
Memphis editha ( Comstock, 1961 ) Dias & Casagrande & Mielke, 2012
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3343.1.2 |
|
persistent identifier |
https://treatment.plazi.org/id/03F3251D-FFA3-FFBA-FF00-FC4A42B8542E |
|
treatment provided by |
Felipe |
|
scientific name |
Memphis editha ( Comstock, 1961 ) |
| status |
|
Memphis editha ( Comstock, 1961) , stat. nov. ( figs 16–26 View FIGURES 16–19 View FIGURES 20–26 )
Anaea (Memphis) leonida editha Comstock, 1961 pp. 144 , 145, pl. 27, fig 7 View FIGURES 1–8 (male, dorsal and ventral); holotype; Santa Catarina, Brazil (AMNH).
Memphis leonida editha ; Lamas, 2004, p. 227.
Diagnosis: Both sexes are superficially similar to many species of Memphis from Comstock’s species group VIII ( Comstock 1961). Males are particularly similar to some specimens of the very variable M. moruus stheno (Prittwitz) , however, the forewing submarginal blue spots of M. editha are always suffused; the tail at M 3 is usually longer; the anal angle is rather acute, almost lobed; and the underside of the wings are chestnut red, with several random white spots and characteristic black markings. Females of M. editha can be distinguished of those of M. moruus stheno by the extended basal blue sheen on the upper side of the forewing, with a pattern somewhat reminiscent of the submarginal spots of the male. Additionally, the outer margin of the forewing is rounded, and the apex is never as falcate as most M. moruus stheno . The underside of the wings of females of M. editha are brown, speckled with several random dark brown and white scales, and are never dull beige or greenish in the outer margin of the hind wing as females of M. acidalia victoria (Druce) .
Redescription: ♂: ( figs 16–17 View FIGURES 16–19 ) Forewing average length 26 mm (23–30 mm) (n=20).
Wing shape: Forewing triangular; costal margin rounded; outer margin sinuous, distinctly angled at CuA 2; apex more or less falcate; inner margin straight, strongly emarginated before tornus; tornus enlarged beyond the inner margin; hindwing humeral area rounded and well-developed; costal margin slightly sinuous; outer margin slightly curved, with a long and blunt tail at vein M 3 and conspicuously angled at the tornus; inner margin slightly curved but not emarginated.
Wing color, upper side: Ground color of both wings dark brown; wing bases with a variable iridescent dark blue suffusion; forewing with two grayish blue bands in the spaces R 4 –R 5, R 5 –M 1 and M 1 –M 2 and markings in spaces CuA 1 –CuA 2 and CuA 2 –2A; seldom with similar markings in spaces M 2 –M 3 and M 3 –CuA 1 and bluish sheen alongside the outer margin; hindwing with minute ocelli on the spaces close to the outer margin, the most conspicuous in space M 3 –CuA 1; light blue scaling on the M 3 projection; and anal fold developed, with long brown scales.
Wing color, underside: Ground color of both wings chestnut red with black and white random speckles; forewing with a conspicuous lighter subtriangular area dorsal to the emargination in the inner margin, and a dented line of white scales from the proximal area of the emargination to the apex; hindwing with minute ocelli on the spaces close to the outer margin, the most conspicuously in space M 3 –CuA 1.
Head: Eyes naked; labial palpi dark brown, speckled with chestnut red and white scales; antennal length about half of the forewing length, segments dark brown with some ventral white scales; club slender and elongated, tip reddish.
Body: Dorsal color of thorax dark brown with random iridescent green and blue scaling; ventral surface and legs chestnut red, speckled with white scales. Abdomen dorsal surface uniformly dark brown; ventral surface uniformly pale brown.
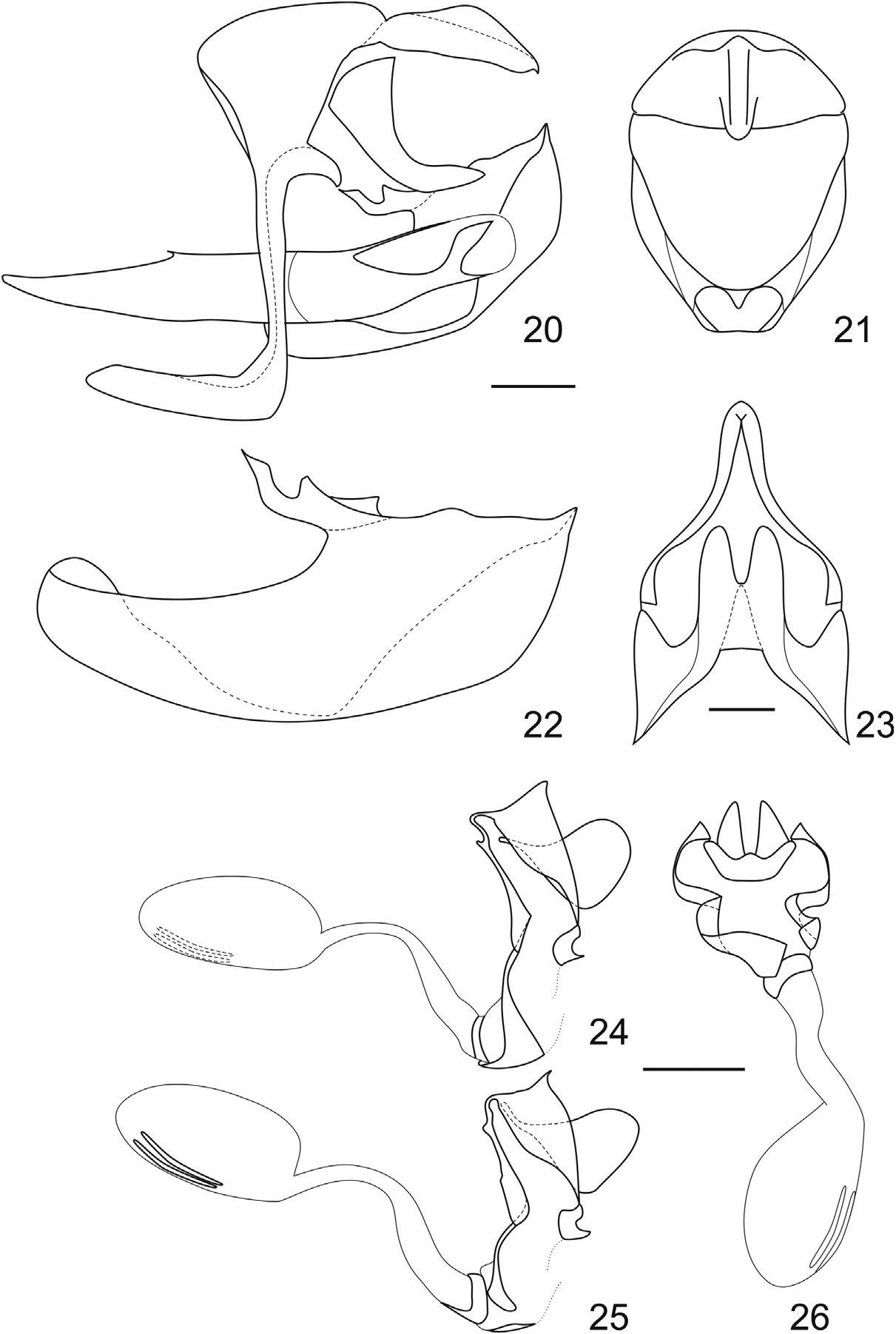
Genitalia ( figs 20–23 View FIGURES 20–26 ): Saccus semitubular, dorsal projection straight and projected dorsad at an acute angle. Tegumen trapezoidal and dorsally curved in lateral view, tightly attached to the anterior part of the uncus and attached to the gnatos by membrane. Gnathos bifid and converging ventrad; ventral part fused medially and strongly projected posteriad, strongly bifid in ventral view. Uncus almost flat and straight, with developed dorsal ridge. Valva ventrally angled, separation between the sacullus and the harpe almost complete at the middle of the valva; pointed projection at the end of the harpe strongly displaced dorsally; ampulla slightly sinuous; costa developed anteriad, with a posterior angled projection. Aedeagus long and slender, folded on the left side and strongly bifid distally. Fultura inferior bifid and subtriangular.
♀ ( figs 18–19 View FIGURES 16–19 ): Forewing average length 30 mm (28–34 mm) (n=20).
Wing shape: As in male, except where noted: forewing outer margin evenly rounded; apex pointed but never falcate; emargination before tornus more developed than male; hindwing larger than male; outer margin strongly rounded and slightly crenulated, with a blunt tail at vein M3, longer than male; outer and inner margins obtusely angled at the tornus; inner margin slightly emarginated.
Wing color, upper side: Ground color of both wings dark brown, duller than male; wing bases with a variable iridescent blue or purple suffusion, lighter than male; forewing with three light blue markings in spaces R 4 –R 5, R 5 –M 1 and M 2 –M 3, markings in spaces CuA 1 –CuA 2 and CuA 2 –2A conspicuous, but partially merged with the basal suffusion; seldom with some blue scaling in spaces R 3 –R 4 and M 3 –CuA 1; occeli and anal fold as in male.
Wing color, underside: Ground color of both wings light brown with dark brown and beige random speckles; forewing pattern similar to male; hindwing with light scaling around the ocelli.
Head: As in male.
Body: Dorsal color of thorax as in male; ventral surface and legs brown, speckled with beige scales. Abdomen dorsal surface as in male; ventral surface uniformly light brown.
Genitalia ( figs 24–26 View FIGURES 20–26 ): Tergum VIII triangular, attached ventrally to the sides of the lamella postvaginalis, and dorsally to the lamella antevaginalis by a slender sclerotized loop; anal papillae round and bristled, projecting the posterior apophysis; posterior edge of the lamella postvaginalis “m” shaped; lamellae antevaginalis thin and assymetrical, with a small dorsal projection; left side much larger than the right, sinuous and ventrally subrectangular; right side slender and sinuous; seminal duct close to the base of the ductus bursae, posterior to a sclerotized ring in the ductus bursae; corpus bursae oval, about two thirds the length of the ductus bursae, bearing two parallel slender signa on the left side.
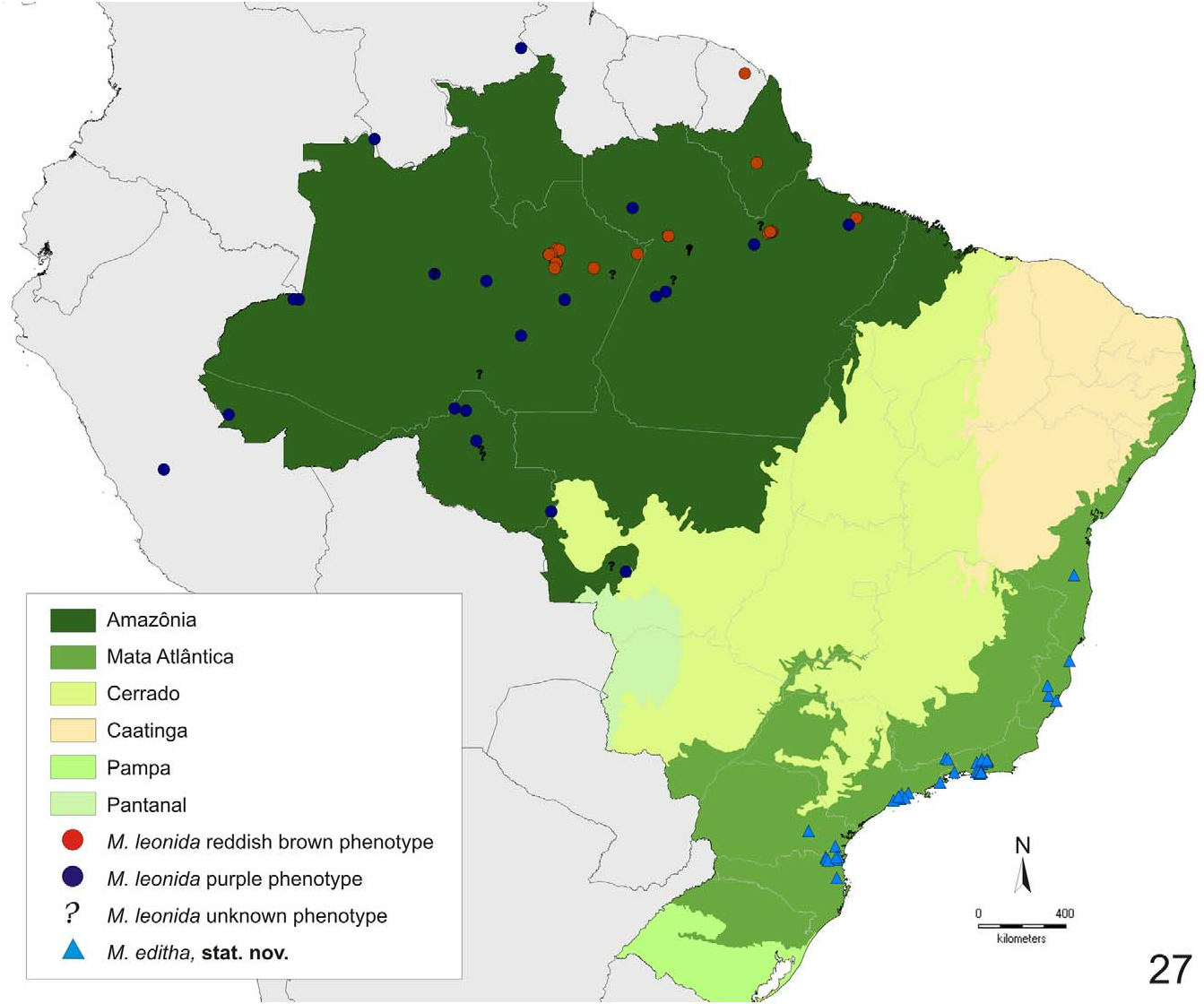
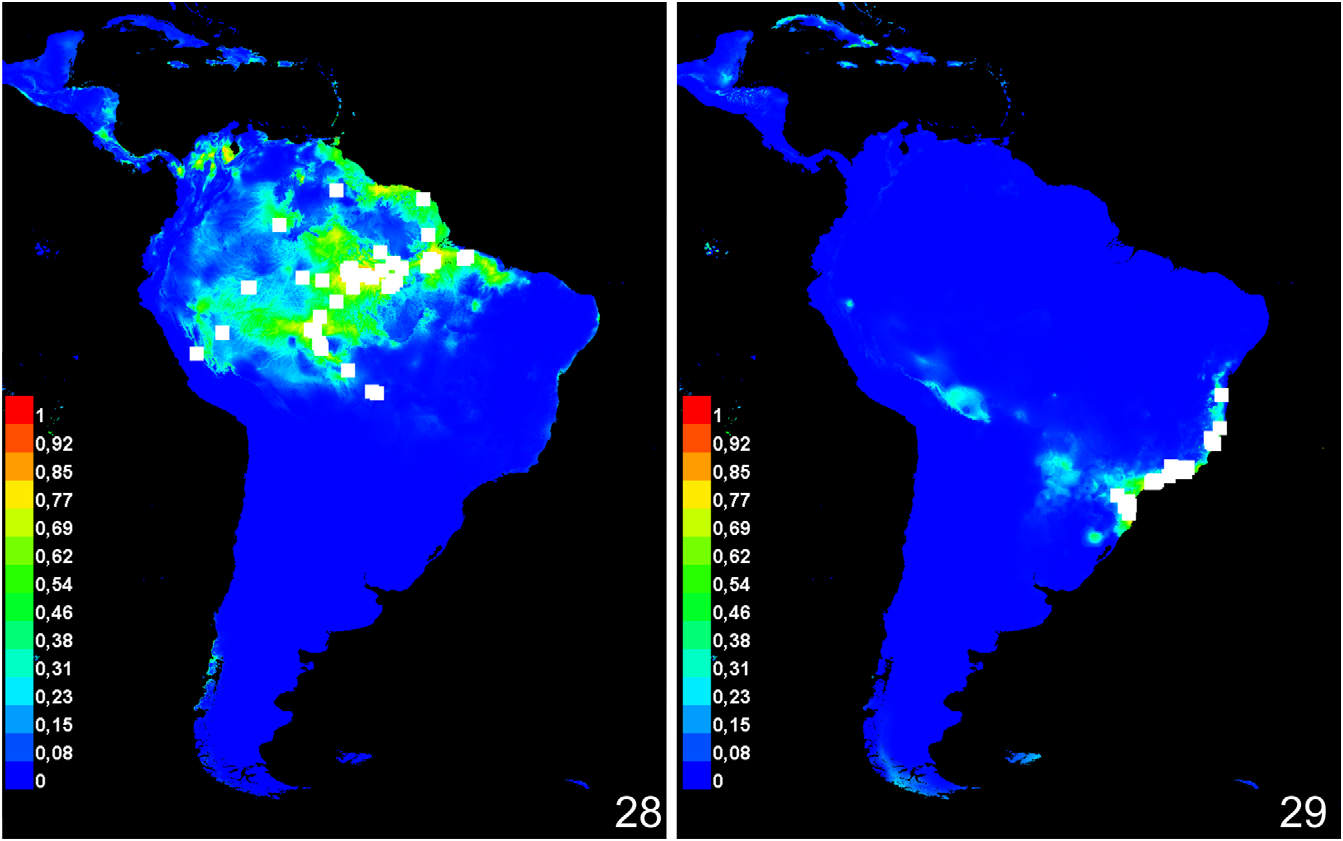
Distribution ( fig. 27 View FIGURE 27 ): Atlantic rain forest along the southeast coast of Brazil: Rio Grande do Sul, Santa Catarina, Paraná, São Paulo , Rio de Janeiro , Espírito Santo and Bahia. Ecological niche modeling (mean AUC = 0.997) predicts the presence of M. editha further west into Minas Gerais and the interior forests of São Paulo and Paraná ( fig. 29 View FIGURES 28–29 ). Ecological niche modeling shows that the most important environmental variables to predict the distribution of M. editha are the preciptation in the driest month (28.9% contribution, 3.3% permutation importance), isothermality (21.2% contribution, 0% permutation importance), precipitation in the driest quarter (20% contribution, 3.3% permutation importance), and temperature seasonality (17.1% contribution, 79.8% permutation importance). Temperature seasonality appears to have the most useful information by itself and have the most information that isn't present in the other variables.
Taxonomic comments: Comstock (1961) described M. editha as a subspecies of M. leonida , based on a series of specimens from the south of Brazil (Santa Catarina and Paraná states). Although some distinction was made between these two taxa, he affirmed that the male of M. editha “does not differ from that of nominate leonida in venation or in male genital armature”. Our detailed investigation showed that both male and female genitalia of M. editha are very different from those of M. leonida , suggesting that M. editha is not closely related with M. leonida or even other species belonging to “ eribotes ” species group.
In most Memphis species the sacculus is strongly developed, with a dorsal rough area, as in M. leonida and M. moruus stheno (Prittwitz) ( Dias et al. 2010a) . However, in “ eribotes ” species group, this condition is not observable in M. editha and in the morphologically anomalous M. hirta (Weymer) and M. otrere (Hübner) . The sacculus not strongly developed, without a dorsal rough area also occurs in all species belonging to II (or “ pasibula ”) and VIIID (or “ iphis ”) species groups; M. anna (Staudinger) , belonging to III (or “ aureola ”) species group; a few species belonging to the large and heterogeneous VIIIA (or “ polycarmes ”) species group; and M. oenomais , belonging to VIIIB (or “ morvus ”) species group. The shape of the ventral part of the gnathos, long and rounded in ventral view, is also similar to species belonging to “ iphis ” species group; M. forreri (Godman & Salvin) , belonging to “ morvus ” species group; M. otrere and M. hirta . The shape of the uncus, straight, with a prominent triangular ridge, is characteristic of M. editha .
The female genitalia, although typical of species belonging to VIII species group ( Dias et al. 2010a), has a sclerotized ring in the posterior part of the ductus bursae. This sclerotized ring is also present in species belonging to “ pasibula ” and “ iphis ” species group, a few species belonging to “ polycarmes ” species group, M. hirta , and M. otrere . The shape of the lamella postvaginalis, “m” shaped and posteriorly strongly indented, is characteristic of M. editha .
In addition to other differences pointed out by Comstock (1961) and described here, M. editha has an unusual wing shape, with the outer margin of the forewing angled and the anal angle of the hind wing almost lobed. It is likely that Comstock’s (1961) subdivisions of VIII species group, based only in some characters of the valva and the presence of a tail at M 3 on the hindwing, are not monophyletic. As Comstock (1961) himself stated, the group-
ings were proposed mainly for taxonomic convenience. M. editha seems to be restricted to the Atlantic rain forests along the coast of Brazil and is abundant in its area of occurrence. This species is not expected to occur further in the Atlantic interior forests, which are the most well sampled region of Brazil ( Carneiro et al. 2008). Larvae use as host plant a number of species of Piper and Peperonia ( Piperaceae ) (J. Bizarro comm. pers.) and are similar to immature stages from Comstock’s (1961) species groups VIIIB and VIIIC (“ morvus ”- type sensu Dias et al. 2010b).
Type material: Anaea (Memphis) leonida editha has the following labels: / leonida editha ♂ 50C/ HOLO- TYPE / A. L. EDITHA W. P. C. DET. W. P. COMSTOCK/ Collection Frank Johnson/ St. Catherines Brazil / ( AMNH).
Examined material: BRASIL, Bahia, Ilhéus, São João, Água Preta , 4 ♂, IX–X.1928 (ex coll May), 29898, 29900, 29901, 29909 ( MNRJ); Espírito Santo, Conceição da Barra, 1 ♂, 29.XII.1969 (C. & C. T. Elias leg.), DZ 19.700 ( DZUP); Santa Teresa, 1 ♂, 26–29.?.1968 (Ebert & Brown leg.), DZ 20.184 ( DZUP); Campinho, 1 ♂, 9.I.1949 (Pinto leg.), DZ 19.580 ( DZUP); Colatina, 2 ♂, VIII–IX.1937 (ex coll May), 52826, 29494 ( MNRJ), #1, no data, 29934 ( MNRJ); Rio de Janeiro , Rio de Janeiro , 2 ♂, no data, 1 ♂, 3.VIII.1962 (ex coll Gagarin), DZ 20.017, DZ 19.337, DZ 20.058 ( DZUP), 1 ♂, 1 ♀, no data (ex coll D’Almeida), DZ 19.967, DZ 20.616 ( DZUP), 1 ♂, no data (ex coll M. Rosa), 51469 ( MNRJ); Rio de Janeiro, Jacarepaguá , 1 ♂, 8.VI.1952 (Pearson leg.), ( MNRJ); Rio de Janeiro, Jacarepaguá, Três Rios , 1 ♂, 14.VII.1922 (ex coll D’Almeida), DZ 19.749 ( DZUP); Rio de Janeiro, Gávea , 1 ♂, 27.VIII.1940 (ex coll Gagarin), DZ 19.927, ( DZUP); 1 ♂, no data ( MNRJ), 4 ♂, no data (ex coll May), 29824, 29814, 29815 ( MNRJ); 1 ♂, no data (Guinle & Oiticica leg.), 5934 ( MNRJ); Rio de Janeiro, Paineiras , 1 ♀, 20.IV.1952, 1 ♀, 4.IV.1954 (H. Ebert leg.), DZ 19.711, DZ 19.856* ( DZUP), 1 ♂, no data, 1 ♀, 2.I.1955 (ex coll Gagarin), DZ 20.573, 20.121 ( DZUP); 3 ♂, no data (ex coll May), 29839, 29820, 9833 ( MNRJ), 1♂ II.1926 (ex coll May), ( MNRJ), 1 ♂, III.1927 (ex coll May), ( MNRJ), 2 ♂ I.1927 (ex coll May), 6830, 6832 ( MNRJ), 1 ♂ IV.1923 (ex coll May), 6831 ( MNRJ) 1 ♂ IV.1946 (ex coll A. Costa), 4711 ( MNRJ); Rio de Janeiro, Morro do Sumaré, 1 ♂, 31.VII.1967 (H. Ebert leg), DZ 19.903 ( DZUP), 4 ♂, no data (ex coll A. Costa), 1540, 1639, 2145, 7558 ( MNRJ); Guapimirim, 1 ♀, no data (Sandig leg.), DZ 19.722 ( DZUP), 1 ♂, 29.VI.1941, 1 ♂, 20.VIII.1940 (ex coll Gagarin), DZ 19.259, DZ 19.840 ( DZUP), 1 ♂, 19.II.1964, 1 ♀, 24.II.1965 (Ebert leg.), DZ 19.425*, DZ 19.384 ( DZUP); Guapimirim, Caneca Fina, Rio Sucavão , 5 ♂, 3-4.VII.71, 31.VII.72, 1.V.1972, 23.VI.1975, 12.VII.1976 (Pearson leg.), ( MNRJ); Resende, Penedo, 1 ♀, 24.VI.1956 (ex coll Frey), DZ 19.672 ( DZUP); Xerém, Serra do Tinguá, 1 ♀, 30.VII.1968 (Ebert leg.), DZ 19.620 ( DZUP); Petrópolis, 1 ♂, V.1936 (ex coll Oiticica), ( MNRJ); Petrópolis, Meio da Serra , 1 ♂, 3.V.1941, 2 ♂, 10.V.41 (Parko leg.), 5983, 5948, 5990 ( MNRJ); Duque de Caxias, Imbariê, 2 ♂, 20.II.1965, 1♂, 15.II.1966, 1♂, 16.II.1966 (H. Ebert), DZ 20.043, DZ 20.600*, DZ 20.090, DZ 20.138 ( DZUP); Itatiaia, south side of Serra do Itatiaia, 1 ♂, 3.XI.1967 (H. Ebert leg.), DZ 19.767 ( DZUP); Teresópolis, Barreira, 23.III.1955 (ex coll Gagarin), DZ 19.804 ( DZUP); Teresópolis, Guapi-Colomi, Serra dos Órgãos , 2 ♂, 3 ♀, no data (Parko leg.), 5985 ( MNRJ), 1 ♂ 20.III.1941 (Parko leg.), 5984 ( MNRJ); Angra dos Reis, Jussaral , 2 ♂, II.1937 (ex coll A. Costa), 4713, 4714 ( MNRJ); Paraná, Ponta Grossa, 10 KM E, 7.II.2010 (O.& C.Mielke leg.), DZ 19.635* ( DZUP); Guaratuba, Castelhanos, 3 ♂, 10.I.2010 (Dolibaina leg.), DZ 19.574, DZ 20.161, DZ 19.917 ( DZUP); Santa Catarina, Brusque, 1 ♂, 20.I.1968 (V. Becker leg.), DZ 20.130 ( DZUP); Joinville, 1 ♂, 9.II.1969, 8 ♂, 1 ♀, 9.X.1971, 1 ♂, 20.XI.1970 (O. Mielke leg.), DZ 19.713*, DZ 19.941, DZ 20.007, DZ 20.305, DZ 19.372, DZ20.640, DZ 20.127, DZ 20.069, DZ 19.230, DZ 19.677*, DZ 20.465 ( DZUP), 2 ♂, 4.II.1973, 1 ♂ 10.X.1971, 1 ♂, XI.1971 (Miers leg.), DZ 20.630, DZ 20.276, DZ 19.219, DZ 19.940 ( DZUP), 3 ♂, II.1971 (H. Ebert leg.), DZ 19.270, DZ 20.145, DZ 19.325 ( DZUP), 1 ♂, 16–21.I.1971 (H.& K. Ebert leg.), DZ 20.547 ( DZUP), 1 ♂, no data, 23109 ( MNRJ), 1 ♂, no data (ex coll Arp), ( MNRJ), 4 ♂, no data (Schmidt leg.), 6840, 6842, 6837, 6838 ( MNRJ), 2 ♂, no data (Brückner leg.), 6860, 6861 ( MNRJ); Joinville, Neudorf, 1 ♂, 31.V.41, 1 ♂ 10.V.1941 (Schmidt leg.), 5974 ( MNRJ); Joinville, Morro do Finder, 1 ♂, 1 ♀, 13.VIII.2001, 1 ♂, 21.VIII.2001, 1 ♂, 4.XI.2003, ex-larva Piper sp. (Bizarro leg.), DZ 19.974*, DZ 20.125, DZ 20.336*, DZ 19.816 ( DZUP); São Bento do Sul, Rio Vermelho , 1 ♂, VII.2007 (Rank leg.), DZ 20.605 ( DZUP); São Bento do Sul, Rio Natal , 1 ♀, VII.2009 (Bizarro leg.), DZ 19.355 ( DZUP); São Bento do Sul, 1 ♂, XII.1936 (Weiss leg.), DZ 19.450 ( DZUP); Rio Grande do Sul, Torres, 3 ♂, 2.I.2011, 3.IV.2011, 5.IV.2011, 3 ♀, 4.I.2011, 24.I.2011, 3.4.2011 ( CLDZ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Memphis editha ( Comstock, 1961 )
| Dias, Fernando Maia Silva, Casagrande, Mirna Martins & Mielke, Olaf Hermann Hendrik 2012 |
Memphis leonida editha
| Lamas, G. 2004: 227 |