
Pethia punctata ( Day, 1865 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3884.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:ECA4FF1F-DC78-481A-9C4B-8CEDE704DF70 |
|
DOI |
https://doi.org/10.5281/zenodo.5628549 |
|
persistent identifier |
https://treatment.plazi.org/id/03F49B7C-C370-E016-B8D1-F89BFDFFFDC2 |
|
treatment provided by |
Plazi |
|
scientific name |
Pethia punctata ( Day, 1865 ) |
| status |
|
Pethia punctata ( Day, 1865) View in CoL
Puntius punctatus Day, 1865 View in CoL
Barbus punctatus ( Day, 1865) View in CoL : Day (1873)
Barbus (Puntius) ticto View in CoL (non Hamilton, 1822): Hora et al. (1939) Puntius ticto View in CoL (non Hamilton, 1822): Jayaram (1991), Talwar & Jhingran (1991) Puntius ticto punctatus Day, 1865 View in CoL : Rema Devi (1992)
Puntius muvattupuzhaensis Beevi & Ramachandran, 2005 View in CoL
Pethia muvattupuzhaensis ( Beevi & Ramachandran, 2005) View in CoL : Pethiyagoda et al. (2012)
Material examined: Day’s material (syntype?), 1 ex., MCZ 4303, Canara ( Cannanore, on the Malabar Coast, Kerala, India), coll. F. Day (photograph examined); Day’s material (syntype?), 1 ex., BMNH 1889.2.1.755, Wayanad, Kerala, India, coll. F. Day (photograph examined); 7 ex., BNHS FWF 107–113, collected from Vembanad Lake, Kerala, India (9o54'35''N, 76o20'34''E, 1–2 m a.s.l.), by Unmesh Katwate and Fibin Baby on 29.v.2014; 4 ex., WILD-14-PIS-111 to 114, collected from Edathua in the rivers flowing into the Vembanad Lake, Kerala, India (9o54'35''N, 76o20'34''E, 1–2 m a.s.l.), by Unmesh Katwate and Fibin Baby on 29.v.2014; 3 ex., ZSI- WRC-P/4092, collected from Edathua in the rivers flowing into the Vembanad Lake, Kerala, India (9o54'35''N, 76o20'34''E, 1–2 m a.s.l.), by Unmesh Katwate and Fibin Baby on 29.v.2014; 3 ex., CRG-SAC-2010.05.01-03, collected from near Cochin /Ernakulam, Kerala, by F. Baby on 18.v.2010; 6 ex. Other material: BNHS FWF 86–90, 92, Bandiwade, Gad River, Sindhudurga District, Maharashtra, collected on 15.ix.2013 by U. Katwate and S. Rane; 1 ex. BNHS FWF 91, collected from Terekhol River at Madkhol, Maharashtra, by U. Katwate, M. Paingankar and N. Dahanukar on 9.viii.2013.
Diagnosis. Pethia punctata can be distinguished from all other species of Pethia by the following combination of characters: barbels absent, lateral line complete, with 23–25 pored scales; predorsal scales 8; scales in transverse line ½4/1/3½; dorsal fin originating almost opposite to, or slightly before pelvic-fin origin; gill rakers 7 on first ceratobranchial; 4+26 total vertebrae including posterior most compound centrum; banding pattern includes a small black humeral spot covering anterior half of the fourth scale of the row below the lateral-line row; two minute dark spots below the humeral spot; a prominent spot on the caudal peduncle surrounded by a golden hoop covering scales 19–21 on the lateral-line row; and dorsal fin with 2–3 longitudinal rows of black spots, third row occupying only anterior portion of dorsal fin, extending also to 2 supernumerary and last unbranched fin rays.
Description. Photograph of topotype and Day’s material (possible syntypes) provided in Figs. 2 View FIGURE 2 A, B, C, D. Morphometric and meristic data provided in Table 1 View TABLE 1 .
Body short, deep; compressed laterally; predorsal contour almost straight or slightly convex, rising gradually to dorsal-fin origin, thereafter sloping down towards hypural notch. Ventral profile convex up to posterior end of anal-fin base, sloping gradually towards hypural notch. Caudal peduncle broad, short, its length 1.3–1.6 times its depth.
Head small, laterally compressed. Snout round, pointed, smooth, shorter than eye diameter, with a distinct lateral fold and fleshy overhanging upper lip. Eyes large, dorso-laterally positioned, closer to snout tip than margin of operculum, diameter 1.2–1.4 times interorbital width. Mouth small, subterminal, ventrally ‘U’ shaped, angle of gape not reaching to vertical from anterior margin of eye. Upper and lower lips thick, fleshy, lower lip posterior groove not interrupted. Barbels absent.
Dorsal fin originating opposite or slightly anterior to pelvic-fin origin, closer to tip of snout than to base of caudal peduncle, its distal margin concave, height greater than head length. Dorsal fin with 3 unbranched (including 2 supernumerary rays) and 8 branched rays, last unbranched ray strong, osseous, densely serrated posteriorly, with 12 (1), 14 (6), 16 (2), 17 (3) or 18 (2) serrae on its distal half, 2 (10) or 3 (4) serrae on its apical half. Pectoral fin with one unbranched and 9 (2) or 10 (12) branched rays, its tip rounded, reaching one or two scales anterior to pelvic-fin origin or sometimes marginally reaching pelvic-fin origin. Pelvic fin with one unbranched and 7 branched rays, its tip rounded, not reaching vent when adpressed. Anal fin with 3 unbranched (including 2 supernumerary rays) and 5 branched rays, its distal margin concave with rounded corners. Caudal fin forked, lobes making more than half of fin length, tips pointed. Principal branched caudal-fin rays dorsally 9 (14), ventrally 8 (14); procurrent rays dorsally 7 (14), ventrally 6 (14).
Lateral line complete with 23 (11), 24 (2) or 25 (1) pored scales in lateral series, which curves gently downwards to base of pelvic fin, then running almost straight to caudal-fin base. Scales in transverse line ½4/1/3½, predorsal scales 8, prepelvic scales 9 (13) or 10 (1), preanal scales 13 (8) or 14 (6), circumpeduncular scales 12. Pelvic axillary scale present, reaching to 1/4th of adpressed pelvic-fin length.
Osteology. Complete lateral view of the cleared and stained specimens of P. punctata (topotypic male and female) are provided in Figure 3 View FIGURE 3 A and C, respectively.
Neurocranium: Dorsal, lateral and ventral view of neurocranium provided in Figure 4 View FIGURE 4 A–C. In general appearance it is broad and wide at otic region, narrow and slopes dorsally towards ethmoid region. Ethmoid region composed of by mesethmoid, prethmoid, ethmoid, kinethmoid ( Figure 4 View FIGURE 4 A), vomer and large winged lateral ethmoid. Ethmoid region slopes down anteroventrally, meet triangular mesethmoid, which placed in between lateral wings of ethmoid portion. Lateral ethmoid contributes large portion in ethmoid complex, clearly visible, lateral processes well developed, tapers and slopes down anterolaterally, covers anterior most portion of orbital section. Prethmoid closely attached with anterior most tips of vomer. Nasals are in form of small bony plate, anterior down to the lateral ethmoid. Frontals large and contributes as largest anterior portion of neurocranium, with clear supraorbital lateral-line canal running along its lateral margin. Parietals large, with a sensory canal running on lateroposterior side forms distal margin of neurocranium. Orbitosphenoid large, distinct, attached dorsally with fontanel and ventrally with dorsovertical flap of parasphenoid. Pterosphenoid pierced, anteriorly attached with distal margin of orbitosphenoid. Parasphenoid long elongated with dorsovertical flap, attaches anteriorly with vomer and cartilaginous trabecula communis, runs posteriorly till otic region. Prootic with large auditory foramen seems to be the largest bone of the ventral surface of cranium, attached posterolaterally with anterior most part of the subtemporal fossa. Autopterotic forms posterolateral corner of the neurocranium, ventrally articulates with hyomandibular. On the ventral side of cranium, autopterotic forms the posterolateral wall of the deep subtemporal fossa. Supraoccipital attaches with the parietal on the anterior side, dorsally concave, spine well developed but does not supersedes weberian complex. Exoccipital large, well ossified, forms posterior wall of the otic capsule and the posterodorsal part of the otic bulla. Basioccipital large covers ventral portion of the otic bulla, and the articulation between the neurocranium and the first vertebra, it is a posteroventral most point of the occipital region. Basioccipital process well developed, elongated with round concave masticatory plate on the anterior side and it reaches half of weberian complex ventrally.
Hyopalatine arch and opercular series: See Figure 5 View FIGURE 5 A. The dorsal edge of the hyomandibular articulates with the autosphenotic, the autopterotic, and the prootic cranium bones and unites with the opercle in anterodorsal portion. Laterally, the hyomandibular overlaps with the posterodorsal margin of the preopercle. Opercle overlaps with interopercle and subopercle anteroventrally. The quadrate well ossified, zone of cartilage, a remnant of the palatoquadrate cartilage is less visible. Its large posteroventral process extends laterally and overlaps anterior most point of the preopercle. On the anterior side, articular condyle of the quadrate articulates strongly with the anguloarticular. The metapterygoid is well ossified, ventrally attaches with the quadrate on anterior, preopercle and hyomandibular on posterior and overlaps laterally with symplectic. Endopterygoid and ectopterygoid well ossified like metapterygoid and overlaps on dorsal and anterior face of quadrate.
Upper jaw constitutes of premaxilla and maxilla, relatively large and well ossified. Premaxilla proportionately smaller than maxilla curved anteriorly and free of any joints with maxilla and dentary, while only attached with strong ligaments. Premaxilla articulates with the dentary on posteroventral surface. The lower jaw comprises Meckel’s cartilage, dentary, anguloarticular and the coronomeckelian. Dentary articulates with maxilla on dorsolateral side, broad on posterior edge and pointed towards anterior edge. Coronomeckelian bone presents on dorsal surface of remnant of rod shape Meckel’s cartilage. Kinethmoid a small bone contributes to ethmoid complex is present in association with maxillae and premaxilla, bound with small ligaments.
Infraorbital series and scleral skeleton: See Figure 5 View FIGURE 5 B and C. Five infraorbital bones present, bearing laterosensory canal which encircles the orbital margin. Infraorbital 1 is well developed rhomboidal and wider than infraorbital 2. Infraorbital 2 with laterosensory canal appears like a thin tube, encircles orbital margin. Infraorbital 3 large among all and overlaps preopercle. Infraorbital 4 and 5 are also well developed but less wider than infraorbital 1 and 3. Supraorbital large and well ossified, covers eye dorsally. Infraorbital 6 absent.
Scleral bones well ossified, covers proximal and distal perimeter of eye sclera in lateral view, dorsal and ventral perimeter of eye is cartilaginous (Figure, 5C).
Hyoid arch: See Figure 5 View FIGURE 5 D–F. Anterior ceratohyal distinct, broad posteriorly, narrows down anteriorly. It is attached anteriorly with dorsal hypohyal and ventral hypohyal and posteriorly with posterior ceratohyal. Dorsal and ventral hypohyal are small and well ossified with no cartilaginous remnant. Large urohyal, with three well spread lateral flanges, attached anteriorly to ventral hypohyal with ligaments. Three well developed thin branchiostegal rays in each half of hyoid arch.
Branchial arches: See Figure 6A–E. Three well ossified cylindrical basibranchial, connected by congregated cartilage; first one is smaller than second and third. Three pairs of small, round, ossified hypobranchials, covered with cartilage, attaches to the respective basibranchial and ceratobranchial. First basibranchial attaches to the basihyal by ligaments; we did not identify a cartilaginous connection between basihyal and first basibranchial. Fourth and fifth ceratobranchial connects with basibranchial 3 by strong ligaments and very weak basibranchial 4 cartilage remnants. Well developed five pairs of ceratobranchials present. Ceratobranchial 1 with 7 gill rakers on anterior side and 14 on posterior. Ceratobranchial 2 with 14-15 gill rakers along their anterior margins and 17–18 on posterior. Ceratobranchial 3 with 14 gill rakers on anterior margin and 16 on posterior margin. Ceratobranchial 4 with 11 gill rakers on anterior margin and 15 on posterior margin. Ceratobranchial 5 well ossified, toothed having 13 gill rakers on anterior margin. Proximal half of ceratobranchial 5 marked with 3 rows of well ossified conical teeth. Anterior most teeth row comprises 2, middle 3 and posterior most comprises 4 conical and one small isolated tooth (Figure 6C–E). Upper gill arch constitutes by four pairs of well developed epibranchials. Epibranchial 1 broad and wing shaped while epibranchial 2, 3 and 4 are narrow bodied. We fail to locate pharyngobranchial 1, pharyngobranchial 2 and 3 are well developed, pharyngobranchial 4 cartilage very less developed and minute.
FIGURE 6. Pethia punctata , topotype, female, BNHS FWF 111, 48.5 mm SL; (A) ventral gill arches, dorsal view; (B) dorsal gill arches, ventral view; (C-E) 5th ceratobranchial teeth; (F) pectoral girdle, lateral view. Bb1-3, basibranchial 1-3; Bb4C, basibranchial 4 cartilage; Bh, basihyal; Cb1-5, ceratobranchial 1-5; Cl, cleithrum; Co, coracoid; DRC, distal radial cartilage; Eb1-4, epibranchial 1-4; Hb1-3, hypobranchial 1-3; Pb2-3, pharyngobranchial 2-3; Pb4C, pharyngobranchial 4 cartilage; Pcl, postcleithrum; PeR1–4, pectoral radial 1–4; Sc, scapula; Scl, supracleithrum.
Pectoral Girdle: See Figure 6F. Pectoral girdle position most anteriorly and does not overlap weberian apparatus laterally; articulate with posterolateral surface of neurocranium through vertically aligned supracleithrum and cleithrum. Supracleithrum small, laterally overlaps with cleithrum by articulating more than 3/ 4th area. Cleithrum large, narrow and pointed dorsally, articulates with postcleithrum lateroventrally. Postcleithrum single, elongated and well developed supports pectoral girdle posteriorly. Coracoid large, rectangular, articulates posteriorly with scapula and pectoral radials, dorsally overlaps with cleithrum. Four large well ossified pectoral radials present. Six distal radials less ossified, with three medial elements predominantly cartilaginous. Nine distal radial present, first three distal radial appear to be well ossified whereas remaining six are cartilaginous and abbreviated as distal radial cartilage, supports one unbranched and 10 branched (i. 9.i) pectoral fin rays.
Pelvic girdle: See Figure 7 View FIGURE 7 A–B. Pelvic girdle consists of a pair of thin, flat and elongated basipterygium, each supporting one unbranched and eight branched fin rays (i. 7.i). On the anterior side basipterygium bears two spinelike processes, external and internal. External process slightly elongated than internal. Five well ossified pelvic radials present, pelvic splint is well developed and laterally associated with first unbranched pelvic fin ray.
Dorsal fin: See Figure 8 View FIGURE 8 A. Dorsal fin constitutes twelve dorsal fin rays (iii. 8.i) supported by a series of pterygiophores placed between neural spines of vertebrae V8 /V9–V 14/V15. First three pterygiophores constitutes a large proximal–middle radial, articulates with two unbranched supernumerary and one serrated last unbranched dorsal fin ray, distal radial absent. Posterior to proximal–middle radial first two pterygiophores articulates with respective branched fin rays by having only distal radial, middle radials untraceable in this articulation. Middle radial appears in articulation with distal radial and dorsal base of pterygiophore on third proximal radial. All middle and distal radials are well ossified. Last tenth branched fin ray found to be closely united with ninth fin ray, whereas articulated separately with last pterygiophore, dorsal fin stay. Four free supraneurals, Sn5–8 present on anterior side of dorsal fin.
Anal fin: See Figure 8 View FIGURE 8 B. Anal fin constitutes 9 fin rays (iii. 5.i) supported by a series of anal pterygiophores placed between hemal spines of vertebrae V18 /V19–V 20/V21. First three pterygiophores constitutes long, elongated proximal–middle radial, articulates with two unbranched supernumerary and last unbranched anal fin ray, distal radial present with no traces of middle radial. Middle radial appears in articulation with distal radial and dorsal base of pterygiophore on first proximal radial. All middle and distal radials are well ossified. Last sixth branched fin ray found to be closely united with fifth anal fin ray, whereas articulated separately with last pterygiophore, anal fin stay.
Caudal fin: See Figure 9 View FIGURE 9 A–B. Seven dorsal procurrent caudal-fin rays and six ventral procurrent caudal-fin rays. Nine dorsal and eight ventral principal caudal fin rays present. Principal caudal fin rays supported by neural and hemal spines of third preural centrum, epural, pleurostyle, six hypurals and a parhypural. A single inter–hemal spine cartilage of preural centrum 4 present, placed anterior to ventral tip of hemal spine of preural centrum 3 in male and anterior to ventral tip of first hemal spine of preural centrum 2 in female. Single epural runs laterally with anterior margin of pleurostyle on dorsal half, distal tip of pleurostyle cartilaginous. Parhypural and hypural 1-6 well developed, broad and ossified. Sexual dimorphism in caudal fin skeleton is distinct in P. punctata . In male, pleural centra 2 supports single neural and hemal spine, pleural centra 3 supports well developed neural and hemal spine which support peduncle arch. In female, pleural centra 2 represents pair of neural and hemal spines, supports peduncle arch. Neural and hemal spines of third pleural centra less developed and short in female specimen as compare to male. Free uroneural absent
Weberian apparatus: See Figure 10 View FIGURE 10 . A complex structure made up by first four vertebral centra and their associated elements, covers the anterodorsal portion of swim bladder. The first centrum relatively small but distinct, possesses lateral process 1 (Lp1). Second centra equivalent in size of first, possesses large lateral process of second vertebra (Lp2). Lp2 is more developed and distinct than LP1 and grown laterally in wing shaped processes. Third and fourth centrum is much larger and each equivalent to size of first two. Scaphium and claustrum small, rounded and well ossified, present on dorsal in association with first centra. Tripus well developed, triangular grows posteriorly and overlaps inner arm of the os suspensorium, associated with third vertebral centrum. Outer arms of the os suspensorium much grown, long and reaches behind the postcleithrum bone of pectoral girdle. Neural arch 3 and neural arch 4 well developed and ossified. Supraneural 3 seems to be a second largest bone of weberian apparatus after outer arm of the os suspensorium. Supraneural 2, 3 and neural spine 4 is associated with neural arch 3 and 4. Supraneural 4 absent.
Intermuscular bone: See Figure 3 View FIGURE 3 A and C. Two series of intermuscular bones, epineural and epipleural. Total 23 epineural and 10 epipleural intermuscular bones present; first epineural appears before neural spine of seventh vertebra whereas first epipleural appears before hemal spine of sixteenth vertebra. All epipleural and epineural bifurcated at tip and becomes wider in caudal peduncle portion.
Vertebral column: See Figure 3 View FIGURE 3 A and C. 4+4 predorsal vertebrae; 4+26 total vertebrae including posterior most compound centrum, with 4+14 abdominal and 12 caudal vertebrae. Vertebral centra much elongated than deep. Neural pre and postzygopsis appears on all centra posterior to weberian complex. Hemal postzygopsis appear on all caudal vertebrae posterior to 15th centra. First hemal prezygopsis appears on 20th centra. Total 11 ribs are associated with 5th to 15th vertebrae.
Coloration in life. (Image 8a-d in Katwate et al. 2014b). Body yellow. Position of humeral and caudal spots same as described for preserved specimens, spot on caudal peduncle surrounded by a golden hoop covering scales 19–21 of lateral-line row, saddling caudal peduncle dorsally. Dorsal fin with 2–3 longitudinal rows of intense black spots, third one occupying only anterior portion of dorsal, yellow in breeding male, red and devoid of black spots in female. Pectoral, pelvic and anal fins yellow. Caudal fin yellow. Sclera bright yellow in breeding males, iridescent silver in females and immature males.
Coloration in preservative. For general appearance see Figure 2 View FIGURE 2 A–D and for illustration see Figure 10 View FIGURE 10 ; body above lateral-line scale row brown; snout, head, dorsum dark brown; lower lip, cheek, opercular region below inferior border of eye cream; ventral region uniformly cream white. Body with prominent intense black humeral spot covering anterior half of fourth scale on scale row below lateral-line row; two minute black spots below humeral spot. A prominent spot on caudal peduncle covering 19th–21st scales of lateral-line row, saddling caudal peduncle dorsally. Dorsal fin with 2–3 longitudinal rows of dark black spots, third one occupy only front portion of dorsal and also extends on 2 supernumerary and last unbranched fin rays ( Figures 1 View FIGURE 1 , 2 View FIGURE 2 A, 2C), in breeding and mature males, dorsal fin plain usually without any color bands in female ( Figure 2 View FIGURE 2 B). Anal and caudal fins hyaline, without any color bands or spots. Pectoral and pelvic fins lightly pigmented with melanophores. Base of scales studded with black melanophores.
Phylogenetic position. Phylogenetically, P. punctata from Cochin, Kerala (putative topotype), and Maharashtra (populations studied in Katwate et al. 2014b) formed a monophyletic group nested within genus Pethia ( Figure 11 View FIGURE 11 ).
Distribution. Pethia punctata is a widely distributed species in the Western Ghats of India, mainly confined to west-flowing rivers, from 8º–16ºN latitudes ( Figure 12).
Remarks. While describing the fishes of “ Cochin, on the Malabar Coast of India ”, Day (1865) described P. punctata with evident and distinguishable prominent characters. Although Day (1865) did not designate a type for P. punctata in the description, there are surviving syntypes ( Figure 1 View FIGURE 1 C, D) with collection localities. For the redescription, we use Day’s material as syntypes, with additional topotypes collected from the area encompassing the erstwhile state of Cochin, the type locality.
Day (1865), used the following combination of characters to distinguish P. punctata : no cirri; third dorsal spine strongly serrated posteriorly; lateral line first curves gently downwards, and from opposite base of ventral fin passes direct to centre of caudal fin; a black diffused spot on the twentieth and twenty-first scales of the lateral line; the anterior half of the fourth scale from the operculum, of the row next below the lateral line, deep black, and also a portion of the scale above and beneath it; fins yellowish; dorsal spotted with melanophores, in two longitudinal rows, with a third anteriorly, between the other two. Day’s (1865) lateral-line scale count of 27 for P. punctata , is apparently in error, for his syntypes and our topotypes possess only 23–25 lateral-line scales.
Species found in the Western Ghats region such as P. lutea Katwate, Katwate, Raghavan, Paingankar & Dahanukar, 2014 ; P. narayani (Hora, 1937) and P. set nai (Chhapgar & Sane, 1992) are close congeners of P. punctata in having complete lateral line. However, P. punctata differs from its Western Ghats congeners by having following combination of characters: greater number of lateral-line scales (23–25 vs. 19–22 in P. lutea , 22 in P.
narayani View in CoL , 19–21 in P. set nai ); dorsal fin with 2–3 longitudinal rows of intense black spots, third row occupying only anterior portion of fin (vs. plain and without any markings in P. lutea View in CoL , P. na r a y an i and P. setnai View in CoL ); two spots on body, one on humeral region, another on caudal peduncles (vs. three dark bands on body in P. narayani View in CoL and P. setnai View in CoL ); and prominent intense-black humeral spot covering anterior half of fourth scale on scale row below lateralline row (vs. covering third and fourth lateral line scale, extending to one scale up and down in P. lutea View in CoL ). Pethia punctata View in CoL also differs from P. narayani View in CoL by having more osseous and serrated last unbranched dorsal-fin ray (vs. last unbranched dorsal fin ray feeble and without serrations). Pethia punctata View in CoL also differs from other Indian Pethia View in CoL having a complete lateral line with a combination of characters that includes the presence of intense-black humeral spot covering the anterior half of the fourth scale on scale row below the lateral-line row (vs. absent in P. expletiforis View in CoL and inconspicuous on the scale row below third and fourth scales of lateral-line row in P. rutila View in CoL ); 23–25 lateral-line scales (vs. 21–22 scales in P. rutila View in CoL ); ½4/1/3½ scales in transverse line on body (vs. ½5/1/5½ in P. stoliczkana View in CoL ) ( Dishma & Vishwanath, 2013; Laltlanhlua et al. 2014; Day 1871; Linthoingambi & Vishwanath, 2007). Pethia punctata View in CoL differs from P. t i ct o by having a complete lateral line (vs. incomplete lateral line) ( Hamilton, 1822). A more detailed diagnosis for separating P. punctata View in CoL from P. t i c t o will appear in a forthcoming study clarifying the identity of P. t icto (Katwate et al., in prep.).
The distribution, habitat and ecology of P. punctata View in CoL has been discussed in detail in Katwate et al. (2014b).While comparing the type material and topotypic specimens of P. muvattupuzhaensis View in CoL , we observed that the characters suggested by Beevi & Ramachandran (2005) to distinguish it from P. punctata View in CoL are invalid: the two nominal species are indistinguishable in all respects. Further details on the synonymy of P. muvattupuzhaensis View in CoL are discussed below.
Synonymy of Pethia muvattupuzhaensis View in CoL . Pethia muvattupuzhaensis View in CoL was described based on the holotype ( Figure 13 View FIGURE 13 ) and four paratypes ( Beevi & Ramachandran 2005). However, out of the five type specimens of P. muvattupuzhaensis View in CoL in SRC-ZSI, only three belong to the genus Pethia View in CoL , while two represent another cyprinid genus ( Figure 14 View FIGURE 14 ). We therefore studied only the holotype and the two paratypes that are conspecific with it. Comparison of type series ( Figure 13 View FIGURE 13 ) and recent collection of topotypic specimens of P. muvattupuzhaensis View in CoL ( Figure 1 View FIGURE 1 E, F) and P. punctata View in CoL ( Figure 1 View FIGURE 1 A, B) revealed no significant morphological or meristic differences between the two nominal species.
Beevi & Ramachandran (2005) distinguished P. muvattupuzhaensis View in CoL from P. punctata View in CoL based on two prominent characters, namely: fewer lateral transverse scales rows (2½–3 vs. 4) and dorsal fin devoid of rows of spots (vs. present). However, examination of the relevant type specimens of P. muvattupuzhaensis View in CoL and fresh topotypic specimens shows that there are ½4 scales between the dorsal-fin origin and lateral-line scale row, and 3½ scales between lateral-line row and the pelvic-fin origin ( Table 1 View TABLE 1 ). The transverse scale count of P. muvattupuzhaensis View in CoL matches exactly with that of putative syntypes (MCZ 4303 and BMNH 1889.2.1.755, coll. F. Day) ( Figure 4 View FIGURE 4 C, D) as well as topotypic specimens ( Figure 4 View FIGURE 4 E, F) of P. punctata View in CoL collected from near Cochin, Kerala, and additional specimens from other rivers draining the Western Ghats ( Katwate et al. 2014). Further, Beevi & Ramachandran (2005) mention 2½–3 scales between the lateral-line row and the pelvic-fin origin. However, the illustration provided by them in the description as well as our own examination of the holotype ( Figure 13 View FIGURE 13 ) indicates the presence of 3½ scales, which is the same as that in P. punctata View in CoL ( Table 1 View TABLE 1 ).
Beevi & Ramachandran (2005) also distinguished P. muvattupuzhaensis View in CoL from P. punctata View in CoL by the absence of rows of dark spots on the dorsal fin. The specimens in the type series of P. muvattupuzhaensis View in CoL , however, are female individuals. The rows of black spots on the dorsal fin of P. punctata View in CoL are in fact a sexually dichromatic: only mature and breeding males possess these, whereas females are usually devoid of this character and have an unmarked dorsal fin ( Katwate et al. 2014b). Recent collections of topotypes of P. muvattupuzhaensis View in CoL ( Figure 1 View FIGURE 1 E, F) and P. punctata View in CoL ( Figure 1 View FIGURE 1 A, B) support these characters. By including this sexually dichromatic character, male individuals of topotypic P. muvattupuzhaensis View in CoL key out as P. punctata View in CoL in the key provided by Beevi & Ramachandran (2005).
Beevi & Ramachandran (2005) also distinguished P. muvattupuzhaensis View in CoL from P. punctata View in CoL by morphometric characters such as a more elongated body, shallower body depth, and narrower caudal peduncle. A comparison of the morphometric and meristic values of the type series and topotypic specimens of P. muvattupuzhaensis View in CoL with P. punctata View in CoL ( Table 1 View TABLE 1 ) do not reveal any such difference. Furthermore, we found that no significant osteological differences separate P. muvattupuzhaensis View in CoL as distinct from P. punctata View in CoL (see Figure 2 View FIGURE 2 A −C).
Morphometric data was multivariate normal (Doornik & Hansen omnibus Ep = 33.54, P = 0.7548). There was no significant difference in the multivariate morphometry of the two species (MANOVA: Pillai’s trace = 0.7494, F12,20 = 1.794, P = 0.1498). Genetically, topotypes of P. muvattupuzhaensis View in CoL and P. punctata View in CoL , as well as additional P. punctata View in CoL populations from Maharashtra were not significantly different ( Table 2 View TABLE 2 , Figure 11 View FIGURE 11 ), forming a monophyletic group supported by high bootstrap value (99%).
We conclude that the characters suggested by Beevi & Ramachandran (2005) to differentiate P. muvattupuzhaensis View in CoL from P. punctata View in CoL are invalid: the two “species” are indistinguishable in external morphology, osteology and in the gene fragments investigated. We consider Pethia muvattupuzhaensis View in CoL , therefore, to be a junior synonym of P. punctata View in CoL .
Material examined. Pethia muvattupuzhaensis (n = 19): Holotype, F. 12241, ZSI / WGFRS, CLT, collected from Muvattupuzha river, Ooramana, Ernakulam District, Kerala, India, by K.S. Jameela Beevi & A. Ramachandran on 21.xi.2001; Paratypes, 2 ex., (uncatalogued specimens), ZSI / WGFRS, CLT, data same as holotype; 7 ex., BNHS FWF 114 to 120, collected from Muvattupuzha River, Ooramana, Ernakulam District, Kerala, India (9o57'56''N, 76o31'17''E, 19 m a.s.l.), by Unmesh Katwate and Fibin Baby on 31.v.2014; 5 ex., WILD- 14-PIS-106 to 110, collected from Muvattupuzha River, Ooramana, Ernakulam District, Kerala, India (9o57'56''N, 76o31'17''E, 19 m a.s.l.), by Unmesh Katwate and Fibin Baby on 31.v.2014; 4 ex., ZSI-WRC-P/4091, collected from Muvattupuzha River, Ooramana, Ernakulam District, Kerala, India (9o57'56''N, 76o31'17''E, 19 m a.s.l.), by Unmesh Katwate and Fibin Baby on 31.v.2014.
Pethia lutea (n= 22): Holotype, BNHS FWF 71, collected from Bhira (18.441o N & 73.267o E, elevation 50m), Kundalika River, Raigad District, Maharashtra, India by Unmesh Katwate and Chetana Katwate on 23.xii.2012; Paratypes, 3 exs., BNHS FWF 72, 78 and 79, collected from Bhira (18.441o N & 73.267o E, 50 m), Kundalika River, Raigad District, Maharashtra, India by Unmesh Katwate and Chetana Katwate on 23.xii.2012; 1 ex., WILD-14- PIS-061, collected from Bhira (18.441o N & 73.267o E, 50m), Kundalika River, Raigad District, Maharashtra, India by Unmesh Katwate and Chetana Katwate on 23.xii.2012; 1 ex., ZSI-WRC-P/ 3686, collected from Bhira (18.441o N & 73.267o E, 50m), Kundalika River, Raigad District, Maharashtra, India by Unmesh Katwate and Chetana Katwate on 23.xii.2012; 3 exs., BNHS FWF 73, 80 and 81, collected from Karjat (18.922o N & 73.332o E, 48m), Ulhas River, Raigad District, Maharashtra, India by Neelesh Dahanukar and M. Paingankar on 23.vi.2012; 2 exs., BNHS FWF 74 and 82, collected from Mangaon (18.233o N & 73.256o E, 7 m), Kal River - tributary of Savitri River, Raigad District, Maharashtra, India by Unmesh Katwate and Chetana Katwate on 05.i.2013; 1 ex., BNHS FWF 75, collected from Mahad (18.091o N & 73.466o E, 16 m), Savitri River, Raigad District, Maharashtra, India by Unmesh Katwate, Chetana Katwate, Rajendra Pawar and Vishwas Shinde on 23.ix.2013; 1 ex., WILD-14-PIS-062, collected from Mahad (18.091o N & 73.466o E, 16 m), Savitri River, Raigad District, Maharashtra, India by Unmesh Katwate, Chetana Katwate, Rajendra Pawar and Vishwas Shinde on 23.ix.2013; 1 ex., ZSI-WRC-P/3687, collected from Mahad (18.091o N & 73.466o E, 16 m), Savitri River, Raigad District, Maharashtra, India by Unmesh Katwate, Chetana Katwate, Rajendra Pawar and Vishwas Shinde on 23.ix.2013; 1 ex., BNHS FWF 76, collected from Shivathar Ghal (18.148o N & 73.619o E, 145 m), Savitri River, Raigad District, Maharashtra, India by Unmesh Katwate, Chetana Katwate, Rajendra Pawar and Vishwas Shinde on 26.xi.2013; 1 ex., WILD-14-PIS-063, collected from Shivathar Ghal (18.148o N & 73.619o E, 145 m), Savitri River, Raigad District, Maharashtra, India by Unmesh Katwate, Chetana Katwate, Rajendra Pawar and Vishwas Shinde on 26.xi.2013; 1 ex., ZSI-WRC-P/3688, collected from Shivathar Ghal (18.148o N & 73.619o E, 145 m), Savitri River, Raigad District, Maharashtra, India by Unmesh Katwate, Chetana Katwate, Rajendra Pawar and Vishwas Shinde on 26.xi.2013; 2 exs., BNHS FWF 83 and 84, collected from Poladpur (17.983o N & 73.470o E, 34 m), Savitri River, Raigad District, Maharashtra, India by Unmesh Katwate and Chetana Katwate on 27.xi.2013; 2 exs., BNHS FWF 77 and 85, collected from Sangameshwar (17.187o N & 73.550o E, 12 m), Shastri River, Ratnagiri District, Maharashtra, India by Unmesh Katwate and Saurabh Rane on 16.ix.2013; 1 ex., WILD- 14-PIS- 0 64, collected from Sangameshwar (17.187o N & 73.550o E, 12 m), Shastri River, Ratnagiri District, Maharashtra, India by Unmesh Katwate and Saurabh Rane on 16.ix.2013.
Pethia setnai (n = 35): Holotype, ZSI-K FF2766, collected from Sanguem, Goa, by S. R. Sane on 1.iii.1985; Paratypes, 6 ex., ZSI-K FF2767, collected from Sanguem, Goa, by S. R. Sane on 1.iii.1985; 9 ex., BNHS FWF 53, 63 to 70, collected from Sanguem, Goa, by U. Katwate, M. Paingankar and N. Dahanukar on 10.viii.2013; 3 ex.; WILD-13-PIS-043 to 0 45, collected from Sanguem, Goa, by U. Katwate, M. Paingankar and N. Dahanukar on 10.viii.2013; 2 ex., ZSI-WRC-P/3567, collected from Sanguem, Goa, by U. Katwate, M. Paingankar and N. Dahanukar on 10.viii.2013; 9 ex., BNHS FWF 54 to 62, collected from Terekhol River at Madkhol, Maharashtra, by U. Katwate and N. Dahanukar on 12.vi.2013; 3 ex., WILD-13-PIS-046 to 48, collected from Terekhol River at Madkhol, Maharashtra, by U. Katwate and N. Dahanukar on 12.vi.2013; 2 ex., ZSI-WRC-P/3568, collected from Terekhol River at Madkhol, Maharashtra, by U. Katwate and N. Dahanukar on 12.vi.2013. Osteological details were obtained from Katwate et al. (2013).
Pethia narayani (n = 2): Syntypes, 2 ex., ZSI-K F12180/1, collected from Cauvery River, Coorg, by C.R.N. Rao (only photographs examined).
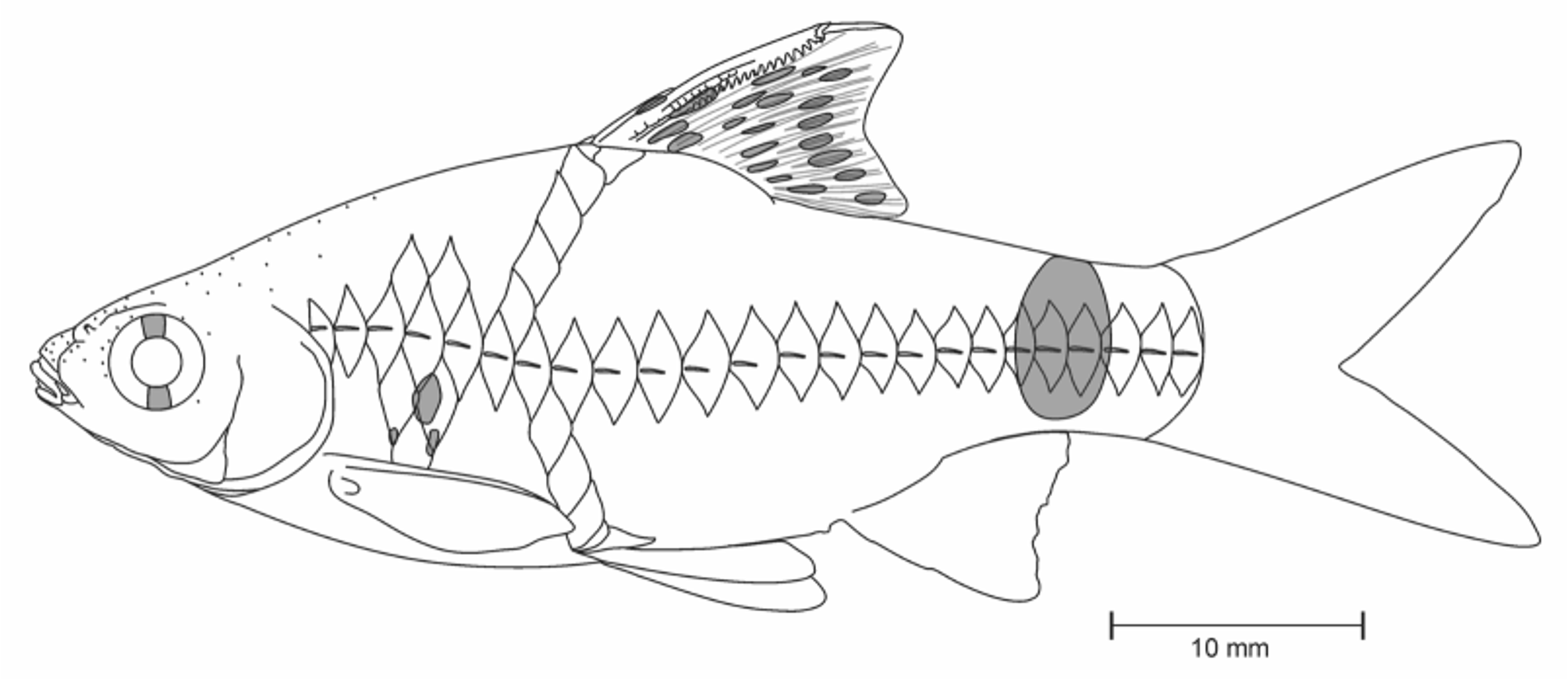

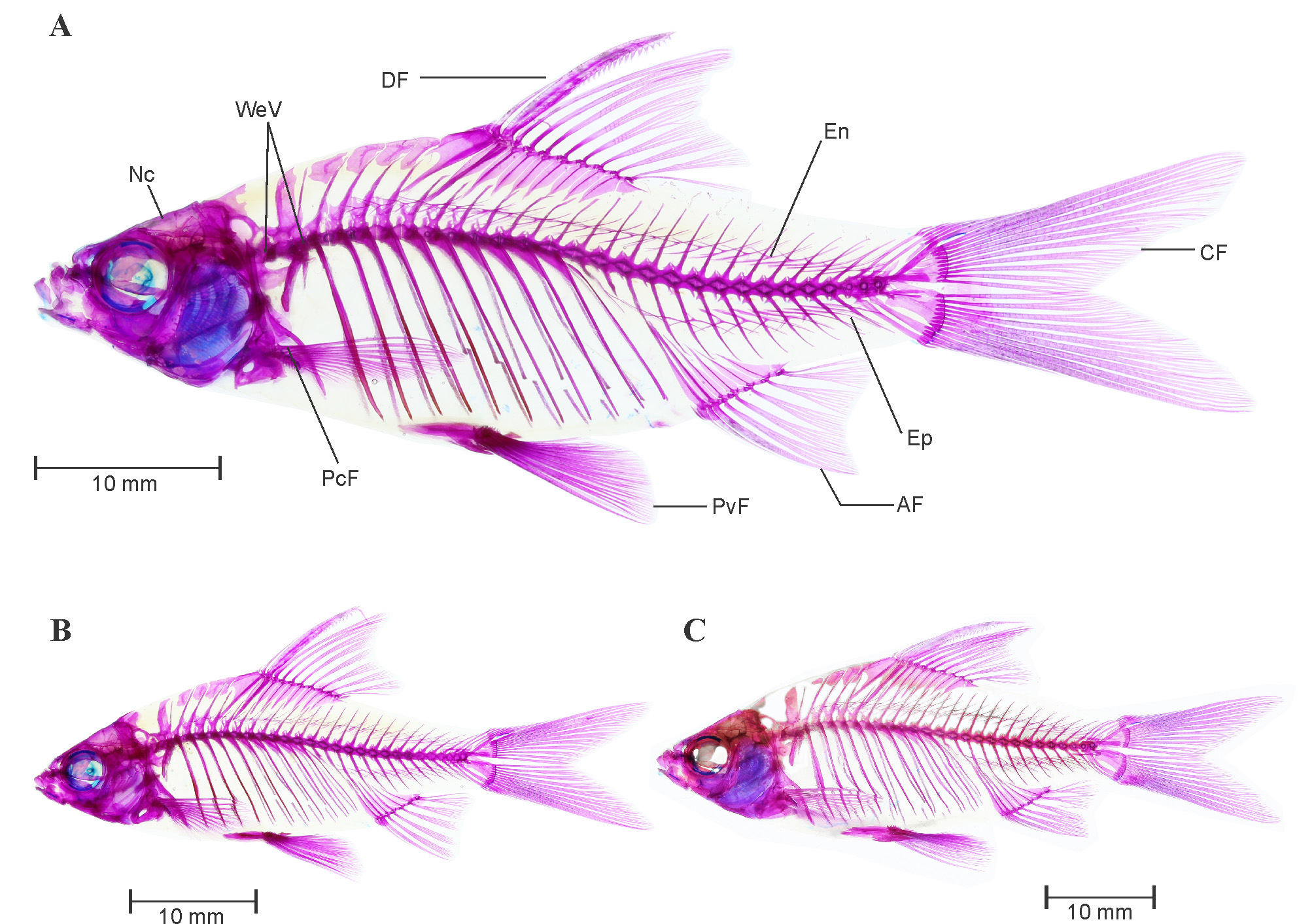
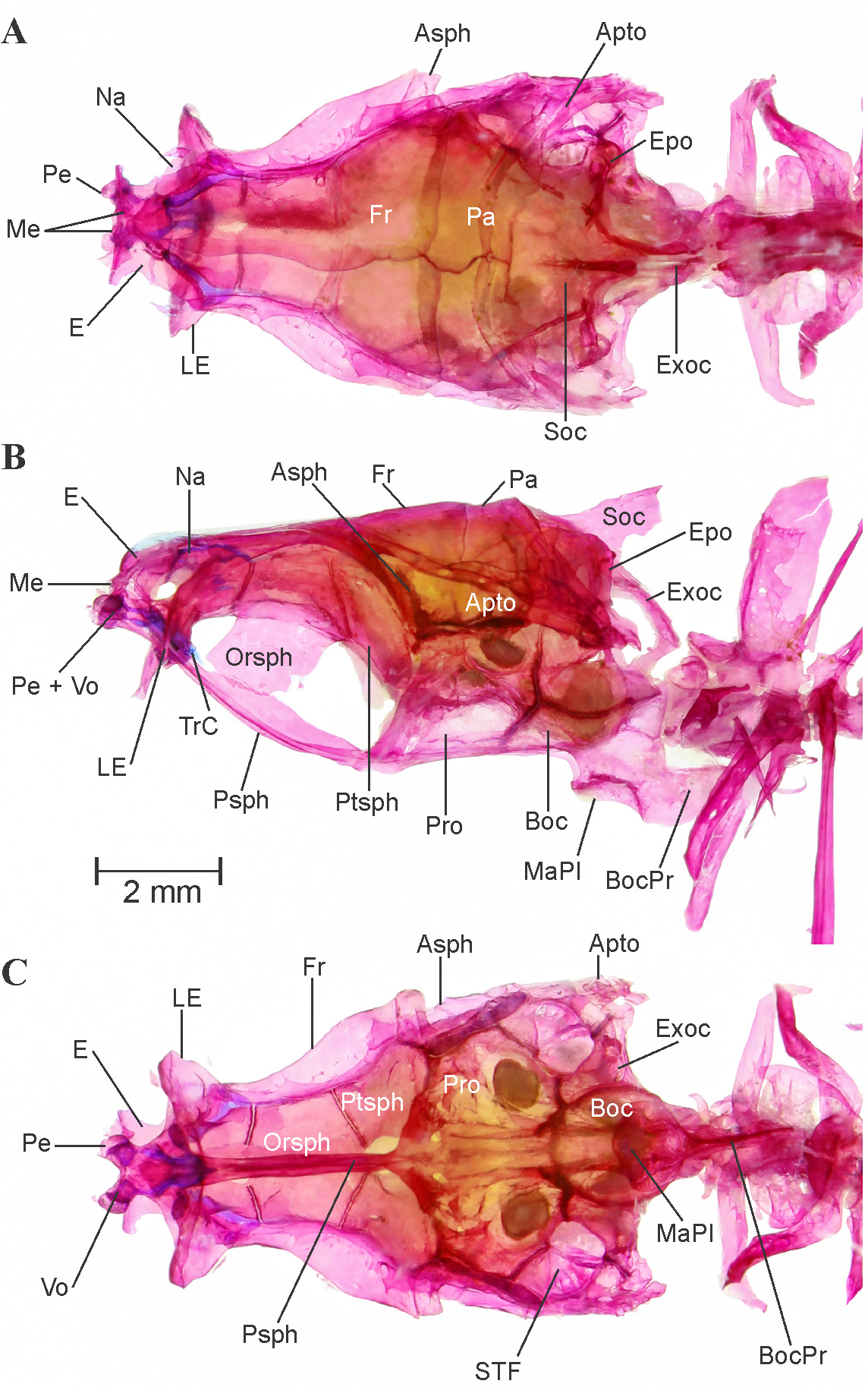
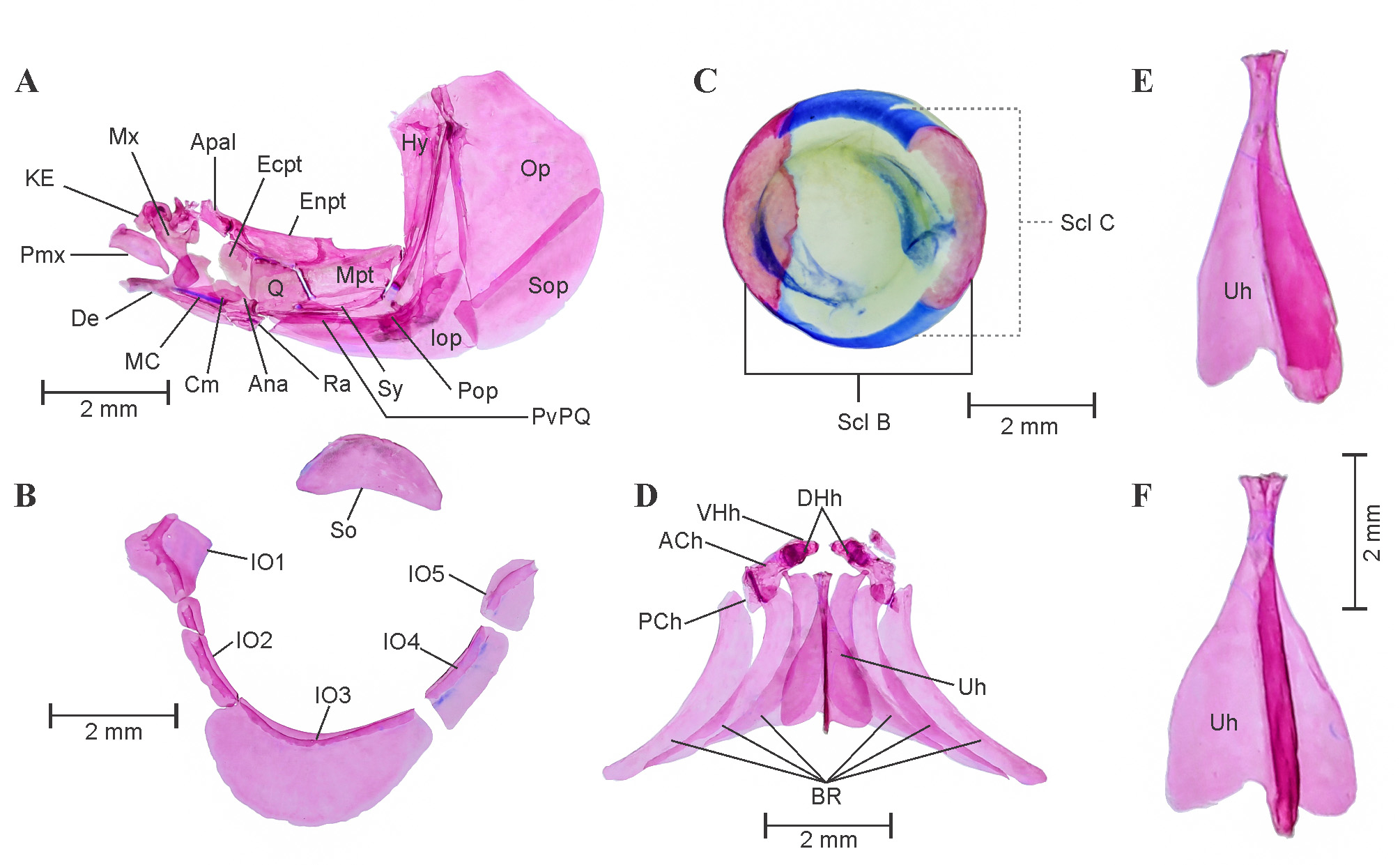
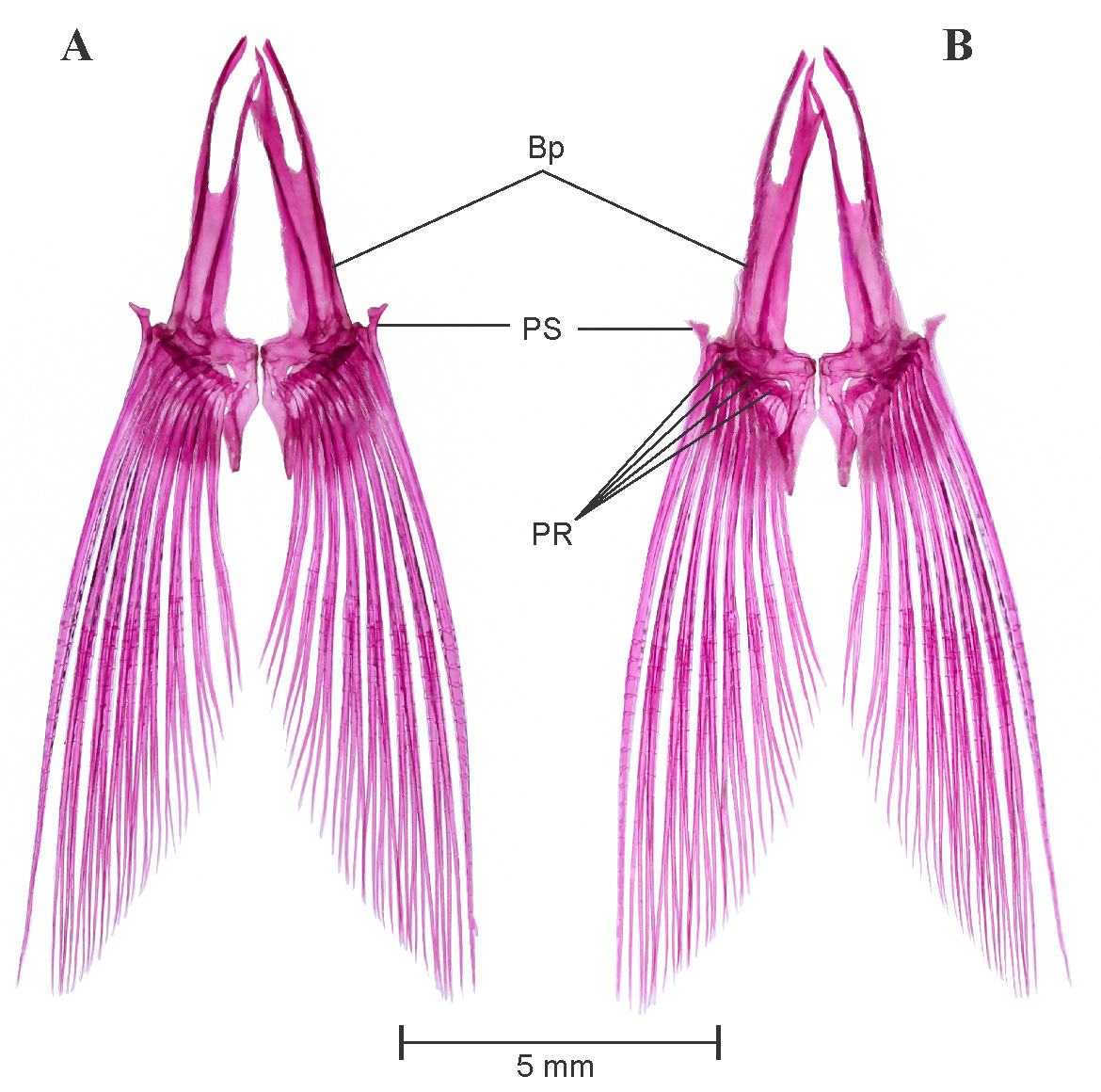
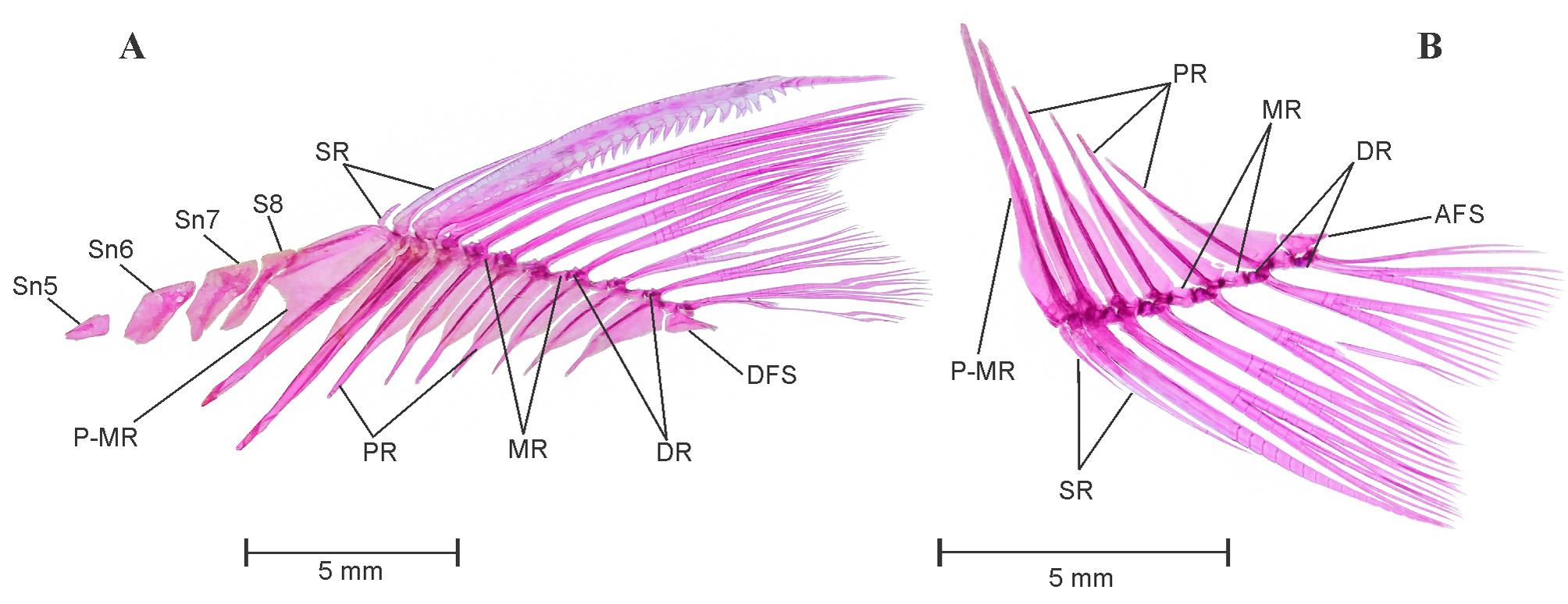
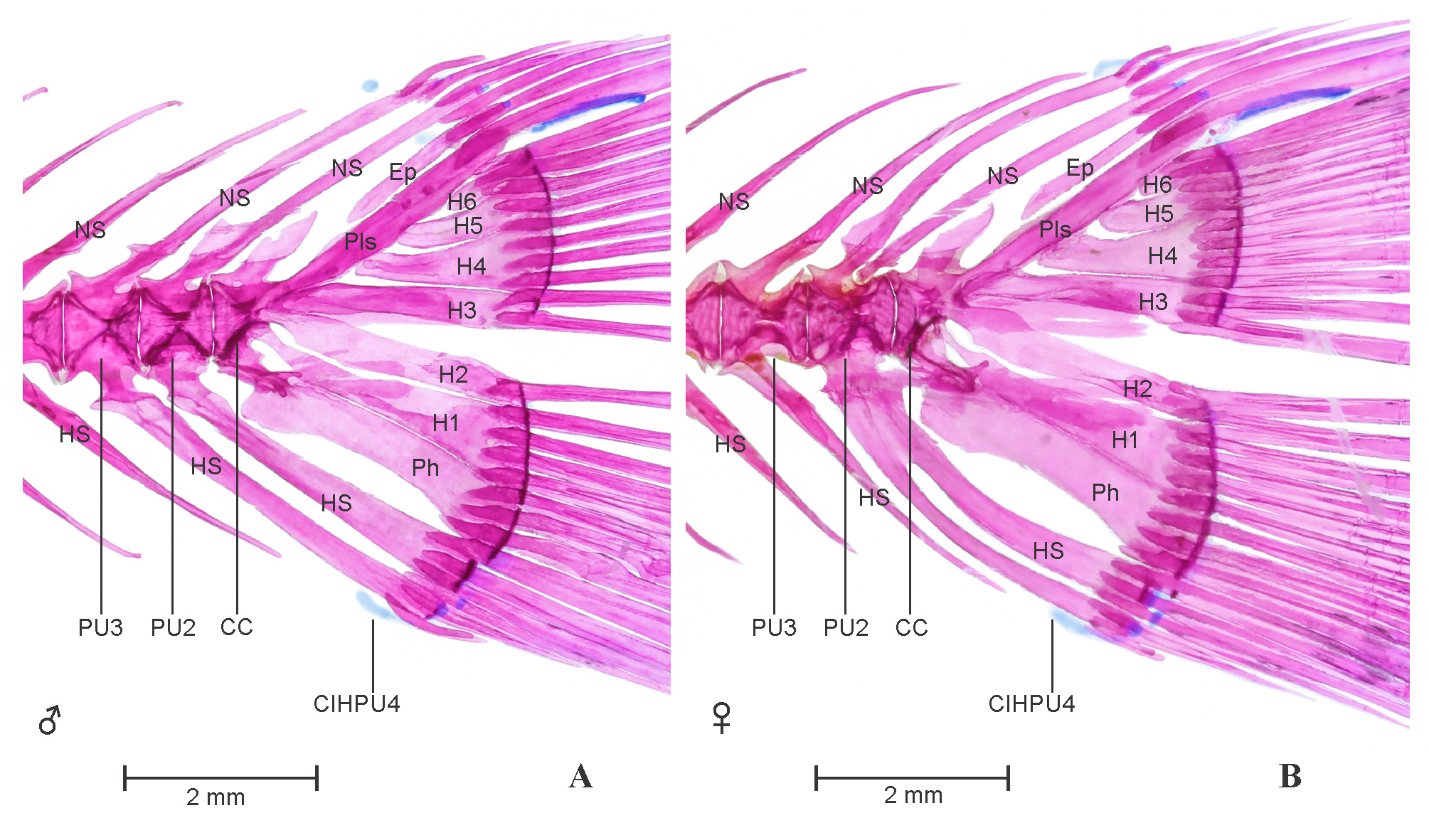
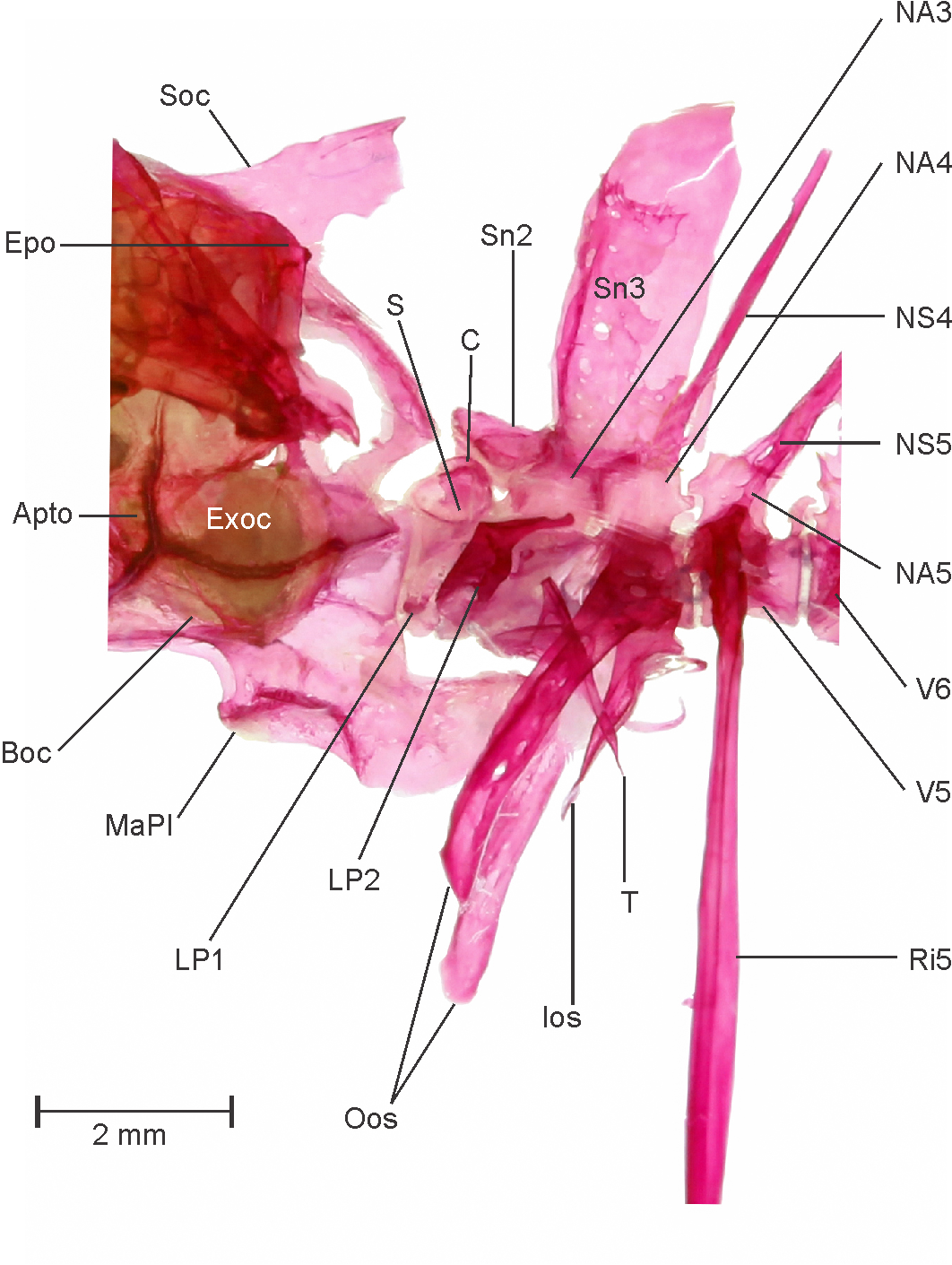
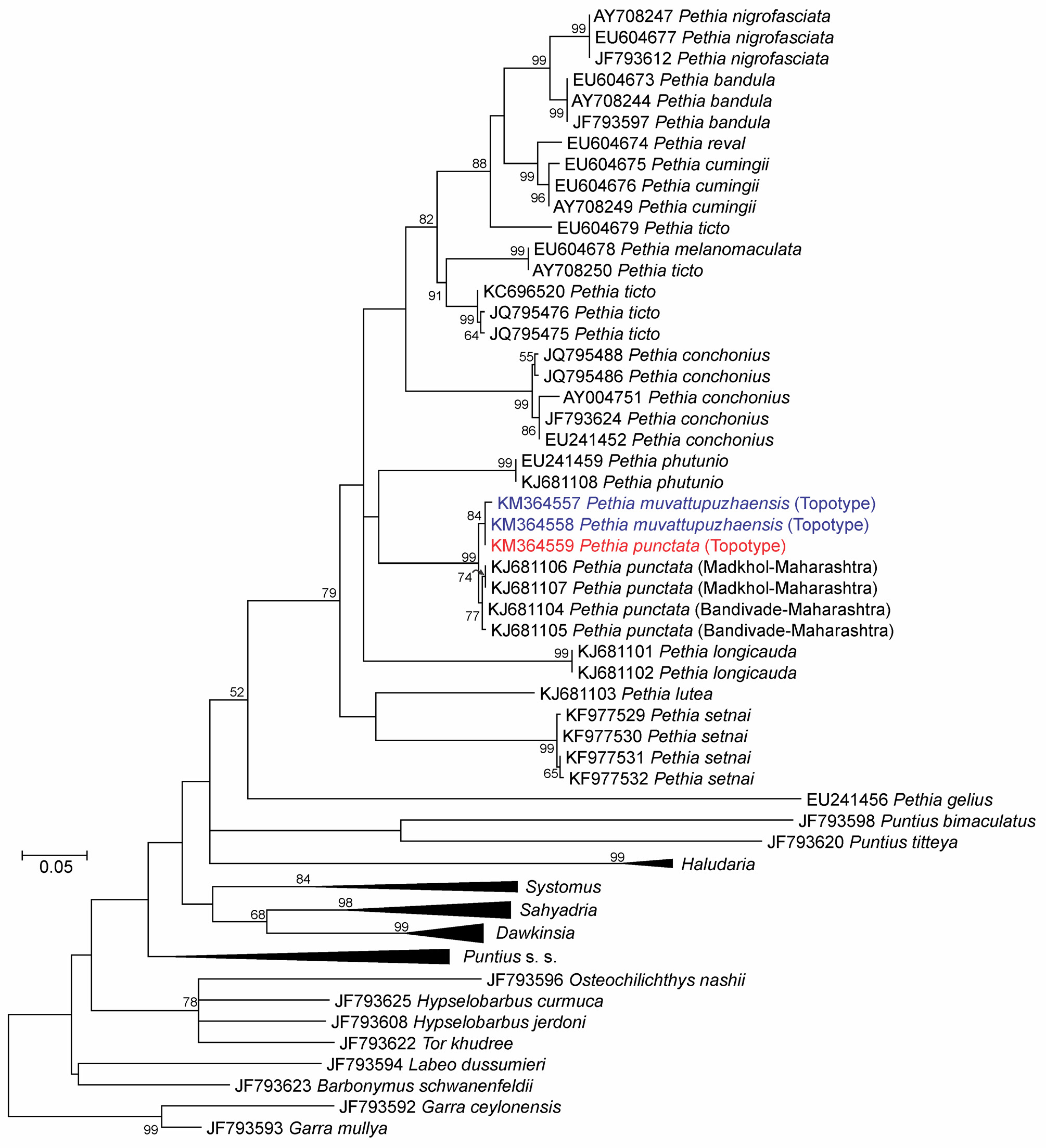


TABLE 1. Morphometric data of Pethia punctata, topotypic material (n = 14; BNHS FWF 107 – 113, WILD- 14 - PIS- 111 to 114, ZSI-WRC-P / 4092) and Pethia muvattupuzhaensis type series and topotypic material (n = 19; type series: Holotype, F. 12241, ZSI / WGFRS, CLT; Paratypes, 2 ex., (uncatalogued specimens), ZSI / WGFRS, CLT; topotypic material: BNHS FWF 114 to 120, WILD- 14 - PIS- 106 to 110, ZSI-WRC-P / 4091).
| Species | Pethia punctata | Pethia muvattupuzhaensis |
|---|---|---|
| Mean (s.d.) Range | Mean (s.d.) Range | |
| Morphometric | ||
| Total Length (mm) | 63.7 (4.2) 58.6–71.7 | 54.0 (3.3) 48.3–59.9 |
| Standard Length (SL, mm) | 49.9 (3.4) 44.8–56.1 | 41.7 (2.9) 37.3–46.6 |
| %SL | ||
| Head length (HL) | 27.0 (1.4) 25.1–29.9 | 26.7 (1.1) 25.1–28.8 |
| Body depth | 36.0 (1.2) 34.7–38.7 | 37.3 (2.0) 34.2–42.3 |
| Body width at dorsal-fin origin | 16.2 (1.5) 12.5–17.9 | 15.3 (2.6) 9.6–17.9 |
| Body width at anal-fin origin | 11.9 (1.7) 7.8–13.9 | 10.4 (1.8) 6.6–12.3 |
| Pre-dorsal distance | 48.8 (1.7) 46.1–51.4 | 49.1 (1.6) 46.3–51.9 |
| Dorsal to hypural distance | 54.2 (1.8) 50.3–56.5 | 54.4 (1.6) 51.0–57.3 |
| Prepelvic distance | 48.1 (1.8) 44.6–51.2 | 48.2 (1.8) 45.0–52.7 |
| Preanal distance | 70.8 (2.0) 67.0–74.7 | 71.5 (1.9) 68.8–75.4 |
| Prepectoral distance | 26.0 (1.5) 23.6–29.2 | 25.7 (1.2) 24.2–29.1 |
| Dorsal-fin length | 28.5 (1.6) 25.8–32.5 | 28.0 (1.5) 24.7–31.1 |
| Dorsal-fin spine length | 22.2 (1.7) 19.0–25.2 | 22.0 (1.5) 19.4–24.2 |
| Length of dorsal-fin base | 17.0 (1.4) 15.1–19.1 | 17.2 (1.3) 15.4–20.3 |
| Pectoral-fin length | 20.4 (1.0) 18.8–22.6 | 20.3 (1.7) 18.0–23.1 |
| Anal-fin depth | 17.4 (0.9) 15.7–18.9 | 16.6 (1.2) 14.5–19.5 |
| Caudal-peduncle length | 19.4 (1.5) 17.3–22.1 | 18.7 (1.5) 15.7–21.5 |
| Caudal-peduncle depth | 14.1 (0.4) 13.3–14.9 | 14.0 (0.7) 13.1–15.3 |
| %HL | ||
| Snout length | 27.3 (2.5) 22.3–32.5 | 25.2 (2.5) 21.3–28.9 |
| Eye diameter | 30.4 (2.3) 27.6–33.6 | 33.4 (3.0) 28.5–38.2 |
| Inter-orbital width | 38.1 (2.3) 34.6–42.1 | 39.9 (2.6) 34.8–44.1 |
| Head depth | 77.0 (3.9) 69.6–81.2 | 77.3 (3.3) 72.6–83.0 |
| Head width | 54.5 (3.1) 47.8–59.9 | 54.3 (2.1) 50.5–60.0 |
| Meristics | ||
| Lateral-line scale | 23−25 | 23−25 |
| Number of lateral-line pores | 23−25 | 23−25 |
| Dorsal-fin spine serrae | 12−18 | 11−18 |
| Transverse scale rows | ½4/1/3½ | ½4/1/3½ |
| Predorsal scale | 8 | 8 |
| Prepelvic scale | 9−10 | 9−10 |
| Preanal scale | 13−14 | 13−14 |
| Circumpeduncular scales | 12 | 12 |
| Dorsal-fin rays | iii 8 | iii 8 |
| Pectoral-fin rays | i 9−10 | i 9−10 |
| Pelvic-fin rays | i 7 | i 7 |
| Anal-fin rays | iii 5 | iii 5 |
| Caudal-fin rays (procurrent) | 7+6 | 7+6 |
| Caudal-fin rays (principal) | 9+8 | 9+8 |
TABLE 2. Pairwise percent distances in cytochrome b (cytb) gene sequence (below the diagonal) and standard errors (above the diagonal) between the topotypic and widely distributed Pethia punctata and topotypic P. muvattupuzhaensis based on Tamura & Nei (1993) nucleotide substitution model with gamma distribution and 1000 bootstrap iterations.
| Sr. No. | Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|---|
| 1 | KM364559 View Materials Pethia punctata (Topotype) | 0.41 | 0.41 | 0.36 | 0.42 | 0.28 | 0.00 | |
| 2 | KJ681106 View Materials P. punctata (Madkhol, Maharashtra) | 0.77 | 0.00 | 0.20 | 0.29 | 0.53 | 0.41 | |
| 3 | KJ681107 View Materials P. punctata (Madkhol, Maharashtra) | 0.77 | 0.00 | 0.20 | 0.29 | 0.53 | 0.41 | |
| 4 | KJ681104 View Materials P. punctata (Bandivade, Maharashtra) | 0.58 | 0.19 | 0.19 | 0.20 | 0.47 | 0.36 | |
| 5 | KJ681105 View Materials P. punctata (Bandivade, Maharashtra) | 0.77 | 0.39 | 0.39 | 0.19 | 0.53 | 0.42 | |
| 6 | KM364557 View Materials Pethia muvattupuzhaensis (topotype) | 0.39 | 1.17 | 1.17 | 0.97 | 1.17 | 0.28 | |
| 7 | KM364558 View Materials P. muvattupuzhaensis (topotype) | 0.00 | 0.77 | 0.77 | 0.58 | 0.77 | 0.39 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pethia punctata ( Day, 1865 )
| Katwate, Unmesh, Baby, Fibin, Raghavan, Rajeev & Dahanukar, Neelesh 2014 |
Puntius muvattupuzhaensis
| Beevi & Ramachandran 2005 |
Pethia muvattupuzhaensis (
| Beevi & Ramachandran 2005 |
Puntius punctatus
| Day 1865 |
Barbus punctatus (
| Day 1865 |
Puntius ticto punctatus
| Day 1865 |