
Allosauroidea, Currie & Zhao, 1994
|
publication ID |
https://doi.org/10.5252/geodiversitas2022v44a12 |
|
publication LSID |
urn:lsid:zoobank.org:pub:231E3A73-1800-4559-A995-013482DEA623 |
|
DOI |
https://doi.org/10.5281/zenodo.6399034 |
|
persistent identifier |
https://treatment.plazi.org/id/03F29C74-FFEB-960A-D66E-6CB1FCDC9BE4 |
|
treatment provided by |
Felipe |
|
scientific name |
Allosauroidea |
| status |
|
Indeterminate Allosauroidea
A1, MPV 2020.1.1, B3, MPV 2020.1.9, are interpreted as belonging to indeterminate Allosauroidea . This assignment is discussed here.
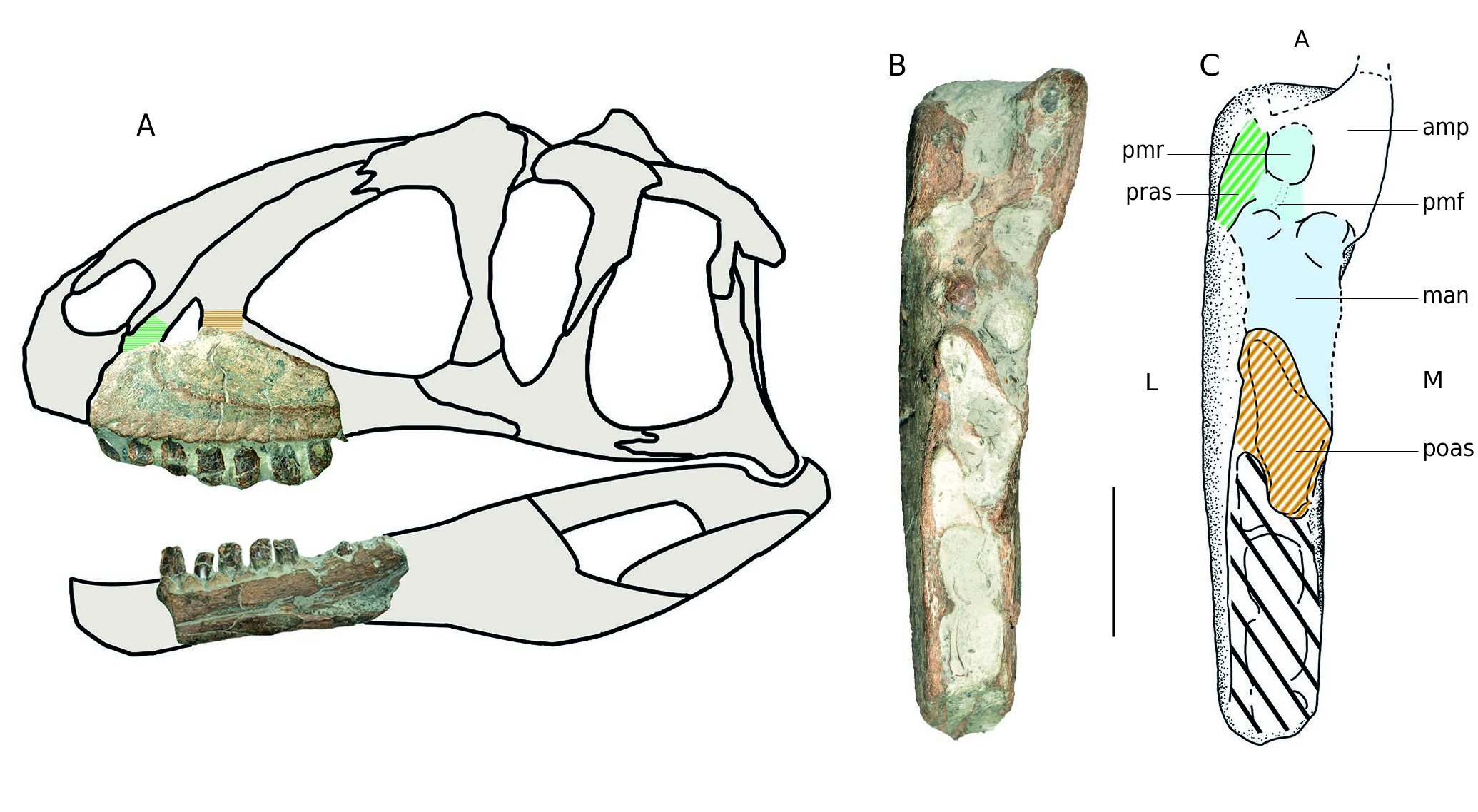
Regarding morphological similarities between the two maxillae A1 and MPV 2020.1.1, it is likely that they belong to closely related taxa. The presence of a maxillary fenestra (see Fig. 9 View FIG ) is informative to differentiate megalosauroids from allosauroids. Indeed megalosauroid like Eustreptospondylus or Dubreuillosaurus possess a maxillary fossa (medially closed maxillary fenestra sensu Allain (2002) and Rauhut & Pol (2019) here considered as maxillary fossa) in contrast to more derived Tetanurae ( Allosauroidea and Coelurosauria) ( Allain 2002; Carrano et al. 2012; Hendrickx & Mateus 2014; Rauhut & Pol 2019). A1, with a maxillary fenestra and hence MPV 2020.1.1, appear closer to allosauroids like Allosaurus or Sinraptor ( Madsen 1976; Currie & Zhao 1994; Chure & Loewen 2020). It is then relevant to assign the specimens to an indeterminate Allosauroidea . In addition to this, following Carrano et al. (2012), the peculiar short anterior ramus similar to Sinraptor would allow to assign the maxillae to a metriacanthosaurid.
The description and placement of the cervical B3 and the cervical/dorsal MPV 2020.1.9 was especially helped and supported by comparisons with Allosaurus and Sinraptor ( Madsen 1976; Currie & Zhao 1994). The two vertebrae are highly pneumatized and present an organization of laminae similar to that of the two above-mentioned allosauroids. In contrast to megalosauroid, the centra are rather short. Yet, Torvosaurus is an exception with short and well pneumatized vertebrae. However, the rim around the anterior articular facet is absent in these specimens although clearly marked in Torvosaurus and megalosauroids ( Britt 1991; Carrano et al. 2012) Furthermore, the highly pneumatized nature of these two vertebrae is more closely comparable to Sinraptor . Indeed, in Allosaurus , pleurocoels are relatively smaller than in Sinraptor and the specimens described here. ( Madsen 1976). This seems appropriate considering the Oxfordian age of Sinraptor ( Currie & Zhao 1994) and Metriacanthosaurus Walker, 1964 , another metriacanthosaurid, which is stratigraphically and geographically close from above specimens. Finally, considering their dimensions and similarities, it is possible that these two bones belong to the same taxon or even the same individual.
MINIMAL NUMBER OF TAXA AND INDIVIDUALS
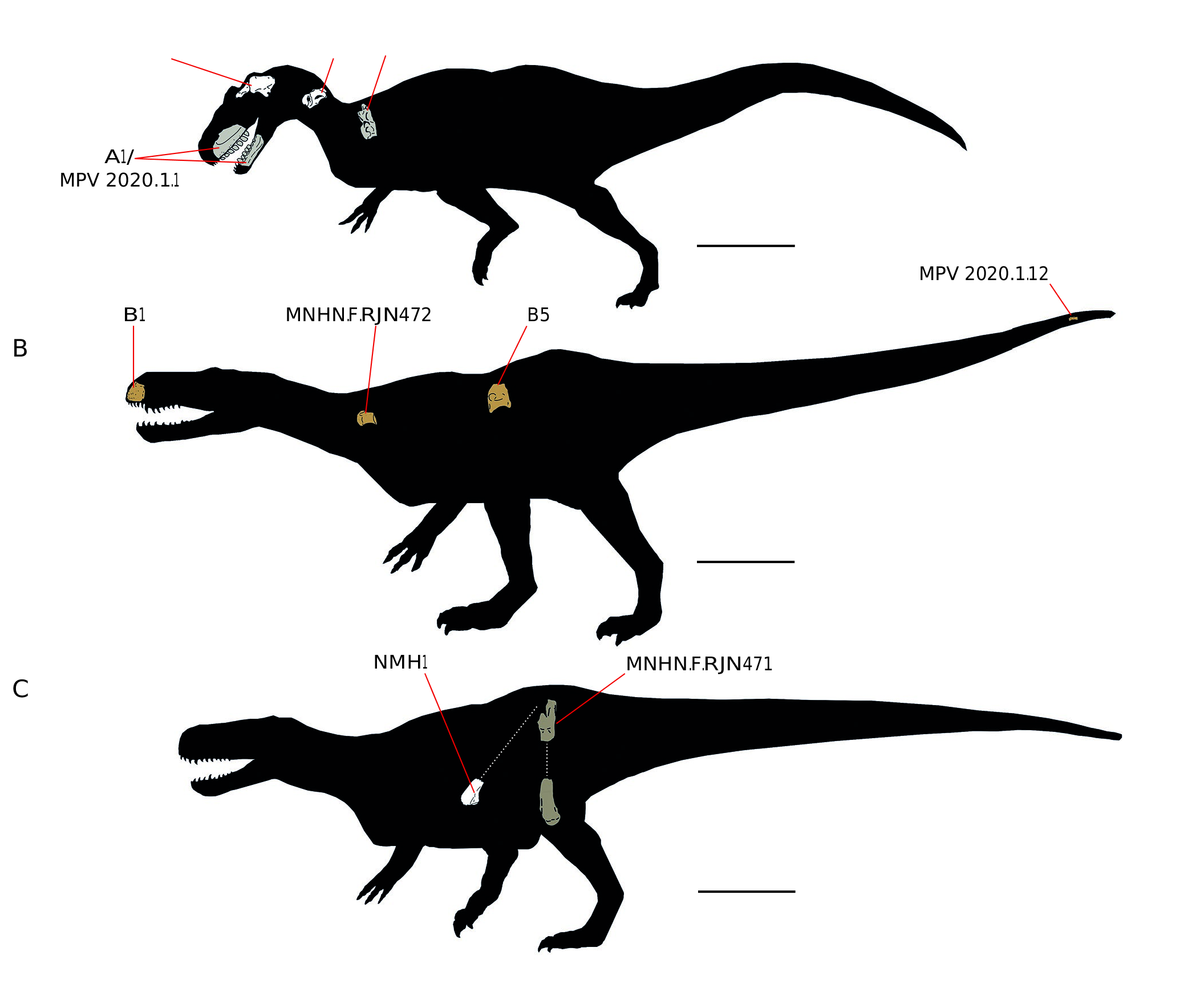
The study of 26 specimens allows to determine that, at least, three different taxa were found in the Vaches Noires cliffs. The first is an indeterminate Allosauroidea ( Fig. 19A View FIG ) with a minimum of two individuals: one being represented by the A 1 specimen and the other by the maxillary fragment MPV 2020.1.1. To these allosauroids can be added the vertebrae B3 and MPV 2020.1.9 and the reconstructed braincase APVSM1+B25392 ( Fig. 5 View FIG ). The second taxon is Streptospondylus, represented by potentially 4 individuals: one is the type specimen to which the vertebra MPV 2020.1.10 and the femur of the Pezy collection (EM pers. obs) may be associated. The second is represented by the cervical vertebra MPV 2020.1.11. Then, the tibia MPV 2020.1.2 referred to Streptospondylus is smaller than the type specimen and indicates a third individual, the size of Eustreptospondylus oxoniensis OUM J. 13558. The last is a large adult from the Oolithe Ferrugineuse de Villers ( Fig. 19B View FIG ). This one is based on specimens B1, B5, MPV 2020.1.12 and MNHN.F.RJN472 and could be the single theropod known from this formation. The last taxon is an indeterminate Megalosauridae with two large individuals represented by femora ( Fig. 19C View FIG ): one is NMH1 and the other is MNHN.F.RJN471 from H14 level of the Marnes de Villers. However, the distinction between this taxon and Streptospondylus is subject to caution because it is based on the femur MNHN.F.RJN93, which is a referred specimen of Streptospondylus ( Allain 2001). Nevertheless, these two types of femur still indicate two different megalosauroids. The rest of the bones from the Callovian and Oxfordian marls without more detailed taxonomic assignments are mostly vertebrae. None of their stratigraphic origin and size seem to increase the MNI. Thus, it has to be noticed that these specimens of indeterminate Tetanurae as well as other bones that were not considered in this study (i.e the allosaurid metatarsal described by Buffetaut et al. [2010]) might be parts of the above-mentioned individuals. Likewise, Piveteausaurus , represented by a unique braincase is problematic. First, it has received various taxonomic assignments: Allosauroidea following Allain (2001) or Megalosauroidea in Carrano et al. (2012). Moreover, a braincase does not increase the MNI in terms of anatomical concordance with the whole studied specimens which are mainly axial elements. It is the same for its stratigraphic origin, which is unknown. Thus, it is also possible that it belongs to one of the above individuals or might even be a braincase of Streptospondylus, as already thought by Piveteau (1923) and Walker (1964), and although this braincase is certainly of a much bigger animal than the type of Streptospondylus. A new study of Piveteausaurus in order to clarify its taxonomic affinities is thus essential and in progress (Monvoisin et al. pers comm.).
With a minimum number approach, there are three theropod taxa at the Vaches Noires cliffs and eight individuals ( Table 3 View TABLE ). Many of them come from the lower Oxfordian (Marnes de Villers, Oolithe Ferrugineuse de Villers and Argiles à Lopha gregarea ) instead of the traditionally envisioned upper Callovian (Marnes de Dives). Moreover, they are based on several observations that are subject to bias especially for taxonomic assignments.Nevertheless, the information gathered from these bones indicates an overestimation of the number of theropod individuals, leading to an over-representation that does not reflect the real composition of an ecosystem.
NEW INSIGHTS ABOUT THE THEROPOD OVER- REPRESENTATION OF THE DEPOSIT
As mentioned before, most of the dinosaur remains from the Vaches Noires cliffs belong to theropods. Only very scarce remains of other groups have been found. A caudal vertebra from the Pezy collection (EM pers.obs) represents the second bone referred to a sauropod but also the only one that is left because the first, a vertebra mentioned by Bigot in 1895, was destroyed during World War II ( Bigot 1945; Buffetaut 2013; Plasse & Buffetaut 2016). The recently described thyreophoran vertebrae (probably from a stegosaur) add another clade to the Vaches Noires Cliffs dinosaurs ( Buffetaut & Tabouelle 2019). However, theropod remains are still over-represented. Most of these remains belong to large carnivorous dinosaurs that represent obvious apex predators. It is difficult to imagine an ecosystem mainly composed of predators. Indeed, it is usually accepted that predators are less abundant than their prey ( Farlow 1993; Naish et al. 2004; Läng et al. 2013). Overabundance of theropods found in other deposits is often explained by an environment that is not suitable to the development of a sufficient flora to feed herbivores ( Naish et al. 2004; Läng et al. 2013). However, studies of the palaeoflora of the Armorican massif during the Bathonian to Oxfordian show that coastal forests were present and thus allowed the development of a herbivorous population ( Le Couls et al. 2016; Le Couls 2017). Therefore, the Vaches Noires Cliffs assemblage obviously does not faithfully reflect the ecosystem from which the animals are derived. Noteworthy the marine taphocoenosis bring a consequent bias to reconstruct a terrestrial ecosystem. However, in other similar marine deposits, like the Callovian clay pit at Argences (Calvados, Normandy), no theropod remains were found, but a sauropod ( Buffetaut et al. 1995) and stegosaurs ( Hoffstetter & Brun 1956; Galton et al. 1980; Galton 1990; Allain & Suberbiola 2003) ( Buffetaut & Tabouelle 2019) have been reported. Still in Normandy, but from the Kimmeridgian, similar marine deposits, have yielded a more balanced fauna with theropods, sauropods, stegosaurs and ornithopods. ( Buffetaut 1994a; Buffetaut & Cacheleux 1997; Allain & Suberbiola 2003). As in the Vaches Noires cliffs, the Oxford Clay Formation of England includes both Callovian and Oxfordian marls. However, the preserved fauna is more diverse, with many dinosaur groups ( Martill 1988; Lomax & Tamura 2014; Buffetaut & Tabouelle 2019). Thus, the over-representation of theropods at the Vaches Noires is enigmatic and many hypotheses have been put forward to explain it. It is possible that theropods frequented coastlines to feed on beached carcasses, which made them more likely to end up in the sea. Piscivorous tendencies among theropods have been demonstrated by geochemical studies, notably in spinosaurid megalosauroids ( Amiot et al. 2010; Brusatte 2012; Allain et al. 2012). Other megalosauroids were likely to feed on fish, as shown by the gastric remains of Poekilopleuron Eudes-Deslongchamps, 1838 ( Allain 2005). This hypothesis is also supported by the fact that megalosauroids may have preferred nearshore environments as suggested by the study of theropod paleogeographic distribution of Rauhut et al. (2016). Moreover, theropod swimming tracks have been found, supporting occasionally aquatic behaviour ( Milner et al. 2006; Ezquerra et al. 2007). However, these tracks are from freshwater environments. Nevertheless, a possible nearshore palaeobehaviour is not sufficient to explain the over-representation, because, as explained before, other marine deposits and especially the Oxford Clay present a more balanced fauna. Finally, a statistical effect is more likely ( Buffetaut &Tabouelle 2019). Indeed, the Vaches Noires cliffs are a 5 km long costal outcrop which has yielded only a few dinosaurs remains so that it is difficult to compare it with a much more extensive geological formation such as the Oxford Clay. The over-representation may be due mainly to the small sample available from the Vaches Noires and may not represent the real ecosystem of nearby islands (i.e Armorican massif).
Another peculiar aspect of the Vaches Noires cliffs is the way fossils are found, especially concerning dinosaur bones from marls situated above the Marnes de Dives, which crop out at the bottom of the cliffs. These remains, usually collected on the beach at the bottom of the cliffs, come from marl flows, the result of which is a huge mixing of the different fossilbearing layers. Only index fossils or a peculiar matrix can help to identify the exact origin of specimens. It was traditionally argued that dinosaur remains from the Vaches Noires cliffs are scattered because of the long time they have spent floating in the sea, during which they decomposed before dropping to the bottom ( Buffetaut 1994a, 2013). However, partial to quite complete skeletons have been found in other similar deposits. It is the case with Eustreptospondylus or other Oxford Clay dinosaurs ( Martill 1988; Lomax & Tamura 2014) or even the Lexovisaurus from the clay pit of Argences ( Hoffstetter & Brun 1956; Galton et al. 1980; Galton 1990). As a matter of fact, it is likely that specimens from fossiliferous layers in cliffs that flow to the foreshore are originally more complete but dissociated by natural erosion. Then those bones are collected at various times by various people so that they are kept apart. This contributes to the over-representation of theropods, insofar as it inflates the sample size. This effect is demonstrated here with the MNI of 8 individuals. Although not absolute, it demonstrates the probable inflation of the number of theropods yielded by the Vaches Noires cliffs. An explicit example are the bones from the same individual found in different collections (i.e APVSM1 frontals with B25392 braincase). Another is the history of the type specimen of Streptospondylus altdorfensis. The bones of that theropod were gathered at different times and not mentioned in the same article although they came from the same skeleton ( Allain 2001). Moreover, a new succession of vertebrae, in direct connection with previously known ones has been recovered (RA, unpublished data).
THE CONTRIBUTION OF THE FOSSILS FROM THE VACHES NOIRES TO OUR KNOWLEDGE OF THEROPODS
The study of unpublished specimens has provided new insights about theropod diversity at the Vaches Noires. For example, Streptospondylus, an iconic dinosaur from that locality is now better known, since it is now known from the Oxfordian. The highly pyritised type specimen rather indicates more Callovian provenance, but as mentioned before, an unambiguous dating is not possible. Thus, with newly referred specimens of this study, Streptospondylus is known from the Oxfordian and possibly the Callovian. Based on our observations, it seems that this theropod, hitherto know from a juvenile ( Allain 2001) could reach a very large size, when adult, in the size range of Torvosaurus . The probably close relationships between Streptospondylus and Eustreptospondylus are once again highlighted. A comparative study of these two taxa is highly needed and could provide new insights about the validity of their division into two genera, especially if this distinction is in fact based on different ontogenic stages. Moreover, there are similarities between Jurassic faunas from Normandy and England, leading to taxonomic revision. This is for example the case with the stegosaur genus Lexovisaurus Hoffstetter, 1957 , present in the English and French Callovian. It is currently considered to be a nomen dubium and replaced by Loricatosaurus Maidment, Norman, Barrett & Upchurch, 2008 ( Maidment et al. 2008), but this suggestion is debated ( Buffetaut & Morel 2009). The supposed «source» of dinosaurs from these two marine deposits are considered as distinct: London-Brabant Massif for the Oxford Clay and Armorican Massif for the Vaches Noires cliffs and other localities in Normandy ( Buffetaut 2013), both representing Jurassic insular ecosystems from the European Archipelago ( Bradshaw et al. 1992).
Despite the limited number of remains and their fragmentary nature, the specimens from the Vaches Noires cliffs have scientific value. They represent remains from a peculiar evolutionary context for theropods. During the Middle to Late Jurassic a faunal transition is observed with a faunal turnover from megalosauroids dominated faunas to allosauroids ones (avetheropods in a broad sense) ( Benson 2010; Rauhut et al. 2016). Although it may provide from an artifact of preservation according to the same authors ( Rauhut et al. 2016, 2018), this effect is also visible in time calibrated trees of recent theropod phylogenies ( Carrano et al. 2012; Hendrickx et al. 2015; Rauhut et al. 2016). The presence of both groups in the Vaches Noires cliffs is therefore informative. Megalosaurids are represented by large size predators and some appear to be close relative to megalosaurines. It is noteworthy that after the Callovian, megalosaurids yet quite common in Europe ( Rauhut et al. 2016, 2018) were hitherto represented by the single genus Torvosaurus , known in North America and Portugal during the Kimmeridgian and Tithonian ( Britt 1991; Hendrickx & Mateus 2014; Rauhut et al. 2016). The studied remains found as close relative to Torvosaurus , as well as the Callovian megalosaurid Wiehenvenator albati Rauhut, Hübner & Lanser, 2016, also close to the genus according to Rauhut et al. (2016), suggest the establishment and wide distribution of megalosaurines theropod, surviving through the late Jurassic ( Rauhut et al. 2016; Malafaia et al. 2017). However, during this transition period, allosauroids are rather rare, especially in European deposits, the Oxfordian Metriacanthosaurus being the only member known (see Rauhut et al. 2016: table 3). The description of allosauroid remains in the present study provide thus new insights about their presence in the European Archipelago. This is all the more important considering the fragmentary nature of Metriacanthosaurus . Indeed, this theropod is problematic because it has not been fully described. It is clear that the entire material lacks a complete review. Despite this, Metriacanthosaurus is the type genus of the Metriacanthosauridae , a subclade of allosauroids ( Benson 2010; Carrano et al. 2012). Therefore, the status of the family is likely not definitive. In addition to this, the A 1 specimen has similarities with metriacanthosaurids also know in China, with the contemporaneous genus Sinraptor ( Currie & Zhao 1994) . Finally, Allain (2005) and Rauhut et al. (2016) suggest that megalosauroids might have preferred nearshore environments (the origin of the studied bones), whereas allosauroids tend to occur in inland environments. Thus, the rarity of allosauroids in the European Archipelago could be explained by a preferential ecological niche. Finally, finding remains of allosauroids despite the environment supposedly not being their preferred option might reinforce the idea of a faunal transition between megalosauroids and allosauroids.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.