
Allosauroidea, Currie & Zhao, 1994
|
publication ID |
https://doi.org/10.5252/geodiversitas2022v44a12 |
|
publication LSID |
urn:lsid:zoobank.org:pub:231E3A73-1800-4559-A995-013482DEA623 |
|
DOI |
https://doi.org/10.5281/zenodo.6399027 |
|
persistent identifier |
https://treatment.plazi.org/id/03F29C74-FFFF-9604-D141-68D2FB6998A1 |
|
treatment provided by |
Felipe |
|
scientific name |
Allosauroidea |
| status |
|
Allosauroidea indet.
CRANIAL SKELETON
Maxilla and dentary
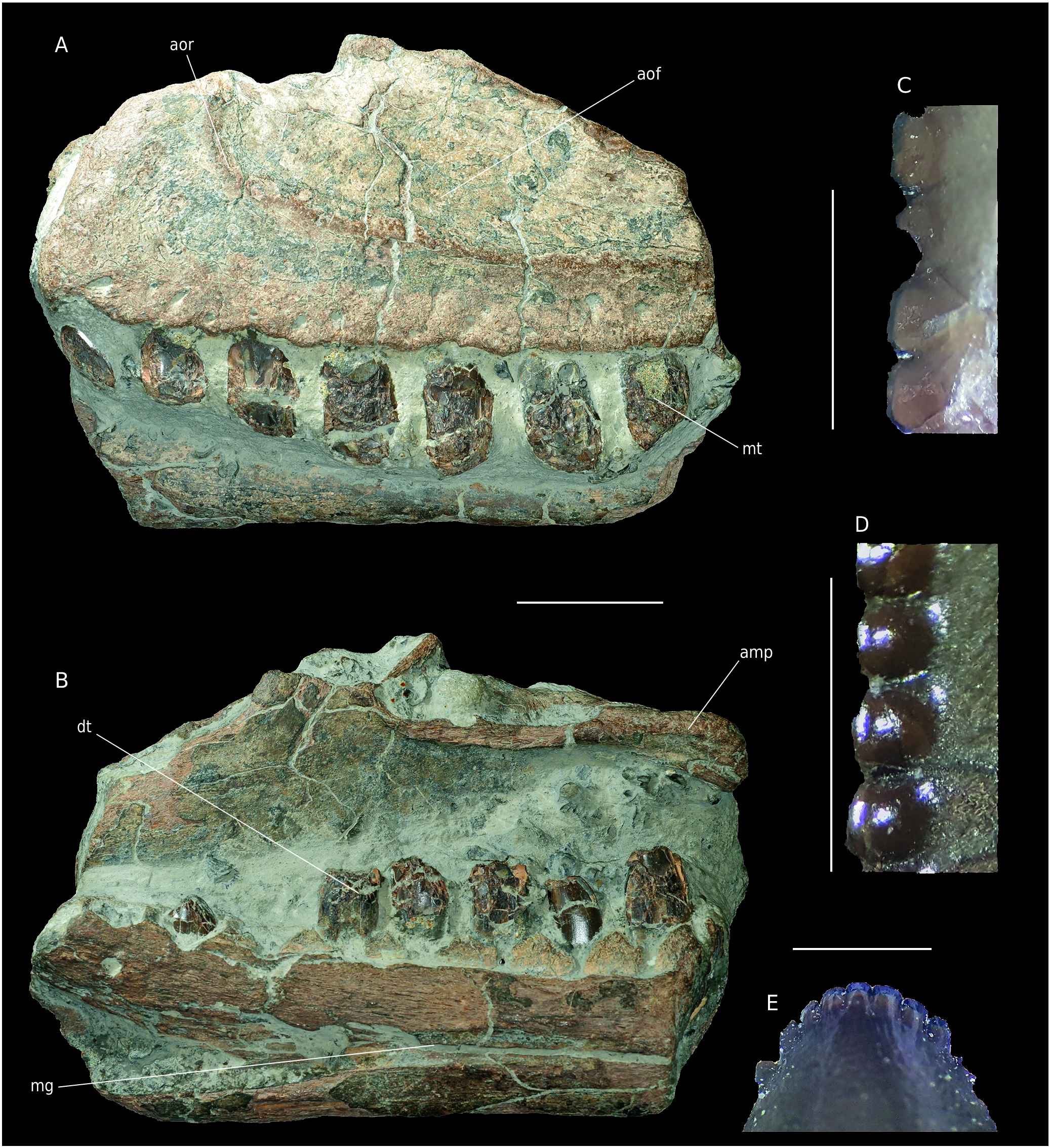
Material. A1 a left maxilla with dentary, MPV 2020.1.1, a left maxilla ( Fig. 8 View FIG ).
Stratigraphy. A1: lower Oxfordian marls according to the A.gregareum shells of the matrix. MPV 2020.1.1: indeterminate between Oxfordian and Callovian marls.
Discovered in 2002 at the bottom of the cliffs by the Anicolas family, the specimen A1 is a fragment of maxilla and dentary in occlusion, held together by matrix.
A1 Maxilla. The bone is 220 mm long and 115 mm high as preserved. Matrix is still present on the medioventral side between the teeth and on the anterior margin (further preparation was not allowed by the owners). As a result, the medioventral part of the specimen is not visible. The bone is obliquely fractured posteriorly and dorsoventrally so that two alveoli are visible in dorsal view. Only the basis of the ascending process is preserved. Fractures are – as often with Vaches Noires cliffs fossils – eroded and water-worn. The teeth are poorly preserved and shattered. As in Sinraptor , the anterior ramus of the maxilla is very short, with a rounded shape ( Fig. 8A View FIG ). In anterior view, the eroded anteromedial process protrudes from the ramus. In medial view, the articular crest and groove of this structure are visible but eroded. A sagittally fractured medial shelf is placed posterior to the anteromedial process. This shelf medially delimits the maxillary antrum located above it. This structure is a cavity at the base of the ascending process delimited anteriorly and posteriorly by preantral and postantral struts respectively. Among Avetheropoda the antrum is laterally and medially open so that it forms a maxillary fenestra ( Fig. 9 View FIG ) ( Carrano et al. 2012; Witmer 1997; Hendrickx & Mateus 2014). Thus, in megalosauroids like Torvosaurus , Eustreptospondylus or Dubreuillosaurus Allain, 2005 , the antrum is medially closed so that they bore a maxillary fossa ( Britt 1991; Allain 2002; Sadleir et al. 2008; Hendrickx & Mateus 2014) in contrast with allosauroids like Allosaurus , Acrocanthosaurus or Sinraptor ( Madsen 1976; Currie & Zhao 1994; Eddy & Clarke 2011; Chure & Loewen 2020). In our specimen the preantral and postantral struts are not preserved but their bases are visible dorsally by differentiating bony surfaces from fractures ( Fig. 9B View FIG ). The basal surface of the promaxillary fenestra is perceptible ahead of the antrum with the promaxillary recess diving into the matrix ( Fig. 9B; C View FIG ). In dorsal view, there are no fractures laterally to the antrum and its surface seems natural. Thus, the structure opens mediolaterally between the antorbital fossa and medial shelf. As a result, it is possible to determine that the specimen bore a maxillary fenestra ( Fig. 9 View FIG ). In lateral view, the antorbital fossa is clearly delimited by a polished rim, the remains of an antorbital crest ( Fig. 8A View FIG ). This fossa strongly invades the maxilla. Below, foramina 5 mm in diameter pierce the ventral margin over its whole length. Seven much fractured teeth are visible, missing their apical parts. On some of them, denticles can be observed ( Fig. 8C View FIG ) but no carina is fully preserved.
Dentary A1. This fragment of dentary ( Fig. 8B View FIG ) is 220 mm long and 70 mm high as preserved. It is broken anteriorly and posteriorly. The main surface of the bone has flaked off so that some parts of the cortical bone are missing. Almost all the lateral surface is covered by the matrix. A longitudinal concavity, posteriorly wide and decreasing to a thin groove (about 2 mm wide) anteriorly is visible on the medial surface. This structure is filled by matrix. It corresponds to the contact area with the splenial bone posteriorly and the Meckelian groove anteriorly. Because of the matrix, it is not possible to see foramina near the groove opening. Interdental plates are unfused. They are rectangular at their base, with a triangular top. This shape is similar to the condition in Sinraptor and specimen MPV 2020.1.1., in which, moreover, the interdental plates are unfused, in contrast with Allosaurus ( Madsen 1976) . To judge from the reduced size of the posterior alveoli and the dorsoventral widening of the bone, the fragment is supposed to be the posterior part of a dentary ( Fig. 9 View FIG ). As for the maxilla, teeth are poorly preserved. Six are erupted in the first five and the eighth alveoli. In the fourth alveolus, the apex of a replacement tooth is visible ( Fig. 8E View FIG ). Because the teeth are splattered, no carina is fully preserved on the dentary teeth, a dental morphology and comparison with other Vaches Noires remains is then not very reliable. Only part of the mesial carina of the second dentary tooth is preserved. It bears sixteen roundedmesial denticles per five millimetres ( Fig. 8D View FIG ). Their shape seems different from those on the maxilla as they are more bulbous and well distinct from the crown ( Fig. 8C, D View FIG ).
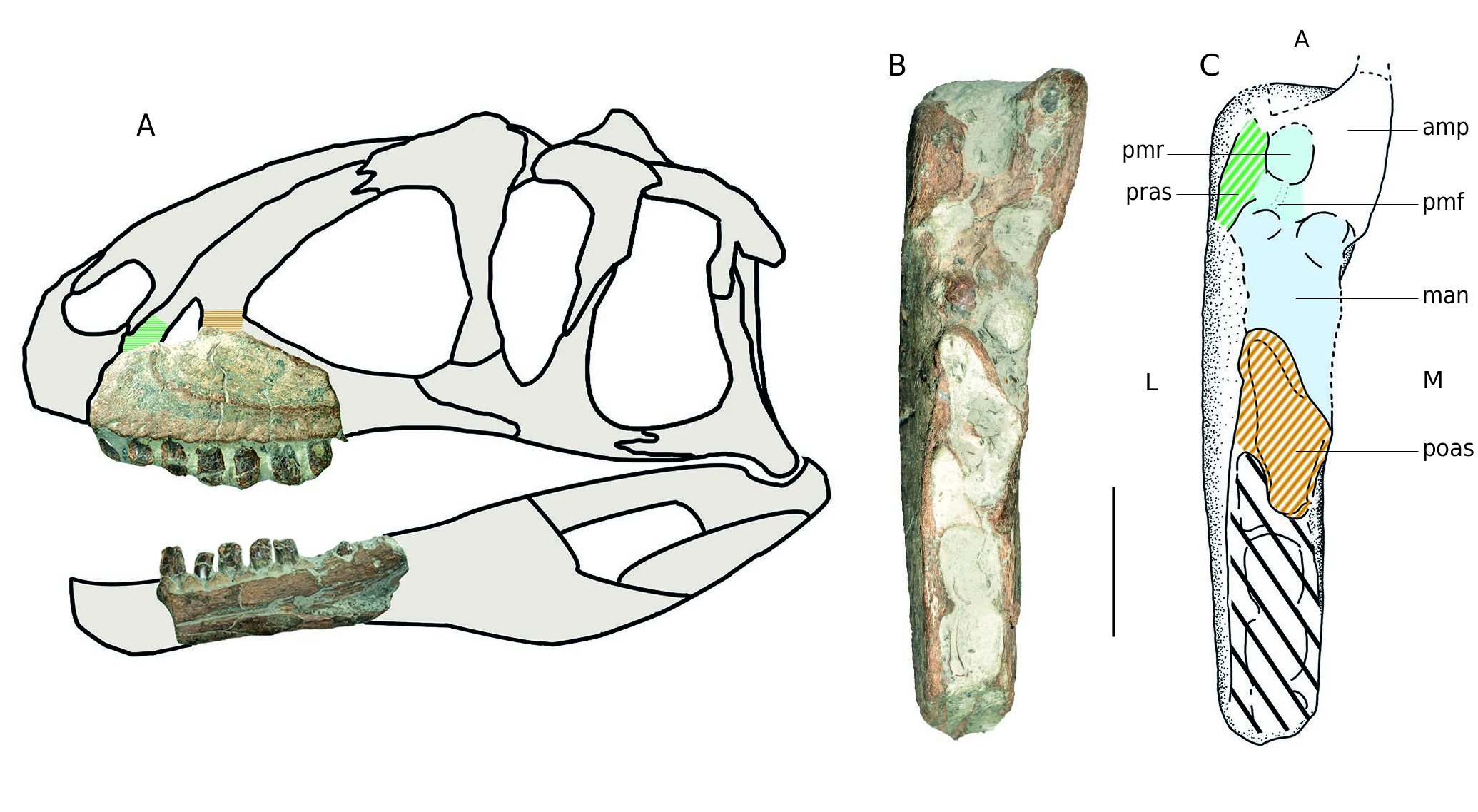
Maxilla MPV 2020.1.1. Already described by Buffetaut et al. (1991), this fragment of a left maxilla from the Pennetier collection was attributed to Megalosaurus sp. The stratigraphic position of this specimen is unclear. Buffetaut et al. (1991) referred it to the Upper Callovian, supposedly torn from the Marnes de Dives by a storm. All the other bones from the Pennetier collection are supposed to come from Oxfordian marls (Pennetier: pers.com). However, because there is no stratigraphic marker on the specimen, the origin of MPV 2020.1.1 must be given as Callovian or Oxfordian marls.
The specimen corresponds to the anterior part of a left maxilla ( Fig. 10 View FIG ) which was originally in contact with the premaxilla. This fragment is characterised by its massive appearance, a deep dorsal depression near the anteromedial process, as well as a large depression underneath it ( Buffetaut et al. 1991). The first corresponds to the promaxillary recess of the promaxillary fenestra, diving into the anterior ramus ( Witmer 1997; Hendrickx & Mateus 2014) ( Fig. 10C View FIG ). This structure is in the anterior part of the ascending process directed towards the nasal. As a result, it had to be placed more anteriorly than in Megalosaurus , Dubreuillosaurus or even Allosaurus ( Madsen 1976; Allain 2002; Benson 2010). Such a configuration, with a remarkably short anterior ramus, is visible in Sinraptor as well as in some carcharodontosaurids such as Acrocanthosaurus ( Eddy & Clarke 2011) and Carcharodontosaurus Stromer, 1931 ( Brusatte & Sereno 2007) but also in specimen A1. The relative proportions of the anterior ramus and the anteromedial process are also similar to the latter specimen, as well as the position of the antorbital fossa. The anteromedial process is not directly dorsal to the interdental plates as in Sinraptor , but rather raised and ventral to the dorsal margin of the anterior ramus as in Allosaurus . Although only the anteroventral corner is preserved, the antorbital fossa is similar to specimen A1 so that the rim is clearly visible, unlike in Megalosaurus . However, MPV 2020.1.1 is much more massive than A1. This may indicate two related taxa or different stages or sexual dimorphism in the same species. These assumptions could be verified if, after cleaning, the large depression under the anteromedial process is also found in A1. As in Sinraptor ( Currie & Zhao 1994) , the interdental plates are nor fused or ornamented, but much more spaced although this may have been emphasized by erosion. The denticles on the teeth of MPV 2020.1.1 are subrectangular and there are about nine per five millimetres ( Fig. 10D View FIG ).
Braincase and frontals
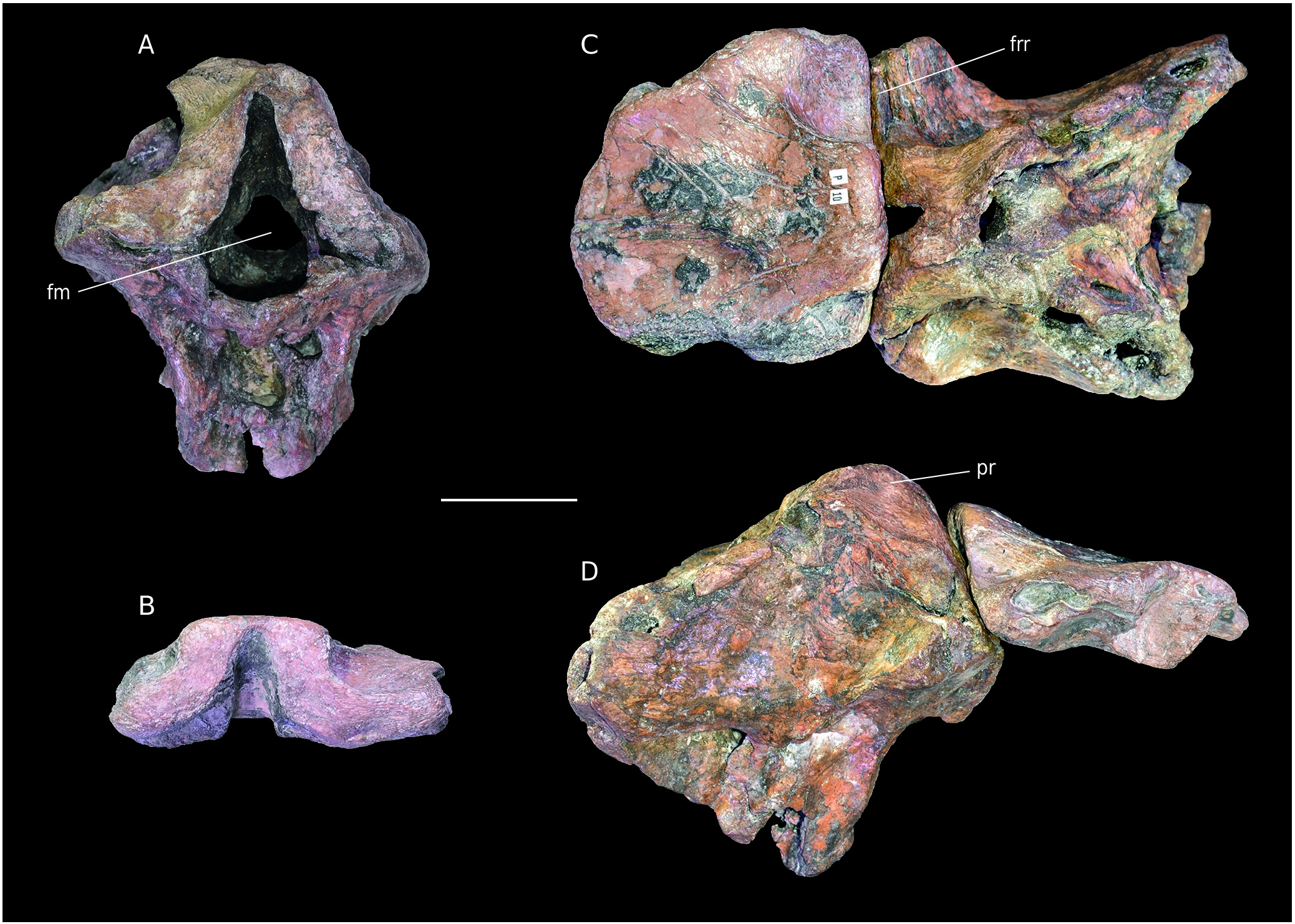
Material. APVSM1 associated frontals, B25392 braincase ( Fig. 11 View FIG ).
Stratigraphy. Callovian or Oxfordian marls.
Both specimens were already described by Buffetaut & Enos 1992 (frontals, Fig. 11B View FIG ) and Knoll et al. 1999 (braincase, Fig. 11A View FIG ). After combining the specimens from the Enos (APVSM1) and Bülow (B25392) collection, it is now clear that these bones belong to the same individual. This was already proposed ( Knoll et al. 1999) and is now confirmed. The bone determined as the sphenethmoid in Knoll et al. (1999) on the braincase coincides with the attachment area on the specimen APVSM1. The supposed sphenethmoid is then here identified as remains of the frontals still fused with the parietal. ( Fig. 11B View FIG ). Therefore, the specimens were separated by a fracture, not by separation along a sutural surface. Differences between these specimens and Piveteausaurus and Eustreptospondylus were already noticed ( Buffetaut & Enos 1992; Knoll et al. 1999). One is the flatness of of the dorsal surface of the frontals, which is not seen in APVSM1. Indeed, in APVSM1, the frontoparietal suture is raised, as in Allosaurus ( Madsen 1976; McClelland 1990). In contrast, a flat dorsal surface of the frontals is visible in megalosauroids such as Eustreptospondylus and Dubreuillosaurus . Moreover, unlike the latter two theropods, but as in Allosaurus , the frontals are short and wide ( Allain 2002). Another distinctive feature is the reduced participation of the supraoccipital to the dorsal margin of the foramen magnum ( Knoll et al. 1999). This character is typical of most Allosauroidea ( Allain 2001, 2002). Because of these previous similarities with Allosaurus and members of the superfamily, braincase B25392 + frontals APVSM1 are interpreted as belonging to an indeterminate allosauroid. This assignment is supported by the similarities with Allosaurus found by the neuroanatomical study of the braincase ( Knoll et al. 1999).
AXIAL SKELETON
Cervical vertebra
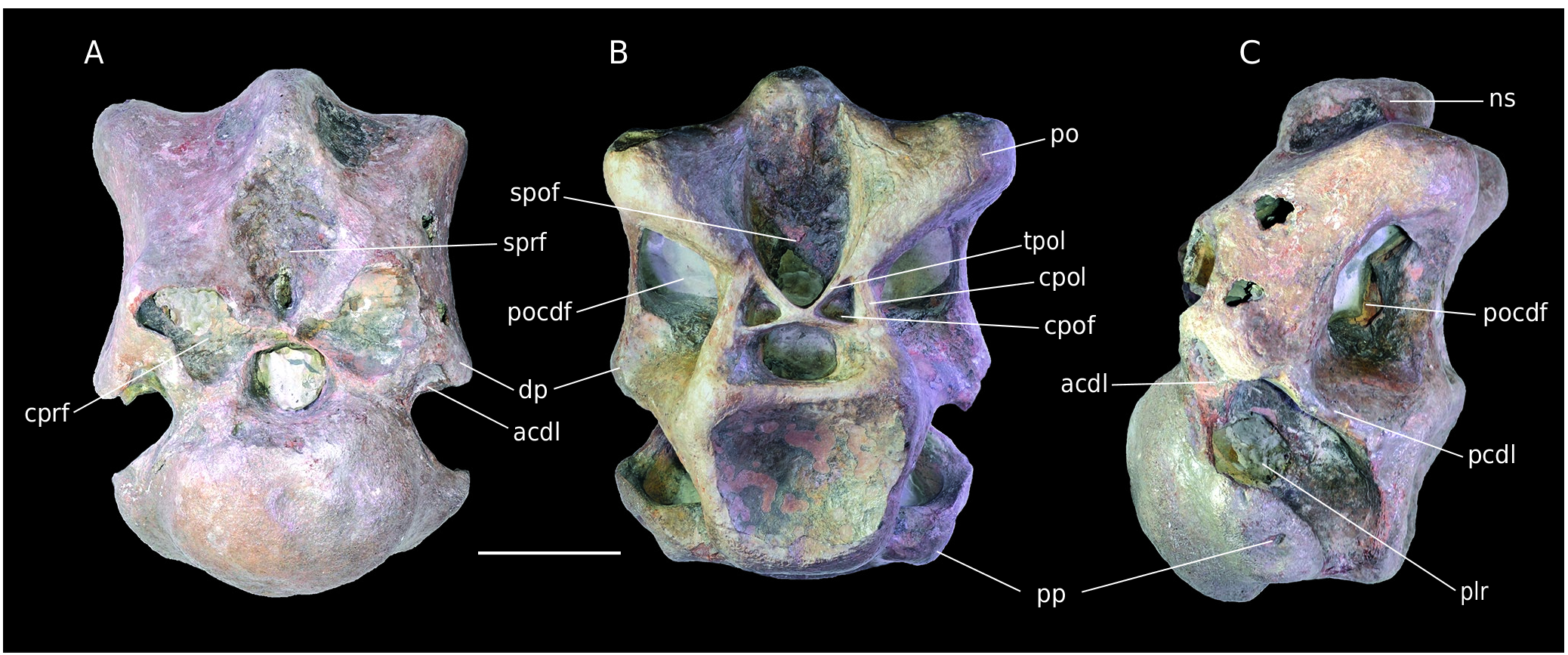
Material. B3, an anterior cervical vertebra ( Fig. 12 View FIG ).
Stratigraphy. Callovian or Oxfordian marls.
B3 is rather well-preserved but water-worn ( Fig. 12A View FIG ). The prezygapophyses and neural spine are broken so that only their bases are visible. The diapophyses are not complete and epipophyses are missing. The posterior articular facet is probably reduced in size because of abrasion of its borders. The deepest parts of the fossae are often hidden by matrix.
The centrum is short and opisthocoelous. The articular facets are wider than high. The vertebra is stocky, as in the Cretaceous theropod Megaraptor namunhuaiquii Novas, 1998 ( Calvo et al. 2004). This contrasts with the strongly opisthocoelous and elongate vertebrae of Eustreptospondylus ( Sadleir et al. 2008) . The specimen has strongly pneumatized centrum and neural arch, much more than in Allosaurus . Pleurocoels expand over the major part of the centrum ( Fig. 12C View FIG ). As in many theropods, they are medially divided into two deep fossae by a bone lamina ( Currie & Zhao 1994). The anterodorsally oriented parapophyses are in the anteroventral corner of the centrum. Their base merges with the ventral margin of the centrum so that they form the anterior half of it. As seen in the fifth cervical of Sinraptor , a depression is present on the ventral face of the centrum. The oval and convex anterior facet is wider than high. The posterior facet is concave and subquadrangular ( Fig. 12B View FIG ). Its peculiar shape is likely accentuated by erosion. This facet is ventrally offset by 40° relative to the anterior one so that the centrum has a flexed appearance ( Fig. 12C View FIG ). Half of the vertebra consists of a stocky neural arch. A rugose spinoprezygapohyseal fossa is present above the neural canal. It is the scar of the interspinous ligament ( Currie & Zhao 1994). In anterior view, fractures on the prezygapophyses make their hollow nature visible. Lateroventral to them there is a triangular fossa formed by the centroprezygapohyseal laminae and anterior centrodiapophyseal laminae. The slightly concave diapophyses are ventrally oriented and laterally triangular. They are supported by the anterior centrodiapophyseal laminae and a strong, nearly horizontal, posterior centrodiapophyseal lamina. Both struts frame the centrodiapophyseal fossa medially and ventrally. As seen in Sinraptor , the posterior centrodiapophyseal laminae also form the ventral margin of a large and deep postzygapophyseal centrodiapophyseal fossa. The postzygapophyses are thick but eroded and waterworn. However, their articular facet is visible and ventrolaterally oriented. The postzygapophyses laterally frame a deep spinopostzygapophyseal fossa. Below the latter, two small symmetrical fossae are present ( Fig. 12C View FIG ). They are surrounded dorsally by the interpostzygapophyseal lamina, laterally by the centropostzygapophyseal lamina and ventrally by the neural canal. They match the centropostzygapophyseal fossae ( Wilson et al. 2011). As in Megaraptor, they are particularly well-defined. The neural spine is worn by erosion and very reduced anteroposteriorly. The bent look of the centrum indicates an “S-shaped” neck of which Allosaurus is a classic example. According to the diapophyses vertical orientation and following Madsen (1976), the specimen is likely a fifth cervical vertebra.
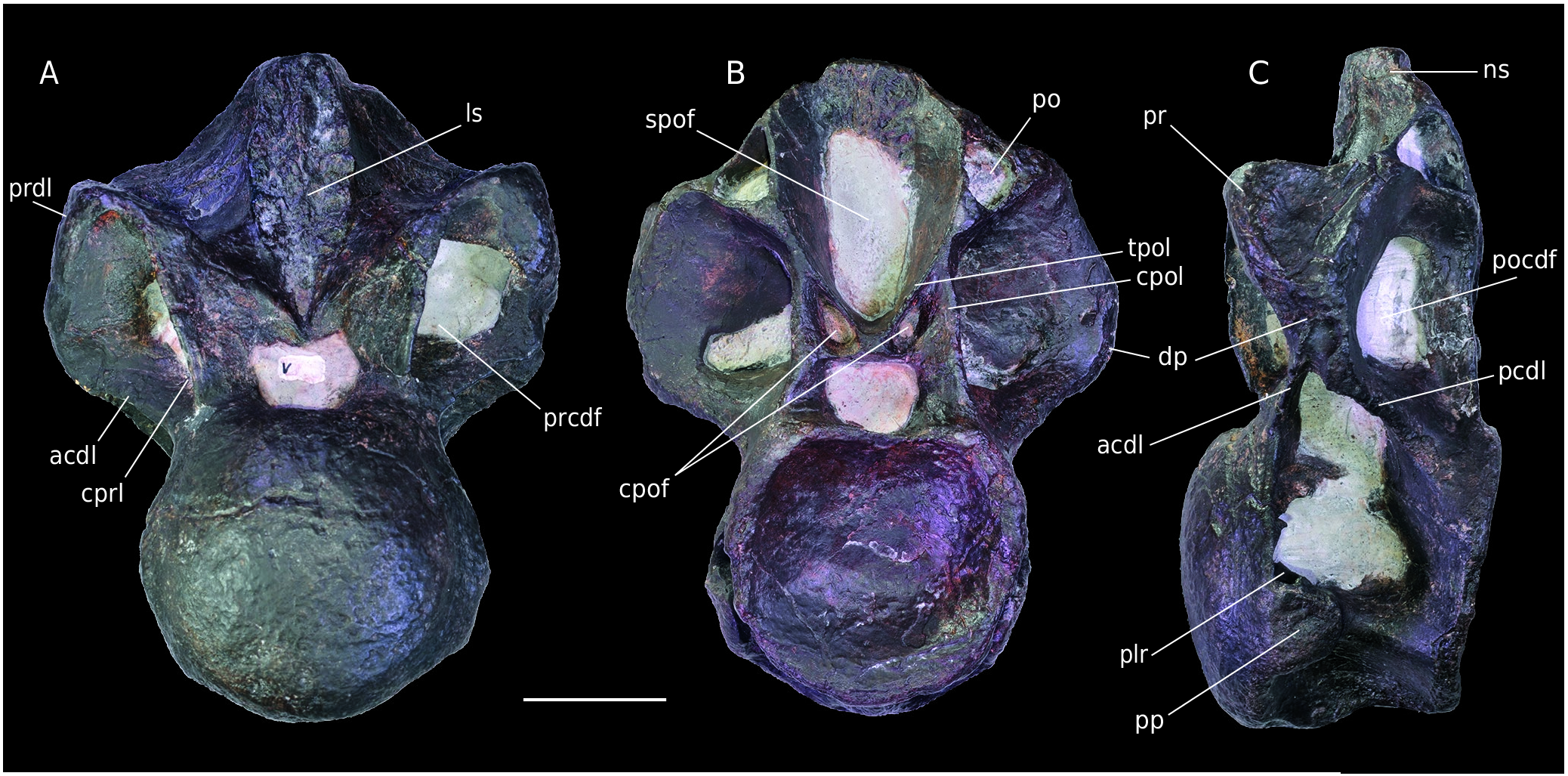
Cervical/Dorsal vertebra
Material. MPV 2020.1.9, last cervical or first dorsal vertebra ( Fig. 13 View FIG ).
Stratigraphy. Lower Oxfordian marls according to the A.gregareum shells in the matrix.
The vertebra is well preserved but the diapophyses and postzygapophyses are fractured. The distal part of the neural spine is missing. The specimen is highly pneumatized, but fossae are filled by the matrix.
The centrum is opisthocoelous and laterally pneumatized with a large pleurocoel on each side, as in B3. These pleurocoels are horizontally divided by a bony lamina. The suboval parapophyses are just below the pleurocoels, at half the height of the centrum. The ventral face of the centrum bears a pronounced ventral keel interpreted as an hypapophysis. The anterior articular facet is rounded and convex. As in vertebra B3, the facet extends laterally to merge with the parapophyses. The posterior articular facet is concave and slightly ventrally inclined. MPV 2020.1.9 is particularly pneumatized and bears the same laminae and fossae as B3. The prezygapophyses have subtriangular, dorsomedially oriented facets. In lateral view, they connect with the anterodorsal part of the diapophyses via a strong prezygodiapophyseal lamina. The spinoprezygapophyseal fossa is only visible just above the neural arch. A wide and high scar is present all over the anterior side of the neural spine. Large triangular fossae are present below the prezygapophyses on the lateral side. They are framed anteriorly by the centroprezygapophyseal laminae, dorsally by the prezygodiapophyseal laminae and ventrally by the anterior centrodiapophyseal laminae. These cavities correspond to the prezygapophyseal centrodiapophyseal fossae ( Wilson et al. 2011) ( Fig. 13A View FIG ). The anterior centrodiapophyseal laminae and posterior centrodiapophyseal laminae merge laterally to support the diapophysis above the centrodiapophyseal fossa ( Fig.13C View FIG ). The position of the laminae and the fossae indicates that the diapophyses were horizontally oriented. In lateral view the diapophyses are triangular. There is a large postzygapophyseal centrodiapophyseal fossa posteriorly to the diapophysis. This structure is framed ventrally by the posterior centrodiapophyseal lamina, dorsally and posteriorly respectively by the postzygodiapophyseal and centropostzygapophyseal laminae. Only the triangular bases of the postzygapophyses remain.They are higher than the diapophyses, hollow and matrix-filled. This shows once again the highly pneumatized condition of the vertebra. A large postspinal fossa is present medially to the postzygapophyses. Above it, as in B3, two small symmetrical fossae are visible ( Fig. 13B View FIG ); they are framed in the same way as in the other specimen (B3), but more elongated. Between these two centropostzygapophyseal fossae, a quadrangular vertical strut is visible. This structure is to be replaced by the hyposphene, to articulate with the hypantrum, in more posterior vertebrae. The vertical strut seems quite developed and it could indicate an early development the hyposphene/hypantrum complex, in contrast with Allosaurus and Torvosaurus , in which it appears from the fifth to the sixth dorsal ( Madsen 1976; Britt 1991). Sinraptor possesses this articulation as early as the first dorsal ( Currie & Zhao 1994). The preserved part of the neural spine is thin and is limited to a bony link between the spinoprezygapophyseal and spinopostzygapophyseal fossae. The bottoms of both these fossae bear an interspinous ligament scar. By comparison with Allosaurus , the low position of the parapophyses and the presence of a hypapophysis indicate a last cervical or first or dorsal vertebra.

| MPV |
Museo Paleontologico Municipal de Valencia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.