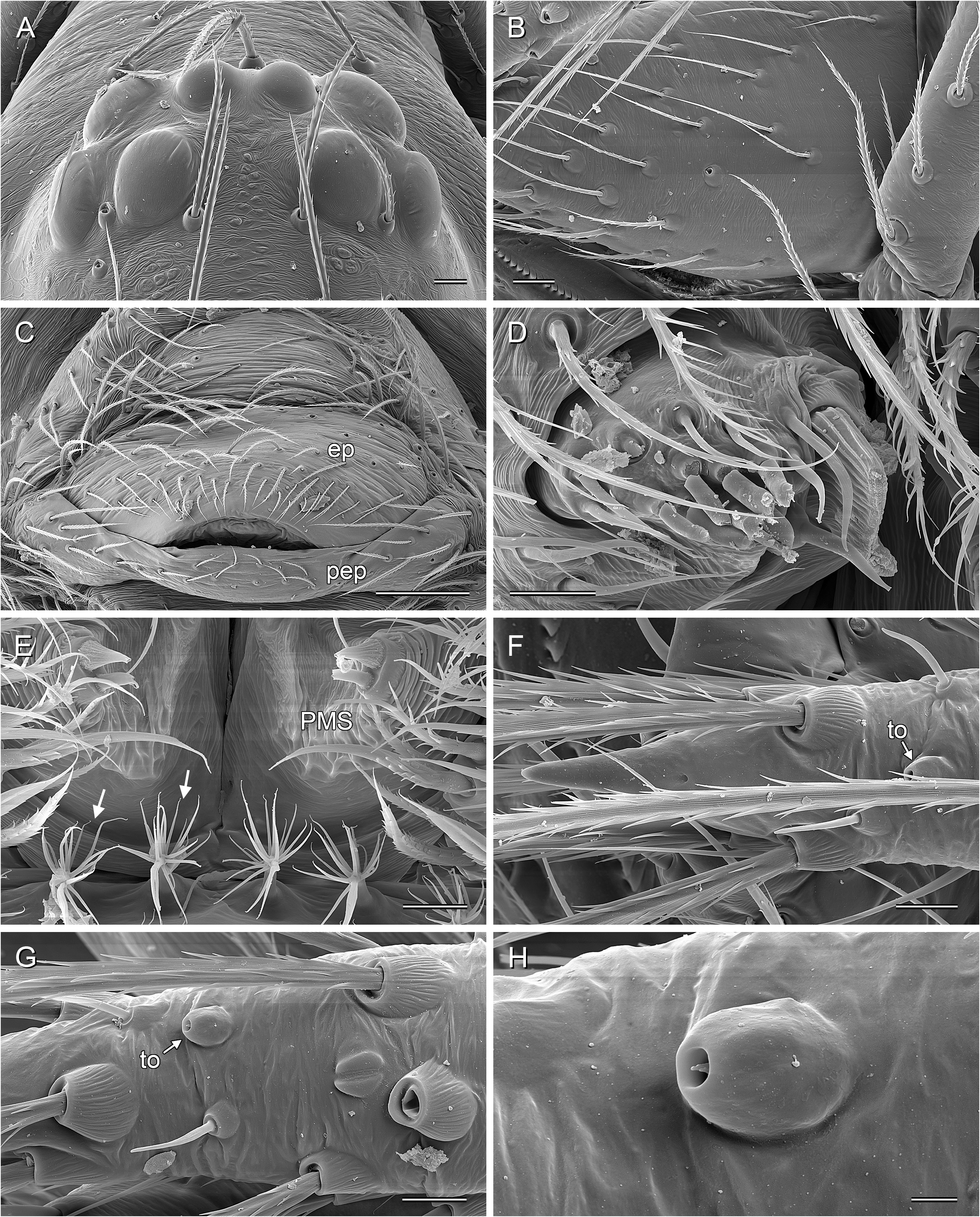
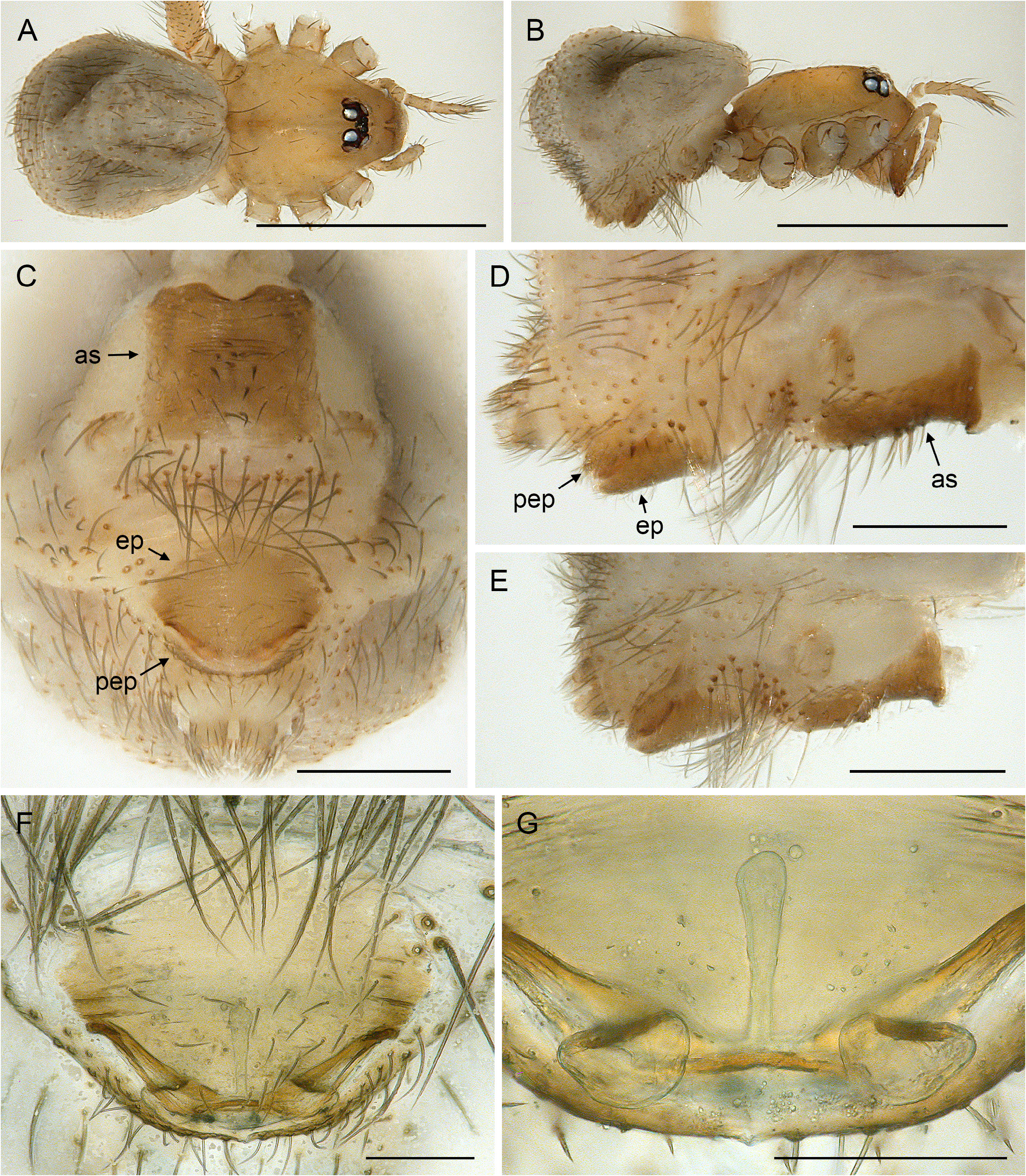
BODY. Ocular area barely raised (cf. Fig. 8A View Fig ), eight eyes, AME relatively large, diameter 20–60 µm, 45–75% of PME diameter; carapace with low and indistinct thoracic groove (cf. Figs 8A View Fig , 13A View Fig ). Clypeus unmodified but sometimes slightly more protruding than in female. Sternum wider than long, with pair of distinct anterior processes near leg coxae 1. Abdomen globular; gonopore with four epiandrous spigots arranged in two pairs ( Fig. 29C View Fig ; examined: P. americana Banks, 1896 ; P. tehuacan sp. nov.); ALS with seven spigots each (cf. Figs 8C View Fig , 29G View Fig ): one strongly widened spigot, one long pointed spigot, and five cylindrical spigots (one of which is unusually large); PMS with two short, pointed spigots; PLS without spigots.
CHELICERAE. With one pair of large frontal apophyses ( Figs 5A–B View Fig , 10A–B View Fig ); with stridulatory files ( Fig. 29A View Fig ), distances between ridges ~2.0–3.8 µm, distances proximally often smaller than distally.
PALPS. Coxa unmodified; trochanter barely modified (with indistinct or without ventral projection); femur proximally with retrolateral-ventral process and prolateral stridulatory pick (modified hair), distally widened but simple, slightly curved towards dorsal, in P. texana Gertsch, 1935 with distinctive brush of feathered hairs ventrally ( Fig. 19A, C View Fig ); femur-patella joints slightly shifted toward prolateral side; tibia globular, with two trichobothria; tibia-tarsus joints not shifted to one side; palpal tarsal organ raised, capsulate with small opening ( Fig. 29E View Fig ; diameter of opening ~1.5 µm – measured in P. tehuacan sp. nov. only); procursus simple and straight, without dorsal flap, not strongly elongated, often with semi-transparent distal element; genital bulb distally cone-shaped, with species-specific sclerotized and membranous elements.
LEGS. Without spines and curved hairs; in some species with very short vertical hairs in higher than usual density on tibiae (only anterior tibiae or all tibiae) (length of hairs ~30 µm). Trichobothria in usual arrangement: three on each tibia (except tibia 1: prolateral trichobothrium absent), one on each metatarsus; slightly feathered ( Figs 18F View Fig , 24E View Fig ); length of dorsal trichobothrium on tibia 1: ~90 µm; retrolateral trichobothrium of tibia 1 in very distal position (at 50–65% of tibia length). Tarsus 1 with 5–8 pseudosegments, sometimes only distally fairly distinct; tarsus 4 distally with one comb-hair on prolateral side (cf. Figs 13H View Fig , 24H View Fig ); leg tarsal organs very small, capsulate with small opening (cf. Fig. 8F–H View Fig ; diameter of opening ~1.3–1.8 µm); three claws (cf. Figs 13G View Fig , 24G View Fig , 29H View Fig ).
Female
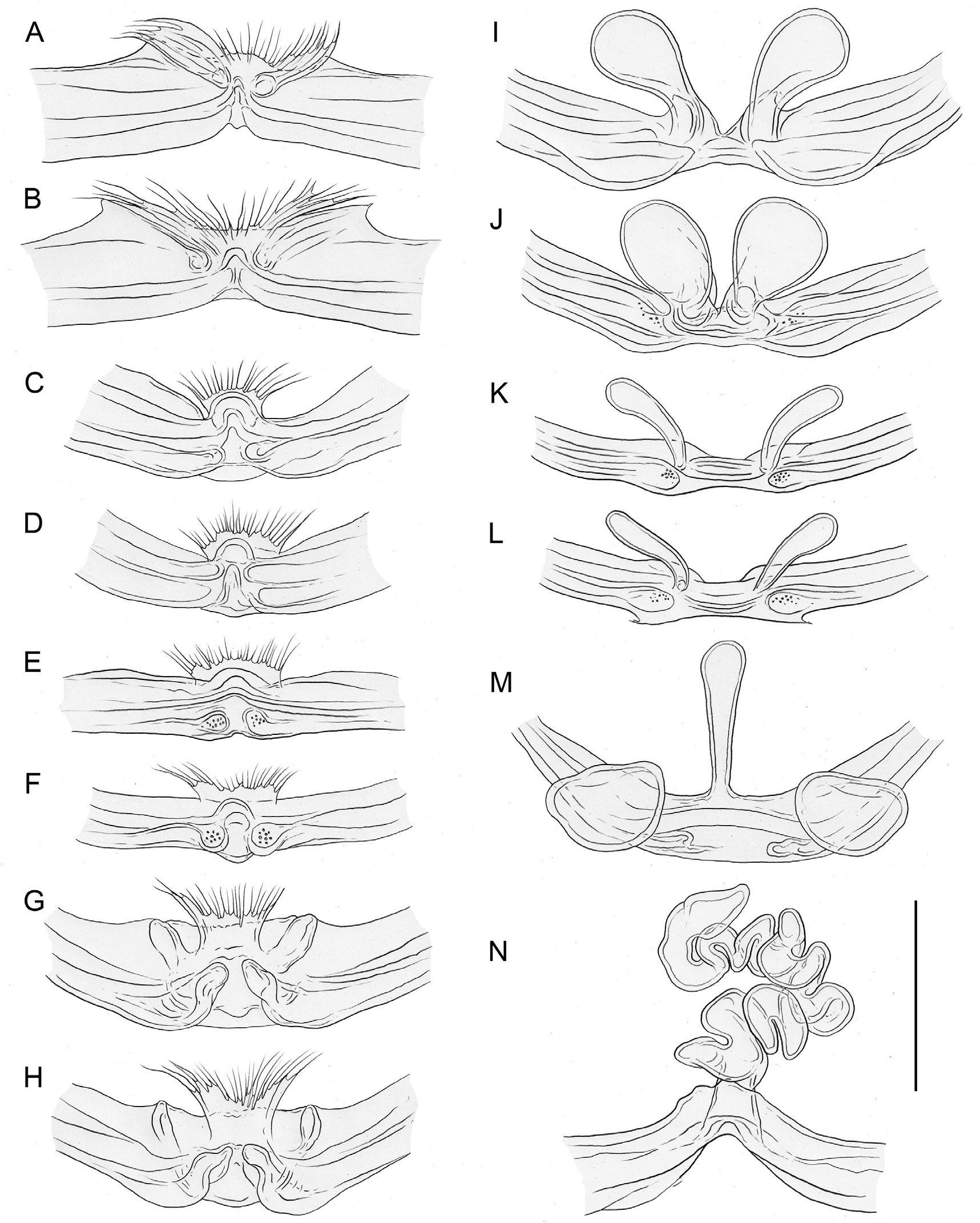
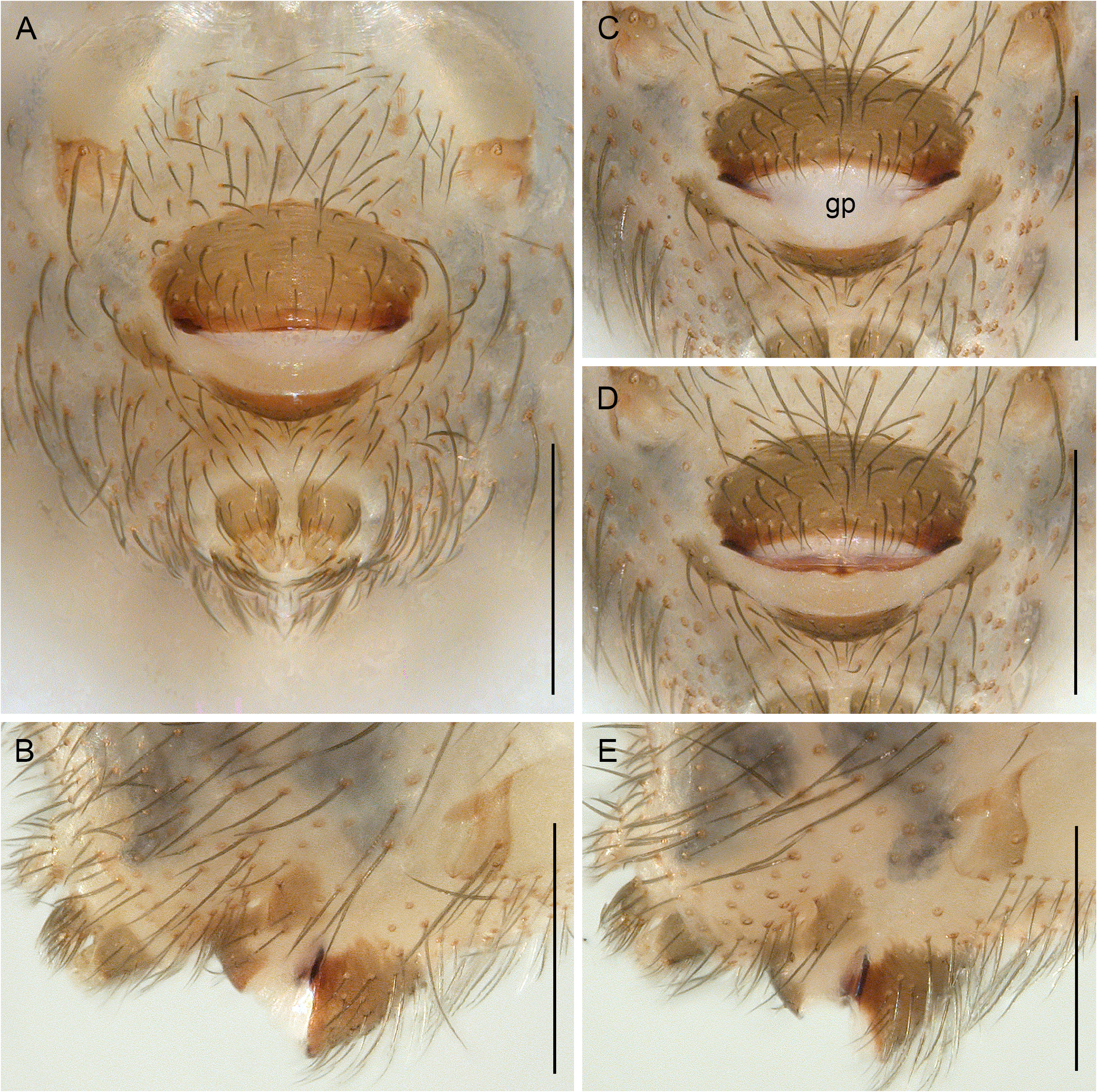
In general (size, colour) similar to male but sternum without pair of anterior humps, leg tibia 1 with usual low number of short vertical hairs, and chelicerae without stridulatory ridges; legs either slightly shorter than in males or of same length (only P. americana with reasonable sample size: male/female tibia 1 length: 1.12). Spinnerets, comb-hairs, and leg tarsal organs as in male; palpal tarsal organ slightly less strongly raised ( Figs 18E View Fig , 23H View Fig ). Epigynum main (anterior) plate large, rectangular to oval, sometimes posteriorly excavated, weakly protruding in lateral view; posterior plate also large, short but wide, median part sometimes separated from lateral parts by whitish band. Without knob-shaped structure between epigynum and pedicel. Internal genitalia variable: either without sacs ( Fig. 33A–H; P View Fig . americana Banks, 1896; P. mazatlan Huber sp. nov.; P. papanoa Huber sp. nov.; P. “Mex354”) or with pair of distinct membranous sacs ( Fig. 33I–L; P View Fig . texana Gertsch, 1935; P. tehuacan sp. nov.); in Caribbean ‘ Pholcophora ’ with single median tube-like sac ( Fig. 33M–N; P View Fig . bahama Gertsch, 1982; P. “ Car 544”; P. “Cu12-325”); with pair of distinct transversal sclerites; apparently without pore plates (possibly with very indistinct pair of pore plates near median line, indicated in Fig. 33E–F, J–L View Fig ).
Relationships
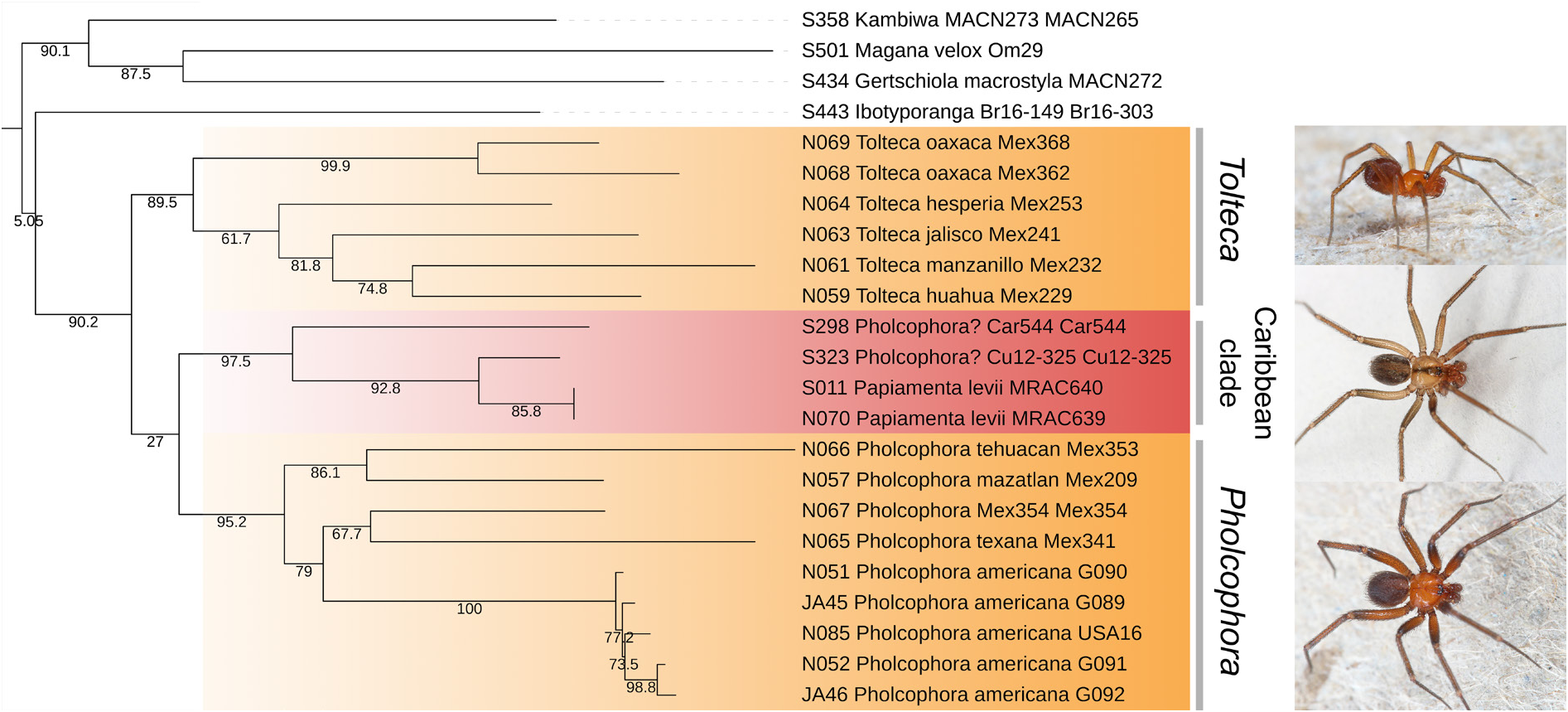
In the molecular analysis of Eberle et al. (2018), the type species Pholcophora americana was part of a North American-Caribbean clade of Ninetinae , together with “ Pholcophora ? Car 544”, an unidentified Cuban species (“Gen. Cu12-325”; treated below as “ Pholcophora ? Cu12-325”), and the genus Papiamenta . The genus Tolteca was not included.
Our new molecular analyses mostly support this North American-Caribbean clade ( Fig. 1 View Fig ; see also general results of molecular analyses above), and they also support the idea that Caribbean ‘ Pholcophora ’ are not true Pholcophora but more closely related with the Caribbean genus Papiamenta . Our analyses suggest a sister group relationship between true Pholcophora and the Caribbean clade ( Papiamenta + Caribbean ‘ Pholcophora ’). By contrast, preliminary analyses of UCE data (G. Meng, L. Podsiadlowski, B.A. Huber, unpubl. data) suggest a sister-group relationship between true Pholcophora and Tolteca . Since we consider these UCE results more reliable, we will not further discuss relationships here, except for two species not yet included in any molecular dataset: Pholcophora maria Gertsch, 1977 from Yucatán, and Pholcophora bahama Gertsch, 1982 from the Bahamas. Judging from the female internal genitalia (compare Huber 2000: fig. 1357 with Fig. 28B View Fig ), we hypothesize that Pholcophora maria is closely related with the newly described P. tehuacan sp. nov. It is thus probably a true Pholcophora . Pholcophora bahama resembles “ Pholcophora ? Car 544” in having a median tube-like sac in the female internal genitalia (compare Huber 2000: fig. 1356 with Fig. 31G View Fig ). It is thus probably not a true Pholcophora but part of the Caribbean clade.
Distribution
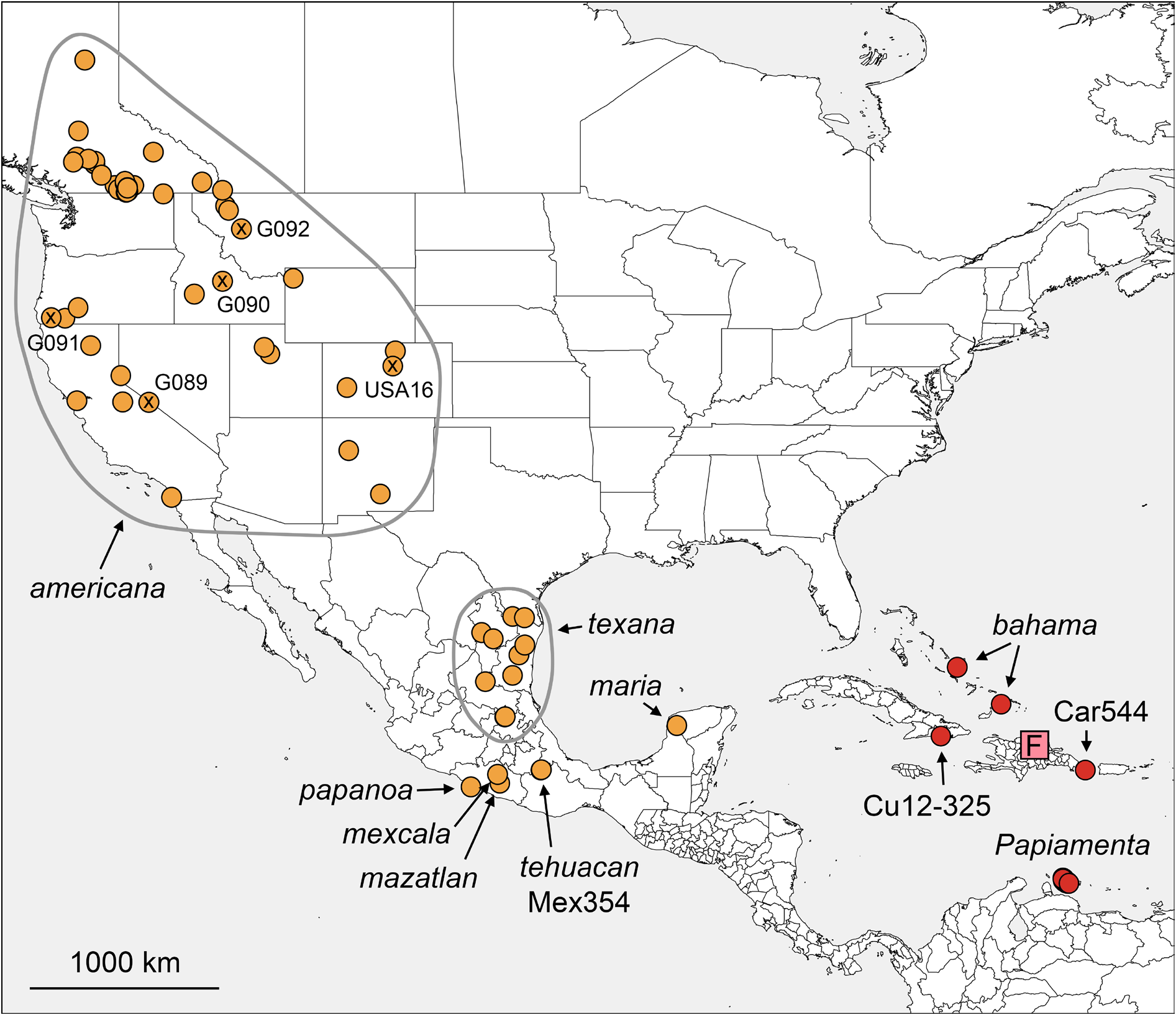
The genus appears limited to North America ( Fig. 2 View Fig ); Caribbean species (including Dominican amber fossils) currently placed in Pholcophora are probably misplaced (see Relationships above). Of the seven North American named species, six are largely or entirely restricted to Mexico.
Natural history
Gertsch (1982) briefly characterized Pholcophora spiders as living “reclusive lives under ground objects, in leaf and plant detritus, and in soil openings and caves”, and mentioned that they “spin web tangles in dark spaces and remain there in close contact with such webs as permanent residents, often in informal colonies”. Our newly collected species and specimens fit this description. Most were collected by turning rocks or sifting litter in shady spots of low and dry forests ( Fig. 34 View Fig ). They usually shared the microhabitat with one or more other pholcids. In only one case we found two species of Pholcophora at a single locality; we never found Pholcophora to share a locality with Tolteca . Females carried their flattened egg-sacs slightly under the prosoma ( Fig. 3 View Fig ); eggs sacs contained ~6– 30 eggs, each with a diameter of ~ 0.40–0.60 mm ( Huber & Eberle 2021). Some females had a genital plug (cf. Fig. 16C View Fig ).
Composition
The genus now includes 11 nominal species. Of these, seven occur in North America and are here considered to represent ‘true’ Pholcophora : Pholcophora americana Banks, 1896 ; P. maria Gertsch, 1977 ; P. mazatlan sp. nov.; P. mexcala Gertsch, 1982 ; P. papanoa sp. nov.; P. tehuacan sp. nov.; P. texana Gertsch, 1935 . The Caribbean P. bahama Gertsch, 1982 is here considered to be misplaced (see Relationships above). All extant species are treated below except P. bahama and P. mexcala for which we have no new data [in 2019 we searched at four localities close to Mezcala (= “Mexcala”) but could not find P. mexcala ].
Three nominal species are only known from amber fossils originating from Hispaniola ( Wunderlich 1988): P. brevipes Wunderlich, 1988 ; P. gracilis Wunderlich, 1988 ; and P. longicornis Wunderlich, 1988 . We did not re-examine the amber specimens but judging from their geographic origin we speculate that the three species are part of the clade including Caribbean ‘ Pholcophora ’ and Papiamenta . However, the three amber species fit the diagnosis above with respect to the strong male cheliceral apophyses originating proximally and the simple rod-shaped procursus. They are unusually small (but in this respect similar to the exceptionally small extant P. tehuacan sp. nov. and P. maria ), and the original descriptions remain silent about male cheliceral stridulation and male sternal humps. The females of the three fossil species remain unknown.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Ninetinae |
Pholcophora Banks, 1896
| Huber, Bernhard A., Meng, Guanliang, Valdez-Mondragón, Alejandro, Král, Jiří, Ávila Herrera, Ivalú M. & Carvalho, Leonardo S. 2023 |
Pholcophora
| Huber B. A. 2000: 113 |
| Gertsch W. J. 1982: 96 |
| Gertsch W. J. 1977: 112 |
| Gertsch W. J. 1971: 76 |
Pholcophora
| Banks N. 1896: 57 |