
Sandsitermes, Cuezzo, Carolina, Cancello, Eliana M. & Carrijo, Tiago F., 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4221.5.5 |
|
publication LSID |
lsid:zoobank.org:pub:2779DDB4-83EB-49C1-8247-3222D1FE3322 |
|
DOI |
https://doi.org/10.5281/zenodo.6021699 |
|
persistent identifier |
https://treatment.plazi.org/id/03F42441-FFF6-FFE6-FF3F-38F0FADEC0EF |
|
treatment provided by |
Plazi |
|
scientific name |
Sandsitermes |
| status |
gen. nov. |
Genus Sandsitermes , new genus
Type-species. Eutermes robustus Holmgren (1906) , by present designation.
Species included. Sandsitermes robustus (Holmgren) , new combination
Etymology. The generic epithet “ Sandsi ” is given in tribute to Professor William A. Sands, distinguished termitologist who made important contributions to the knowledge of termites; as a prefix to “ termes ”, termite.
Distribution. Neotropical region: Peru.
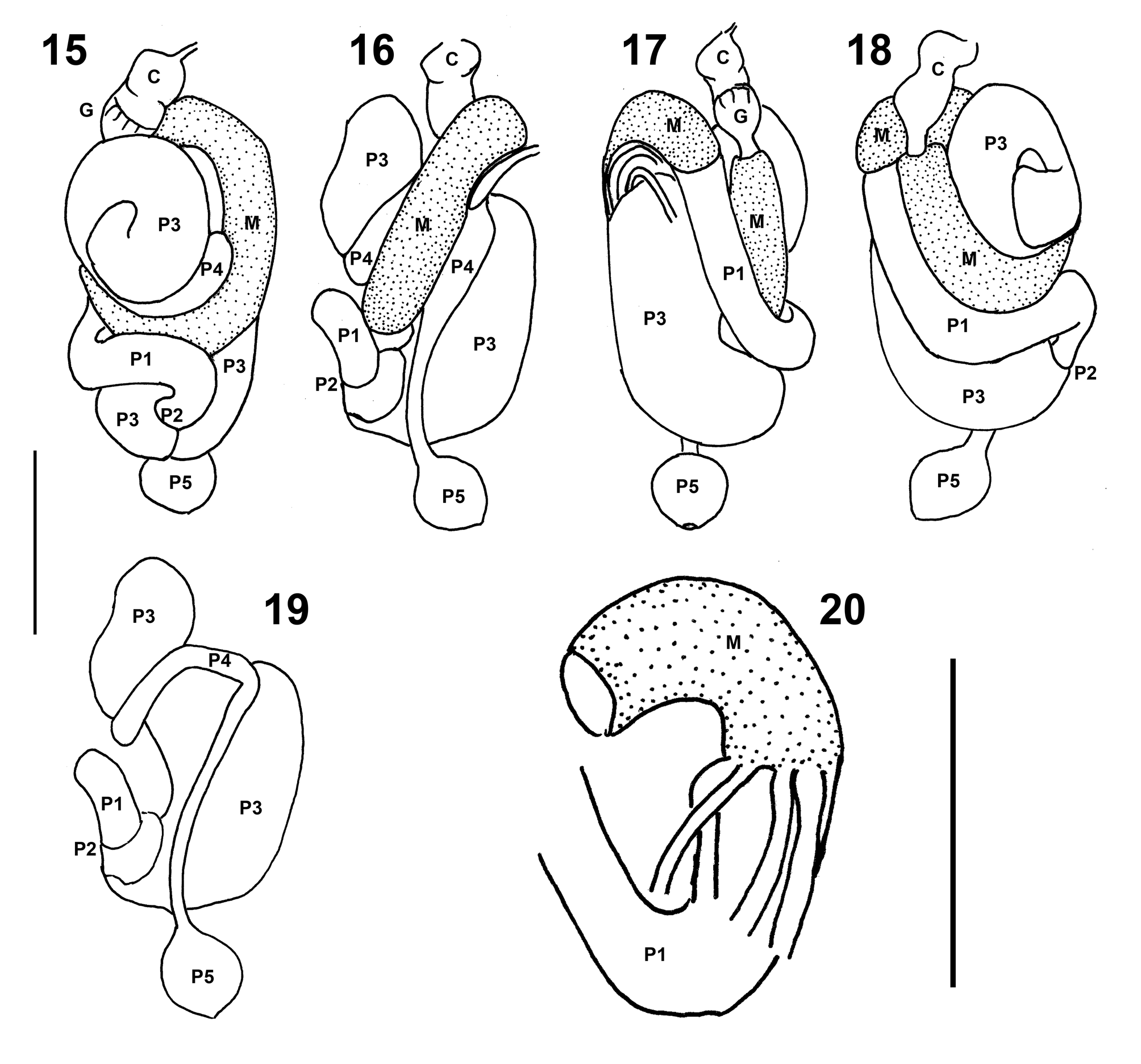
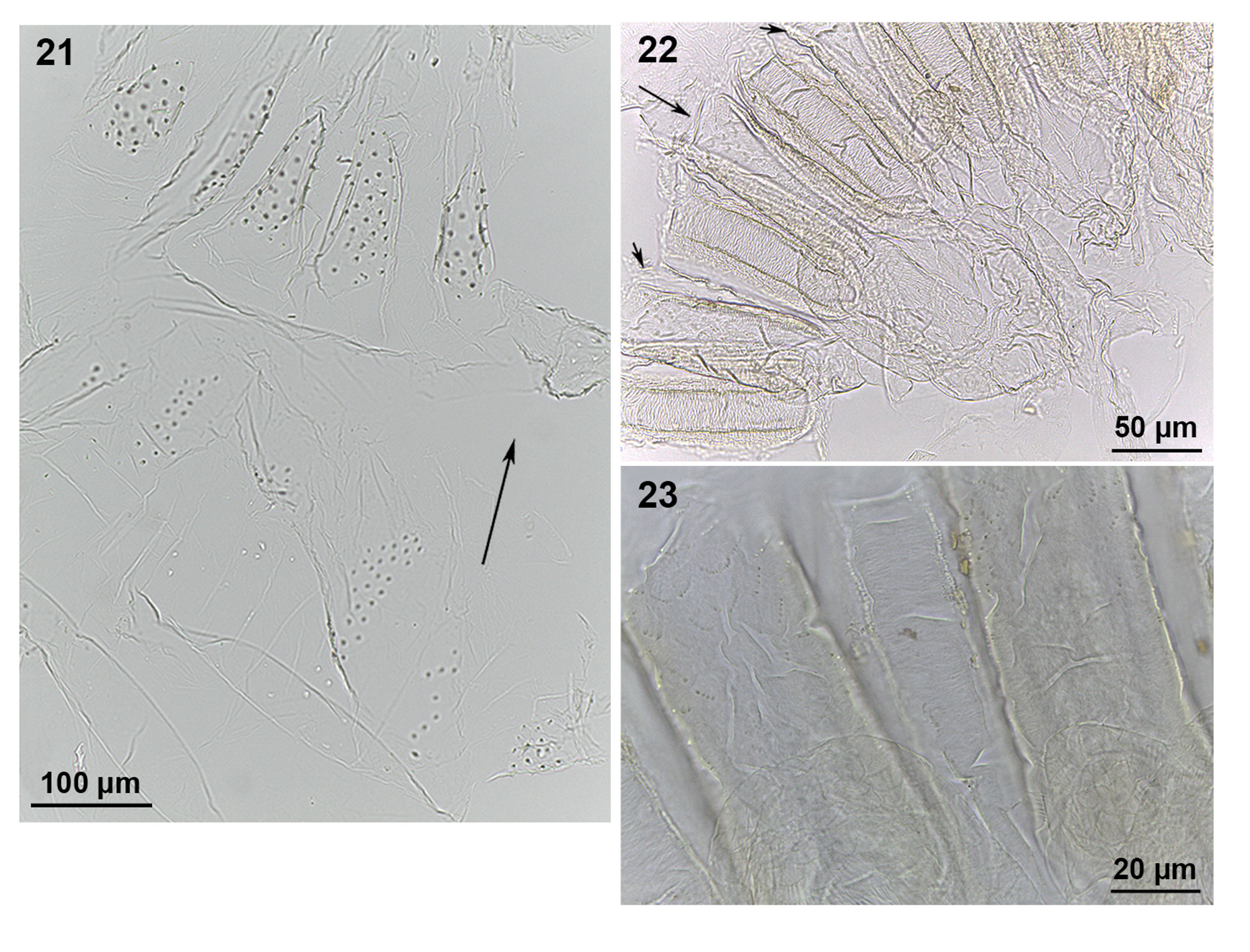
Diagnosis. Imagoes with dorsal surface of head capsule with slight elevation anterior to fontanelle, where the ocelli are situated; postclypeus slightly arched, with anterior margin straight and posterior margin faintly convex; right mandible with distinctly concave molar area with faint ridges and distinct basal notch. Soldiers with long conical nasus, but no apparent uniquely distinct morphological character. Workers showing distinct set of morphological characters involving mandibles with concave molar area with no well-developed ridges, gut coiling with distinct first proctodeal segment (P1) loop before junction with paunch (P3) ( Figs 15–18 View FIGURES 15 – 20 ), and enteric valve (P2) configuration weakly sclerotized, organized in two distinct rings with tiny conical spines ( Fig. 21 View FIGURES 21 – 23 ).
Description. Imago ( Figs 1–3 View FIGURES 1 – 3 ). In dorsal view, head capsule oval. Area between ocelli, at frontal region, not depressed. In profile, dorsal surface of head capsule with slight elevation anterior to fontanelle, where the ocelli are situated. Eyes oval, large, separated from lower margin of head capsule by shorter distance than from eye ocelli. Ocelli elliptical in dorsal view, large, although smaller than antennal socket, situated in anterior half of eyes, and separated from them by less than ocellus width. Fontanelle paler than head, situated in slight depression, Y-shaped, with short anterior arms, and small clear circle above arms. Postclypeus slightly arched in profile, at midline slightly depressed in dorsal view; anterior margin of postclypeus straight and posterior margin faintly convex. Antenna with 15 articles. Mandibles similar to type 2 workers but molar area less concave. Pronotum wider than head without eyes, subtrapezoidal, with anterior margin elevated, anterior lateral corners rounded, lateral margins concave and converging at rear, and posterior margin rounded and not emarginate. Posterior margin of meso- and metanotum deeply emarginate; posterolateral corners of meso- and metanotum pointed. Tibial spurs 2:2:2.
Soldier ( Figs 4–5 View FIGURES 4 – 5 ). Monomorphic. In dorsal view, head capsule varying from oval to rounded, not constricted, and posterior margin not emarginate. In profile, dorsal margin of head capsule with slight hump at the base of nasus, followed by slight depression. In profile, nasus conical, shorter than head-capsule length, and not directed upward. Postclypeus slightly convex in profile. Labrum shorter than wide, with rounded anterior margin, parallel lateral margins and rounded anterior corners. Mandibles with well-defined ‘points’. Antenna with 13 articles. Pronotum with anterior lobe less developed than posterior lobe, with an angle between them of approximately 90°. Procoxa conical, lacking keel on anterior surface. Tibial spurs 2:2:2. Abdominal sclerites well sclerotized, tergites imbricate.
Worker ( Figs 6–11 View FIGURES 6 – 11 ). All workers with trapezoidal head capsule, conspicuous fontanelle situated at middle of head capsule, slightly depressed; postclypeus inflated; antenna with 14 articles; anterior margin of pronotum rounded, not emarginate. Tibial spurs 2:2:2. Two types of workers, distinguished by head-capsule pigmentation, mandible morphology, and pronotum shape and size. Type 2 worker ( Figs 6–7 View FIGURES 6 – 11 ) less frequent than type 1 in al samples studied, differing from it in having head capsule brownish yellow with concavity at front followed by hump; mandibles with external margin showing a conspicuous concavity before apical teeth, apical teeth larger, left M3 separated from molar prominence by wider gap, and narrower molar plate; anterior lobe of pronotum more developed than posterior lobe. There are apparently differences in the head-capsule size in the type 1 category, but we lack enough individuals to measure ( Figs 8–11 View FIGURES 6 – 11 ).
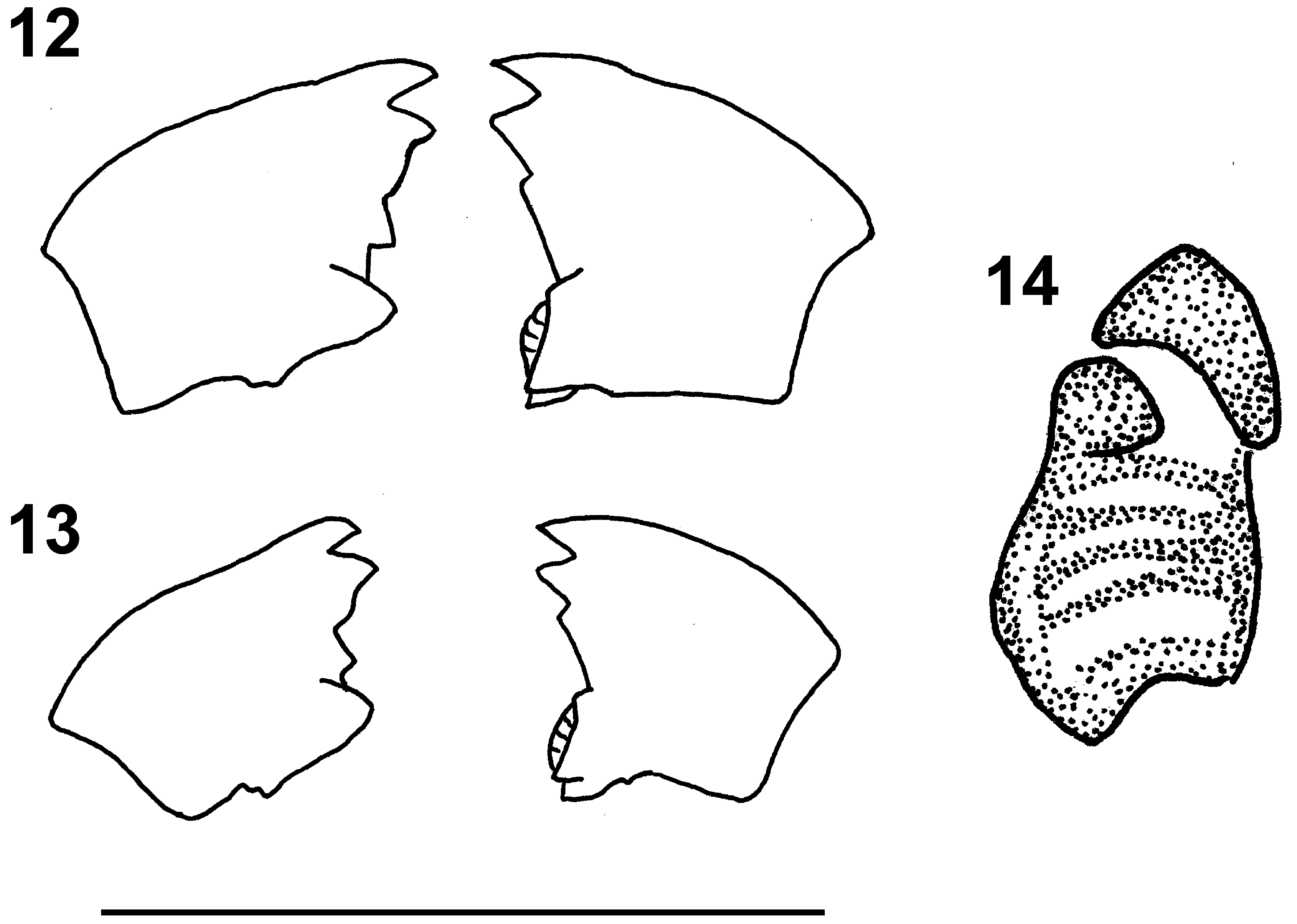
Mandibles ( Figs 12–14 View FIGURES 12 – 14 ). Posterior margins of apical teeth almost straight; posterior margins of apical teeth and anterior margins of first marginal teeth forming acute angle. Left mandible: posterior margin of first marginal tooth almost straight; cutting edge between M1+2 and M3 slightly convex; M3 distinct, but smaller than M1+2 and separated from molar prominence by gap; molar tooth not visible through small gap, its apex hidden beneath molar prominence; molar prominence concave with reduced ridges. Right mandible: M2 distinct, smaller than M1; M2 situated nearer M1 than molar plate; posterior margin of second marginal tooth straight; molar plate concave with reduced ridges compared to those of N. corniger ; basal notch well defined, as 90° angle ( Figs 12–14 View FIGURES 12 – 14 ).
Digestive tube ( Figs 15–23 View FIGURES 15 – 20 View FIGURES 21 – 23 ). Gut coiling in situ forming broad mass. Crop well developed, cuticle ornamented with pectinate scales. Crop and gizzard well separated, equally developed, not voluminous, partially visible at left half of abdomen in dorsal view ( Fig. 15 View FIGURES 15 – 20 ). Gizzard with completely sclerotized cuticular armature (hexaradial symmetry); pulvillar belt slightly more developed than columnar belt, pulvilli I well developed and pulvilli II reduced, both with pectinate scales ( Fig. 22 View FIGURES 21 – 23 ); columnar belt with surface of folds I and II ornamented with scales in proximal area, but difficult to discern ( Fig. 23 View FIGURES 21 – 23 ). Mixed segment well developed; single mesenteric tongue external to mesenteric arc, not constricted proximally. Malpighian tubules arranged in two adjacent pairs but attached individually at mesenteron-proctodeum junction on inner margin of mesenteric arc; tubules dilated along length of mixed segment, tapering after this point. Ileum (P1) tubular, longer than mesenteron length, reaching median line dorsally, after recurving to form short loop distally on right side of abdomen, and ending at enteric valve (P2), which enters directly into paunch (P3). P2 armature weakly sclerotized, organized in two distinct rings with tiny conical spines ( Fig. 21 View FIGURES 21 – 23 ); anterior ring (or upper ring, closest to P1) with spines barely organized in three large cushions alternating with three smaller cushions; posterior ring (or lower ring, closest to P3) with six equal subtriangular cushions. Paunch (P3) as pyriform sac; P3b protruding through mesenteric ring and joining colon (P4) on left side; isthmus conspicuous. Dorsal torsion well developed. P4 tubular, U-turn not dilated, visible in right lateral view. Distal colon also tubular, reaching rectum (P5) in dorsal view. No differences are apparent in the coiling gut of worker types 1 and 2.
Comparisons. The imago caste is known from at least the type-species of the following Neotropical genera of the Nasutitermes group: Caetetermes Fontes ( Cuezzo et al. 2015a, figs 1–3), Caribitermes Roisin, Scheffrahn & Křeček (Scheffrahn et al. 2003 , figs 27, 30), Constrictotermes Holmgren ( Mathews 1977, fig. 145), Cortaritermes Mathews (Mathews 1977 , fig. 146; Cuezzo et al. 2015b, figs 1–3), Diversitermes Holmgren ( Mathews 1977, fig. 124; Oliveira & Constantino 2016, fig. 2), Muelleritermes Cancello, Oliveira & Rocha (Oliveira et al. 2015 , figs 1, 2), Nasutitermes Dudley ( Mathews 1977, figs 114, 119), Obtusitermes Snyder , Parvitermes Emerson , Rotunditermes Mathews ( Holmgren 1906, fig. S1), Tenuirostritermes Holmgren ( Scheffrahn & Rust 1983, figs 7– 10) and Velocitermes Holmgren. The imagoes of Sandsitermes , gen. nov., Caeteterme s, Caribitermes , Cortaritermes , Nasutitermes and Obtusitermes have the postclypeus slightly arched in profile, with its anterior margin straight and the posterior margin slightly convex; while Constrictotermes , Diversitermes , Muelleritermes , Parvitermes , Rotunditermes , Tenuirostritermes and Velocitermes have a noticeably arched postclypeus. The imago of Sandsitermes is distinguished from those of Caribitermes , Cortaritermes and Nasutitermes by the morphology of the right mandible, which has a distinctly concave molar area with faint ridges and a distinct basal notch, more resembling Caetetermes , but differing in the straight posterior margin of M2 of the right mandible.
Remark 1. Not only are the imagoes of many species unknown, but the description of this caste has been neglected in comparison to other castes, and good illustrations have only recently appeared in the literature. Moreover, in many cases, the imagoes of different species of the same genus are very dissimilar, and a new diagnosis is necessary (e.g., Cyranotermes timuassu Araujo and Cyranotermes karipuna Rocha, Carrijo & Cancello ). Considering these points, it is difficult to make a comprehensive comparison among all the imagoes herein mentioned, and we prefer to use the characters already accepted as actually important in the diagnosis for each genus.
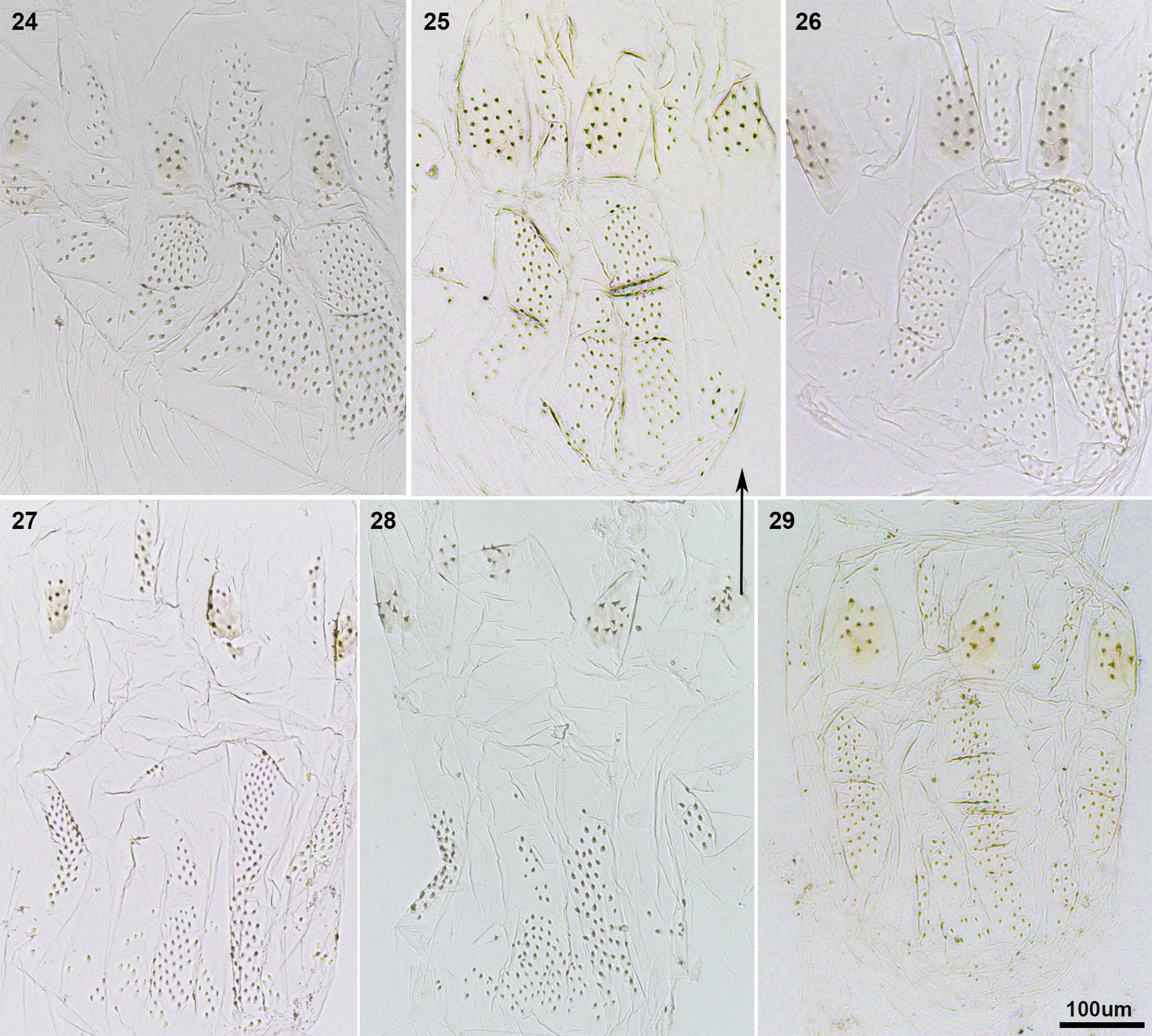
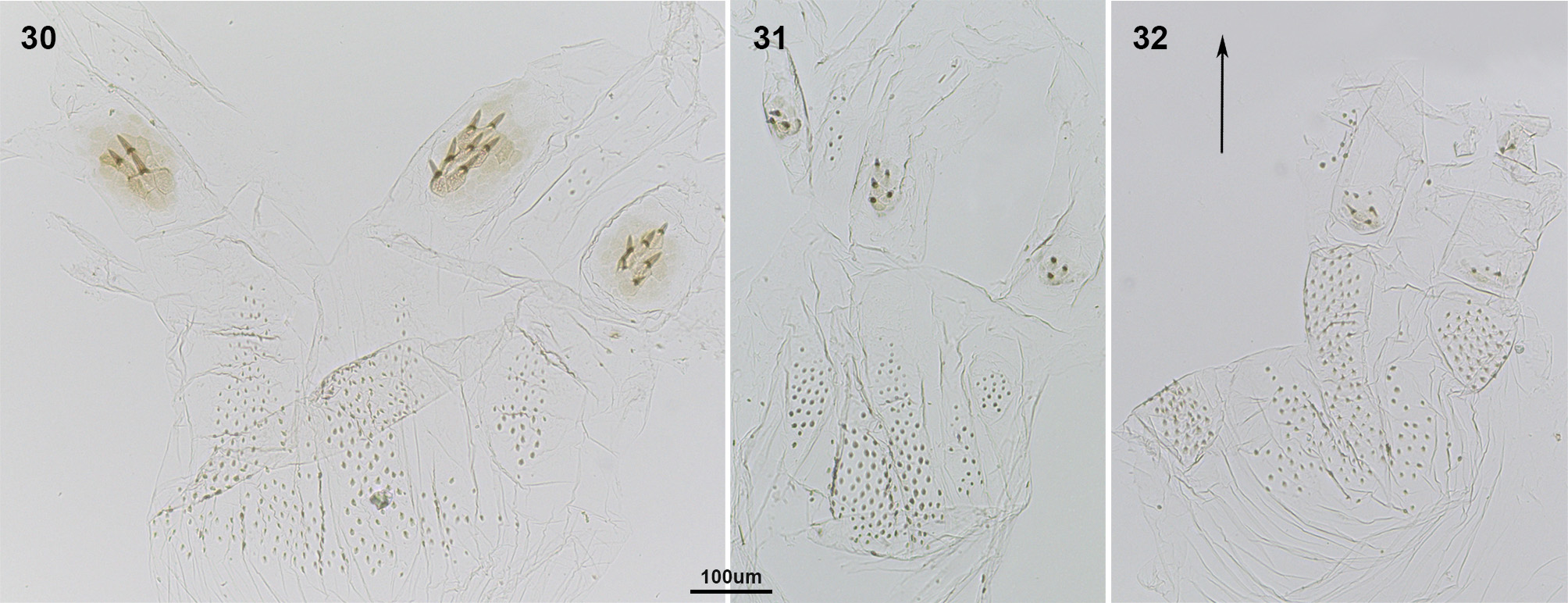
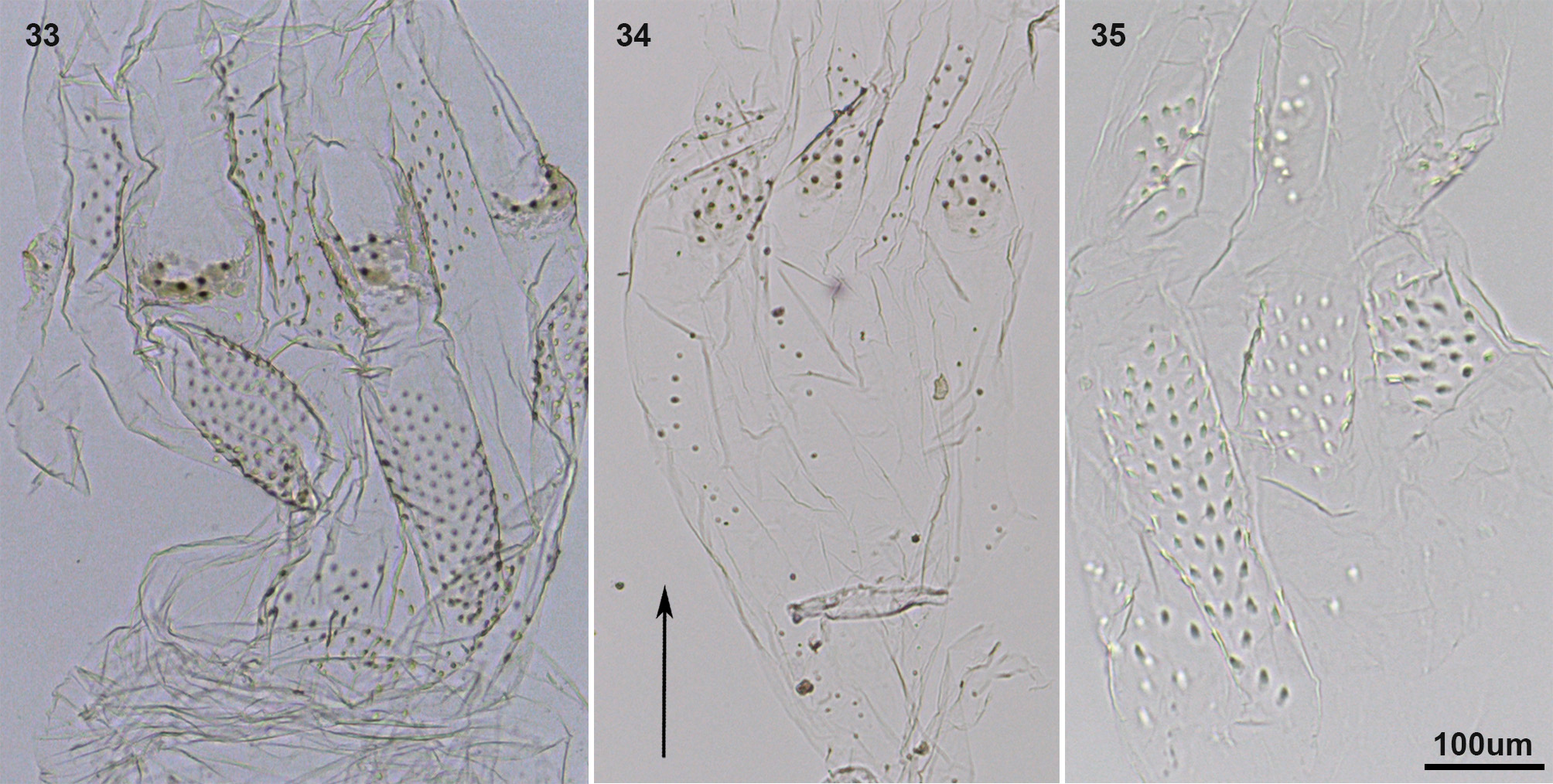
Remark 2. We studied the enteric valves of 13 Neotropical Nasutitermes species, in order to compare them with Sandsitermes . After recognizing the anterior ring and the posterior ring of cushions of these species, we noted that the size, shape and position of cushions vary among the species, for instance: 1) posterior ring show alternating larger and smaller cushions, with the larger having more robust spines, as in N. aquilinus ( Fig. 24 View FIGURES 24 – 29 ; see also Martegani & Torales 1994, figs 1–4), N. jaraguae , N. stricticeps ( Fig. 25 View FIGURES 24 – 29 ), N. surinamensis ( Fig. 26 View FIGURES 24 – 29 ), N. rotundatus ( Fig. 27 View FIGURES 24 – 29 ), N. octopilis ( Fig. 28 View FIGURES 24 – 29 ), N. comstockae ( Fig. 29 View FIGURES 24 – 29 ); 2) posterior ring show a reduction in armature of the smaller cushions, but an increase in spine size of the larger cushions, as in N. acangussu Bandeira & Fontes ( Fig. 30 View FIGURES 30 – 32 ), N. corniger (see Martegani & Torales 1994, figs 5–8, Constantino & Acioli 2009, fig. 6a), N. macrocephalus ( Fig. 31 View FIGURES 30 – 32 ) and N. similis ( Fig. 32 View FIGURES 30 – 32 ). Furthermore, N. wheeleri ( Fig. 33 View FIGURES 33 – 35 ) is distinct from all other species mentioned because some cushions of the anterior ring are reduced, forming a spiny area. Similarly, N. hubbardi ( Fig. 34 View FIGURES 33 – 35 ) and N. banksi ( Fig. 35 View FIGURES 33 – 35 ) both have a distinct ring organization, which is a matter for future studies, and may support their treatment as distinct genera, separate from Nasutitermes . In summary, no Nasutitermes species has six equal subtriangular cushions in the posterior ring, and a little-developed anterior ring forming six areas of spines. Sandsitermes perhaps belongs as part of a natural group with Caetetermes Fontes ( Cuezzo et al. 2015a, fig. 15).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.