
Hyriopsis panhai, Jeratthitikul & Paphatmethin & Zieritz & Lopes-Lima & Ngor, 2021
|
publication ID |
https://doi.org/10.26107/RBZ-2021-0011 |
|
publication LSID |
lsid:zoobank.org:pub:85FF7873-DCCB-4D89-997D-DE63AE1929A9 |
|
persistent identifier |
https://treatment.plazi.org/id/A1B13FBD-8EE4-4392-9E41-C446BB05DC61 |
|
taxon LSID |
lsid:zoobank.org:act:A1B13FBD-8EE4-4392-9E41-C446BB05DC61 |
|
treatment provided by |
Diego |
|
scientific name |
Hyriopsis panhai |
| status |
sp. nov. |
Hyriopsis panhai View in CoL , new species
( Figs. 1 View Fig , 2C, D View Fig , 3C, D View Fig , 4 View Fig )
Hyriopsis ( Hyriopsis) bialatus View in CoL – Brandt, 1974: 272, 273, pl. 21, fig. 36. (in part).
Hyriopsis View in CoL undescribed Pfeiffer et. al., 2021: 450, 457, 458, fig. 35.
Type material. Holotype (length 104.83 mm, height without wing 38.58 mm, width 20.73 mm) ( MUMNH: UNI0201 View Materials ) Bang Ta Khian , Song Phi Nong, Suphan Buri, Thailand, 14°10′49.5″N 100°06′07.2″E GoogleMaps ; Paratypes 3 shells (length 85.63–121.99 mm, height without wing 33.18–46.37 mm, width 17.09–26.30 mm): 1 shell ( MUMNH: UNI1210 View Materials ) Bueng Boraphet , Mueang, Nakhon Sawan, Thailand, 15°42′17.8″N 100°09′50.6″E GoogleMaps . 2 shells ( MUMNH: UNI2494 View Materials , 2495 View Materials ) Huai San River in Bang Khian , Chum Saeng, Nakhon Sawan, Thailand ( 15°53′47.3″N 100°13′09.2″E) GoogleMaps .
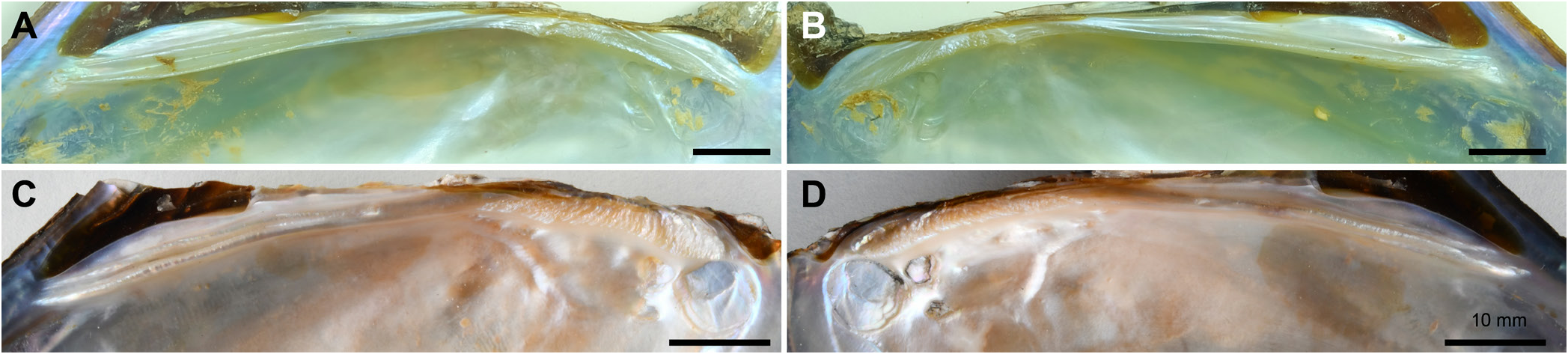
Diagnosis. Shell winged, narrow, elongate and tapering towards the posterior end. Posterior ridges high and obtuse. Ventral margin almost straight. This new species differs from H. bialata in having a more elongated and rounded posterior margin (vs. obtused and somewhat biangulated posterior end), and a distinct separation of anterior adductor muscle scar from pedal retractor muscle scar (vs. a connection between anterior adductor muscle and pedal retractor muscle scars).
Description. Shell moderately thick, winged, compressed and elongate lanciform. Posterior wing prominent and triangular; anterior wing small and pointed. Dorsal margin somewhat curved; umbonal area low. Posterior ridges high and obtuse, running from umbo to posterior end, and with two secondary faint ridges dorsally. Anterior margin round and curved towards anterior wing; posterior margin elongated and rounded. Ventral margin almost straight and slightly curved upward posteriorly. Periostracum greenish to brownish or blackish. Shell surface with fine growth lines. Ligament strong and covered by wing. Pseudocardinal teeth crenulated or vertically striated; one in left valve, two in right; outer tooth in right valve subobsolete. Lateral teeth long and straight or slightly curved; two in left valve, one in right. Anterior adductor muscle scar round, deep and distinctly separated from pedal retractor muscle scar. Posterior adductor muscle scars very shallow. Excurrent aperture smooth, shorter than incurrent. Incurrent with 1–2 rows of short, conically shaped papillae. Small epithelial fold divides excurrent and incurrent aperture. Gills elongated and slightly ribbed. Anterior margin of inner gills slightly longer and wider than that of inner gills. Ectobranchous brooding. Glochidia unknown.
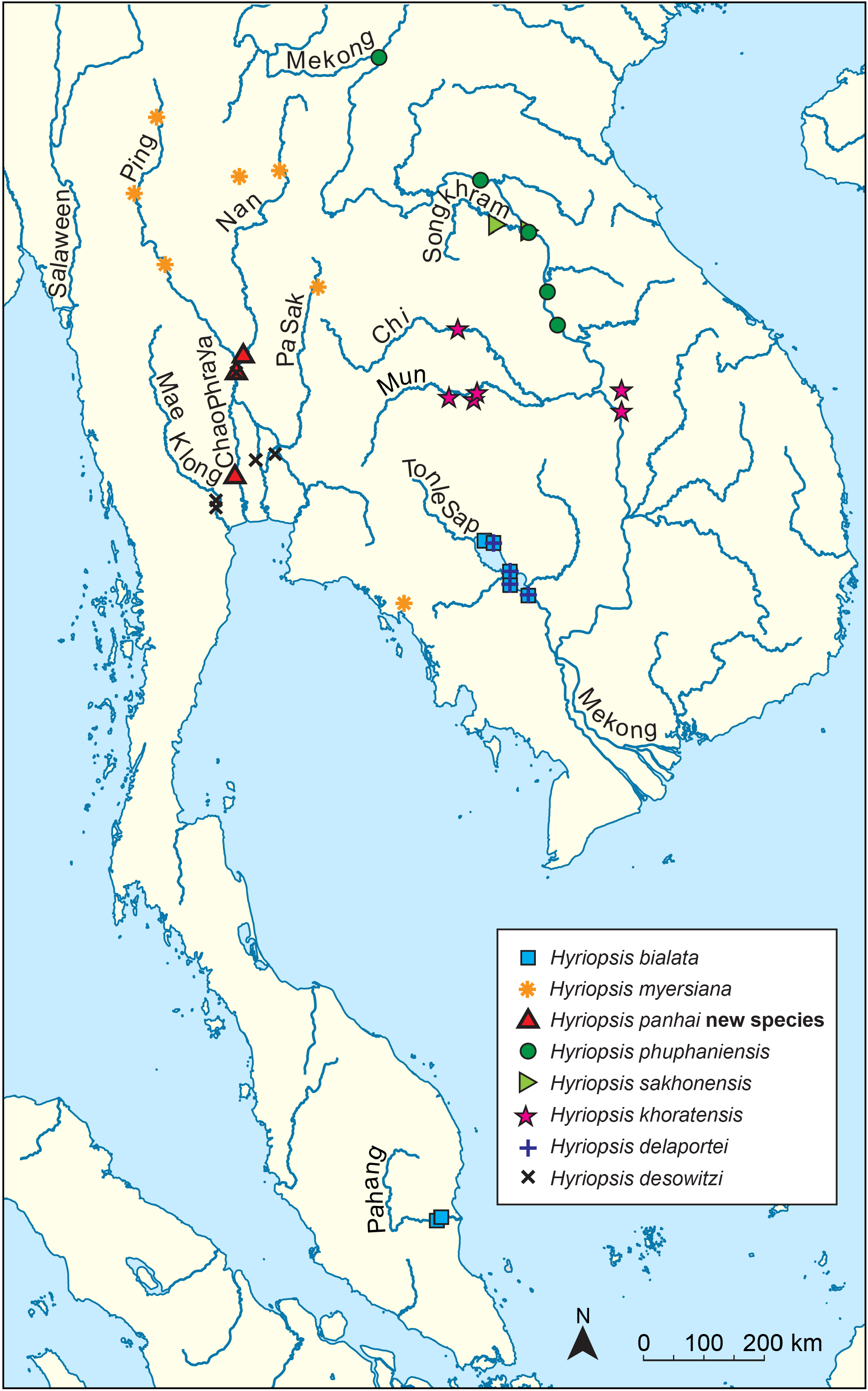
Distribution. This new species is restricted to the Chao Phraya Basin in central Thailand. Its distribution probably extends to the northern part of the basin in Pitsanulok Province based on the record by Brandt (1974).
Habitat. Specimens were collected from muddy and/or sandy substrates. They were found in sympatry with H. desowitzi , Ensidens ingallsianus ( Lea, 1852) , Pilsbryoconcha exilis ( Lea, 1838) , Lens contradens ( Lea, 1838) , and Corbicula spp.
Etymology. This species is dedicated to Prof. Dr. Somsak Panha, a famous Thai zoologist, who has made great contributions to the systematics of invertebrate taxa in Thailand.
Remarks. Hyriopsis bialata recorded by Brandt (1974) as collected from central Thailand and the figured specimen (pl. 21, fig. 36) should be recognised as this new species.
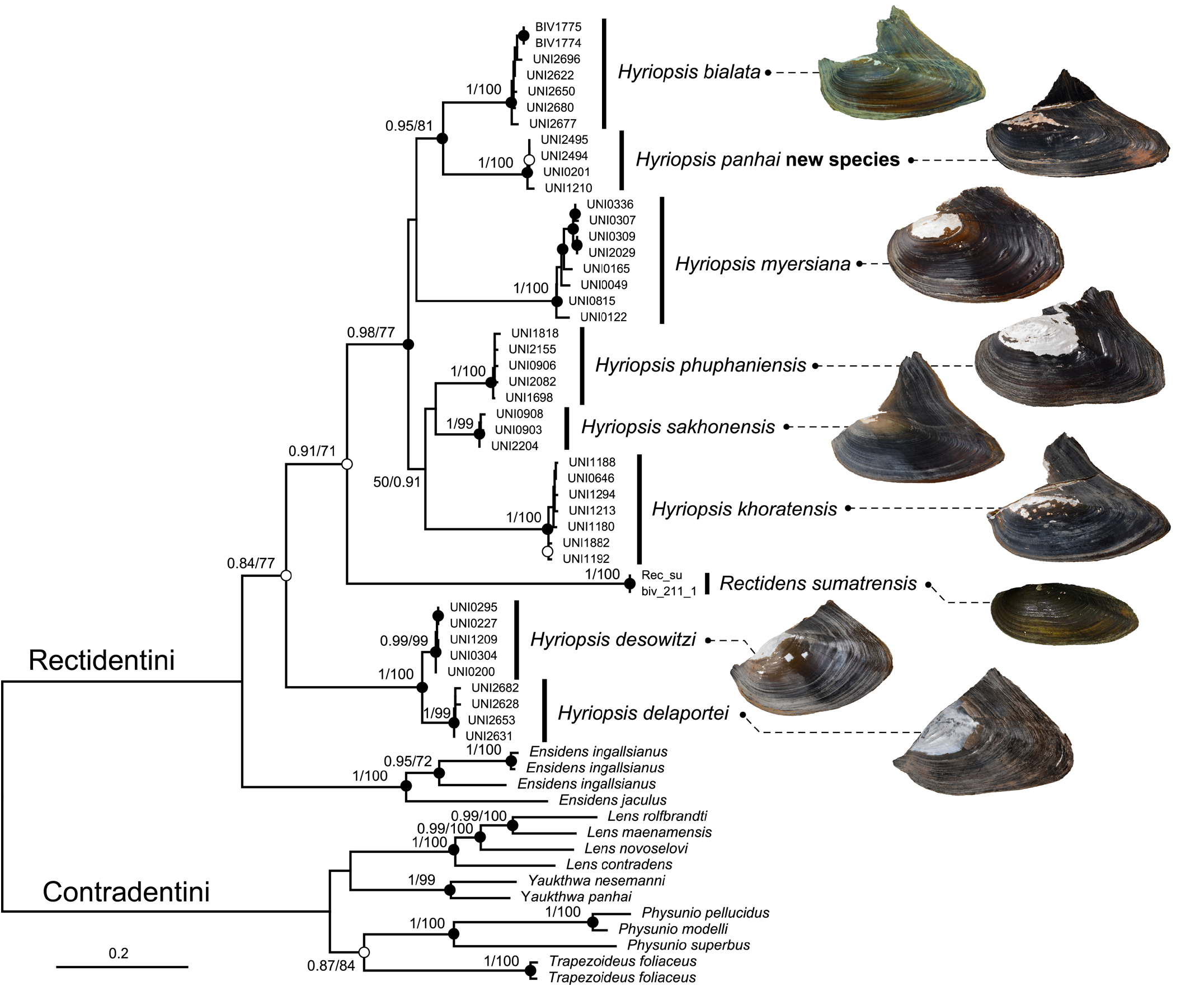
Molecular phylogenies and genetic divergence. The programme PartitionFinder2 suggested dividing the dataset into five partitions, consisting of partitions for 16S and 28S genes, and for each of the three codon positions of the COI gene. The best-fit models for each partition were GTR+I+G for the first codon position of COI, 16S, and 28S; GTR+I for the second codon position of COI; and GTR+G for the third codon position of COI. Multi-locus phylogenies as constructed by Bayesian inference (BI) and maximum likelihood (ML) gave similar topology and congruently recovered Hyriopsis as paraphyletic, consisting of two well-supported clades (77% and 100% bootstrap support for ML, and 0.98 and 1 pp for BI; Fig. 4 View Fig ). The first clade contained the majority of Hyriopsis members, consisting of H. bialata (the type species of the genus), H. myersiana , H. phuphaniensis , H. sakhonensis , H. khoratensis , and the new species H. panhai . All of these taxa were each recovered as highly supported monophyletic clades (99–100% bootstrap support for ML and 1 pp for BI) and separated from each other by high mean uncorrected COI p-distances (6.15–9.92%; Table 2). However, the phylogenetic relationships among them were unclear. Only a sister relationship between H. bialata and H. panhai was highly supported (81% bootstrap support for ML and 0.95 pp for BI), while three taxa from Mekong River Basin, H. phuphaniensis , H. sakhonensis , and H. khoratensis ( Fig. 2 View Fig ), were nested together with nonsignificant support (50% bootstrap support for ML and 0.91 pp for BI). The genetic divergence between H. bialata and H. panhai was 8.30% uncorrected COI p-distance. Another clade of Hyriopsis consisted of two species, H. desowitzi and H. delaportei . These two species are genetically different by 3.21% uncorrected COI p-distance. The paraphyly of Hyriopsis is separated by the Sundaland genus Rectidens , but was only moderately supported by ML analysis (71–77% bootstrap support for ML).
.
)
diagonal 11 / N
A below 10 ± 0.60 0.10 ±
( 9 0.10 1.23 4.69 ± ± 1.39 9.21 ±
tribe 0.10 11.715 13.36 the in 0.14 0.66 1.13 1.38 species 8 ± ± ± ± 0.25 3.21 10.886 12.83 between 0.00 1.29 1.32 1.32 1.33 sequences 7 ± 0.00 ± 11.12 11.93 ± ± 12.90 12.73 ± fragment 6 0.19 ± ± 1.31 1.22 ± ± 1.26 1.22 ± 1.39 ± gene 0.34 12.87 11.01 11.34 11.71 13.11
COI bp 5 0.19 ± ± 1.06 1.28 ± ± 1.16 ± 1.27 1.09 ± ± 1.30
660 0.54 8.09 11.78 9.76 10.50 9.94 12.28 the for matrix 4 ± 0.19 0.87 ± 1.02 ± 1.26 ± ± 1.17 1.22 ± ± 1.14 1.23 ±)
E
. 0.65 6.15 7.85 11.19 9.55 10.50 10.145 11.99
.
S
±
%: 0.30 1.07 1.08 1.14 1.32 1.25 1.30 1.19 1.23
p-distance bold.
3 1.46 ± ± 8.84 ± 8.26 9.92 ± 13.09 ± 10.59 ± ± 11.35 ± 11.66 12.37 ± in
uncorrected shown are 2 ± 0.08 0.08 1.07 8.54 ± 7.24 ± 1.01 8.31 ± 1.08 1.19 ± 9.52 11.43 1.33 ± 1.23 9.69 ± 1.22 ± 9.63 ± 1.14 10.159 ± 1.22 10.93 (
divergence each taxon 1 0.21 ± 1.10 ± 1.05 ± ± 1.00 ± 0.96 ± 1.05 ± 1.32 1.20 ± ± 1.26 1.13 ± 1.21 ± genetic within 0.57 8.30 8.39 7.43 6.93 8.12 12.87 10.32 11.05 10.66 11.53 interspecific distances intraspecific bialata species new, panhai myersiana khoratensis phuphaniensis sakhonensis sumatrensis desowitzi delaportei ingallsianus jaculus Average
2 Table
.
Average Taxa. 1 Hyriopsis Hyriopsis . 2 Hyriopsis . 3 4 Hyriopsis . 5 Hyriopsis . Hyriopsis . 6 7. Rectidens Hyriopsis . 8 Hyriopsis 9. 10 Ensidens .. 11 Ensidens

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.