
Crocidura yaldeni, Lavrenchenko, Leonid A., Voyta, Leonid L. & Hutterer, Rainer, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4196.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:42261121-4F80-4D02-96DE-77AD1CD912C1 |
|
DOI |
https://doi.org/10.5281/zenodo.5614915 |
|
persistent identifier |
https://treatment.plazi.org/id/F4813A6D-1B58-4E0F-B02C-FF4D9FF69F55 |
|
taxon LSID |
lsid:zoobank.org:act:F4813A6D-1B58-4E0F-B02C-FF4D9FF69F55 |
|
treatment provided by |
Plazi |
|
scientific name |
Crocidura yaldeni |
| status |
sp. nov. |
Crocidura yaldeni sp. nov.
Figs. 7 View FIGURE 7 A, 8A, 9A, 10; Table 2 View TABLE 2 .
Crocidura View in CoL sp. B: Bannikova et al., 2001: 56.
Crocidura View in CoL sp. B: Bannikova et al., 2005: 47.
Crocidura View in CoL sp. B: Lavrenchenko et al., 2009: 57.
Holotype. ZMMU S-165342; adult male, dry skin and skull; collector's number 30; collected by L.A. Lavrenchenko on 16 April 1997.
Type locality. Beletta Forest , south-western Ethiopia, 07º34'N, 036º31'E, 1900 m a.s.l. GoogleMaps
Paratypes. ZMMU S-165340 (adult female, dry skin and skull, collector's number 20); ZMMU S-165341 (adult male, dry skin and skull, collector's number 28); ZMMU S-165343 (adult male, dry skin and skull, collector's number 35; Fig. 8 View FIGURE 8 A); all three specimens from the type locality collected by L.A. Lavrenchenko between 13 and 19 April 1997.
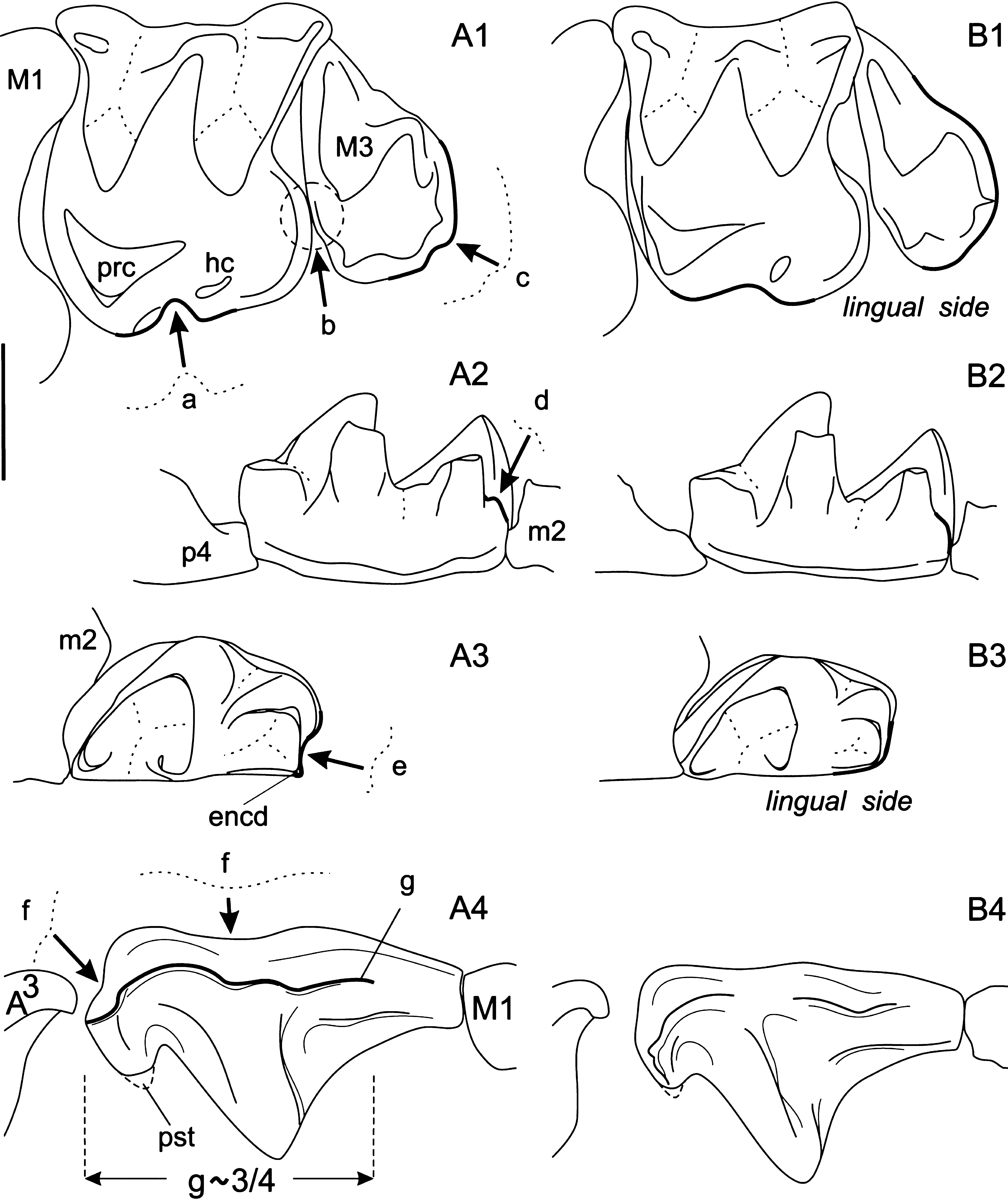
Diagnosis. A large-sized, greyish-brown Crocidura similar in external measurements (HB, TL) to C. thalia but slightly larger, on average, in cranial size ( Table 2 View TABLE 2 ). Distinguished from the latter species by the following features: moderately longer hindfoot and claws; relatively uniformly colored tail; bristle hairs of tail bicolored and longer; skull large; nasal aperture wide; dorsal profile of skull looks concave at the midpoint and, slightly convex above the upper tooth row ( Fig. 8 View FIGURE 8 , A); upper first incisor with a hook-like apex; lingual outline of the second upper molar with a distinct symmetric incision ( Fig. 9 View FIGURE 9 , A1-a); second and third upper molar in firmly contact ( Fig. 9 View FIGURE 9 , A1-b); occlusal outline of the third upper molar more undulated ( Fig. 9 View FIGURE 9 , A1-c); hypoconulids of the lower molars visible in medial view ( Fig 9 View FIGURE 9 , A2-d); postcingulid of m3 forming a notch ( Fig. 9 View FIGURE 9 , A3-e).
Description. Large-sized Crocidura (head and body length 84.0– 99.5 mm; here regarded as large-sized shrew in comparison to examined species) with a moderately long tail, ranging between 62.2–77.6% of head and body length. Dorsal pelage grey-brown with pale ochre shades (differs from C. thalia in slightly less intensive shades); dorsal hairs grey at base, brown at tip. Ventral pelage blackish-grey with pale ochre wash; ventral hairs dark grey at base, pale-yellowish at tip. Dorsal surface of fore- and hindfoot brownish. Tail uniformly colored, dark grey-brown above and brown below. Bristle hairs are long, dark-grey at base, pale-grey at tip, and present along the full length of the tail.
Skull ( Fig. 8 View FIGURE 8 , A) with a long rostrum and wide braincase. The rostral part is distinctly inflated (similar to C. thalia and differs from others Ethiopian endemic and Afromontane-Afroalpine species). The nasal aperture is wide; the posterior margin of the aperture has no medial tip. The dorsal outline of the orbital parts is slightly flexed. The lateral profile of the braincase is slightly angulated ( Fig. 8 View FIGURE 8 , A-a). The nuchal crests are well-developed; their postero-lateral ends are prolonged and joint with noticeable paraoccipital processes ( Fig. 8 View FIGURE 8 , A-pp). The sagittal crest is faint (in adult specimens); the temporal line is clearly recognizable. Mandible with broad body; its lower margin running in a shallow convex inward curve; body achieving its maximum depth below p4–m3. Coronoid spicule massive. External temporal fossa well-developed and wide. Angular process with hooked tip ( Fig. 8 View FIGURE 8 , A).
First upper incisor (I1) is robust; the apex (partly worn) is comparably long and hooked. The talon is welldeveloped, with distinct lingual ridge (lingual cingulum). The posterior ridge of the talon apex is stronglydeveloped. Lateral cingulum is weak and extending dorsally about 3/4 of I1 base height, with a faint bulge on the inferior part. Upper antemolars (A1–A3) are well-spaced, with a weak buccal cingulum and postero-buccal small cuspules. The upper first antemolar (A1) is very large; basal prominence of the anterior ridge is absent. The upper second antemolar (A2) is smallest, approximately 1/3 the height of A1, and 3/4 the height of A3. The upper third antemolar (A3) with straight buccal outline; the posterior edge is not strongly curved and connected with P4. The parastyle of the fourth upper premolar (P4) is well defined (tip partly worn); the upper margin (lateral view) is undulated and distinctly concave above parastyle and main cusp ( Fig. 9 View FIGURE 9 , A4-f); three-quarters of the crown base is surrounded by a cingulum ( Fig. 9 View FIGURE 9 , A4-g). The upper molars (M1–M2) are comparably wide. The lingual edge of the upper second molar (M2) has a well-developed and symmetrically curved incision ( Fig. 9 View FIGURE 9 , A1-a). The upper third molar (M3) is relatively long and wide, in close contact with M2 ( Fig. 9 View FIGURE 9 , A1-b), posterior edge is undulated ( Fig. 9 View FIGURE 9 , A1-c).
The first lower incisor (i1) is wide at base, with a blunt tip; the tooth is partly worn in all specimens, and therefore the denticulation of the cutting ridge is unknown. The second incisor (a1) is elongated, approximately 1/3 of the lower border of the tooth is in contact with i1; its posterior border is considerably (about 1/4) overlapped by the fourth lower premolar (p4); the postero-lateral ridge has a bulge. The basal prominence on the anterior ridge of p4 is present but indistinct, the lingual cusp (metaconid) is small. The hypoconulids of the lower molars (m1-m3) are well-developed and visible from the medial side of the teeth ( Fig. 9 View FIGURE 9 , A2-d). The postcingulid of the lower third molar (m3) forms a visible notch buccal to the entoconid ( Fig. 9 View FIGURE 9 , A3-e).
The chromosomal set of C. yaldeni sp. nov. (2n = 36, NFa = 52) comprises 4 pairs of metacentric, 5 pairs of subtelocentric and 8 pairs of acrocentric autosomes. The X-chromosome is large metacentric; the Y-chromosome is small subtelocentric.
Variation. Specimens of the type series are uniformly colored. In external and skull measurements S-165340 (female) is slightly smaller (HB = 84.0 mm, vs. 94.0– 99.5 mm in males). The qualitative characters are relatively homogeneous.
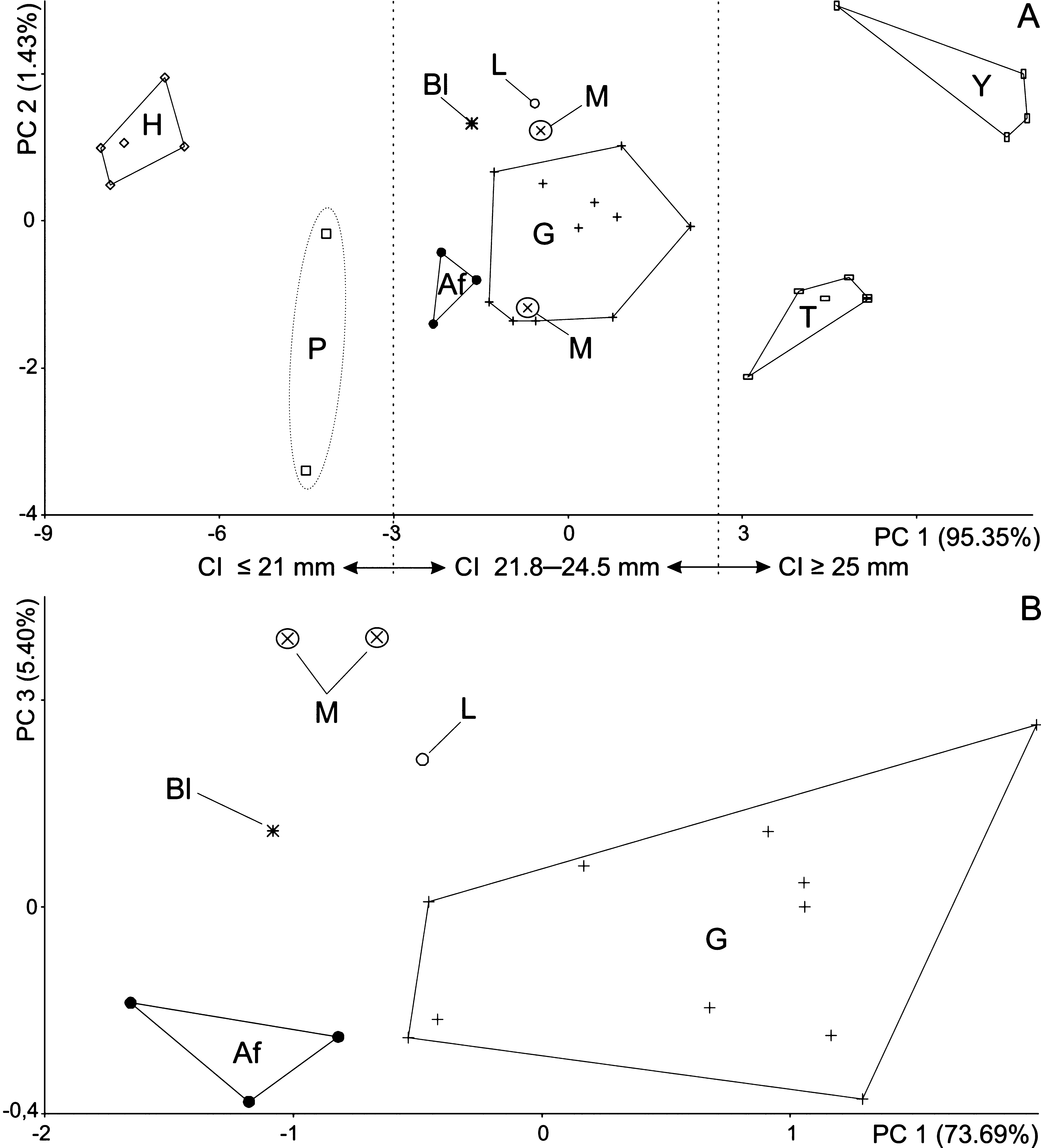
Comparisons. This is a large-sized shrew, comparable in external size (HB, TL) to C. thalia , but slightly larger in cranial size ( Table 2 View TABLE 2 ). It is substantially larger than the following Ethiopian Afromontane–Afroalpine species: C. bottegoides , C. cf. bottegi , C. cf. hildegardeae , C. harenna , C. phaeura , C. parvipes , C. afeworkbekelei sp. nov., C. lucina , C. macmillani , C. baileyi , and C. glassi . The values of CI for all mentioned species range between 14.3–24.51 mm. The new species is considerably smaller than the other Ethiopian Crocidura species found in the Afromontane–Afroalpine biozones: C. zaphiri (HB = 105 mm, CI unknown; Churchfield & Jenkins, 2013a), and C. olivieri (CI = 32.0– 34.3 mm; Churchfield & Hutterer, 2013).
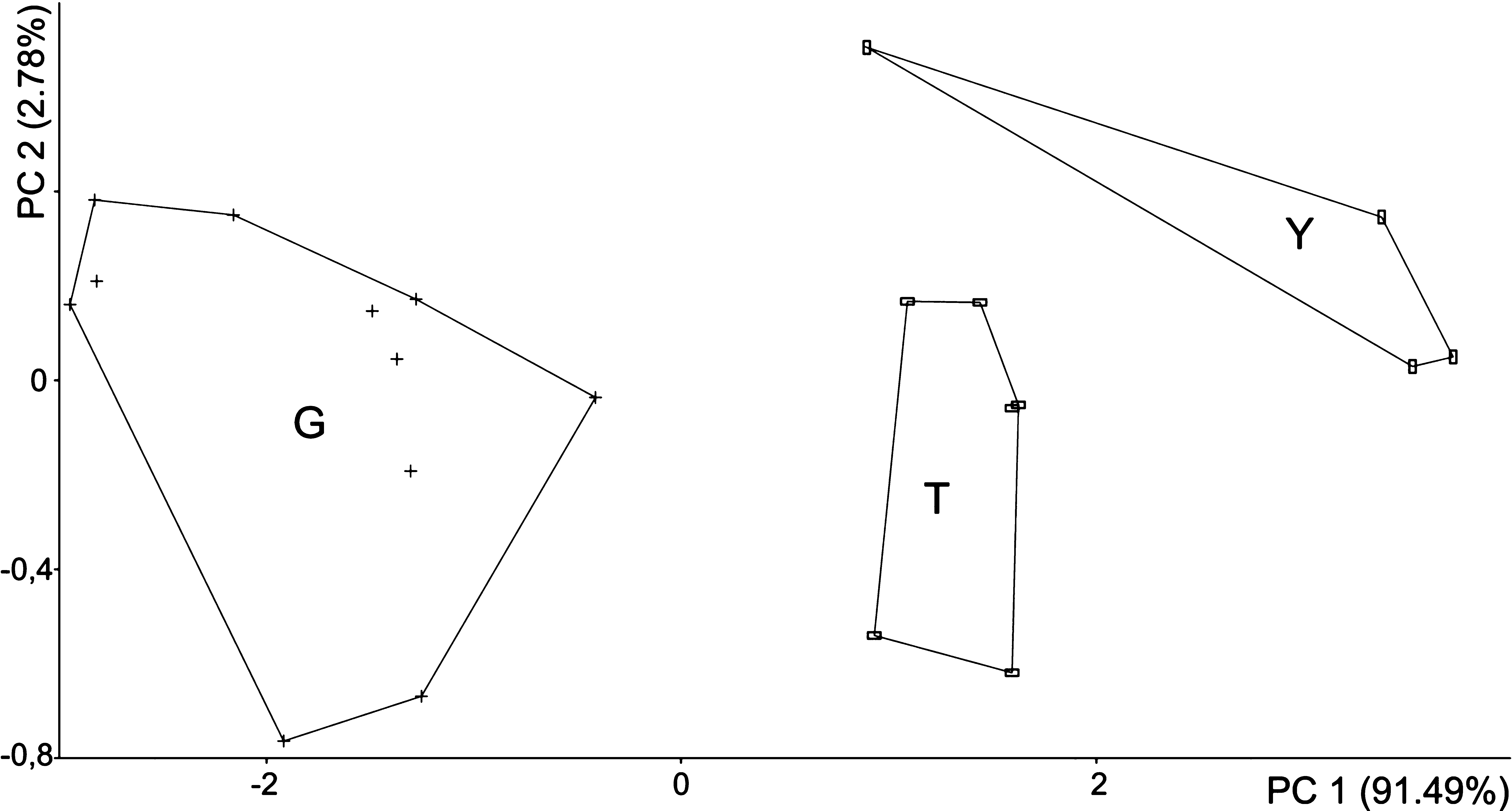
The first PCA (see above, and Fig. 7 View FIGURE 7 A) revealed significant differences between the groups of smaller, medium-sized and larger shrews along significant PC 1. Therefore, a further PCA was performed on two larger species and C. glassi .
The third PCA ( Fig. 10 View FIGURE 10 ) of all 25 cranial and mandibular linear measurements reveals that the smaller C. glassi is clearly separated from the larger C. thalia and C. yaldeni sp. nov. along PC 1. This statistically significant axis accounted for 91.49% of the total variance and is most correlated with measures of the general size of the skull and mandible, such as CI (r = 0.674), PL (r = 0.316), MBL (r = 0.297) and COR (r = 2.788). The second component accounted only for a small percentage of the total variance (2.78%), and therefore was not significant. The PC 2 is most positively correlated with cranial width (PGW, r = 0.576; ZYG, r = 0.349; GW, r = 0.261). As can be seen from Fig. 10 View FIGURE 10 , C. thalia and C. yaldeni sp. nov. are clearly distinguished along the first two principal components. The difference between the two species along the first axis reflects the larger average size of C. yaldeni sp. nov., while the second component is associated with changes in skull shape.
Detailed comparison between similar in size species: C. yaldeni sp. nov. differs from C. thalia in: smaller P4s/ d, M3s/d; bigger M2L, M3W, M3L, UML, ZYG, PGW, LML; longer hindfoot and claws ( Table 2 View TABLE 2 ); the uniformly colored tail (tail of C. thalia is mottled with brown and pale ochre); the bicolored and longer bristle hairs of tail; more massive I1; more compact upper antemolars row; stronger developed incision on lingual edge of the M2 ( Fig. 9 View FIGURE 9 , A1-a vs. B1); the expression of hypoconulids on the lower molars ( Fig. 9 View FIGURE 9 , A2-d).
The standard karyotype is identical to that of two related species, C. thalia and C. glassi ( Lavrenchenko et al., 1997) . Phylogenetic analyses based upon repetitive DNA elements (taxonomic DNA fingerprint) and inter-SINE- PCR (IS-PCR) revealed that genetic distances between C. yaldeni sp. nov. and its two closest relatives, C. thalia and C. glassi , fell within the range usually recorded for interspecific genetic differentiation within Crocidura ( Bannikova et al., 2001; Bannikova et al., 2005). The former analysis strongly supported that C. yaldeni sp. nov. is a sister species to C. thalia ; in the latter analysis C. thalia and C. glassi had a tendency to form a clade against C.
yaldeni sp. nov. The phylogenetic analysis of an extended set of Crocidura species using mitochondrial cytochrome b gene sequences ( Lavrenchenko et al., 2009) revealed that C. yaldeni sp. nov. and C. macmillani form the most basal branch of the group of Ethiopian endemics (including C. glassi , C. thalia , C. lucina and C. baileyi ), whereas C. thalia appears as sister to C. glassi .
Distribution. The new species has been found only in the Beletta Forest (07º34'N, 36º31'E, 1900 m a.s.l.). We failed to trap C. yaldeni sp. nov. in any other site of this forest and other humid Afromontane forest blocks of SW Ethiopia: the Sheko Forest (07°04'N, 35°30'E, 1930 m a.s.l.), the Dushi Area of the Godare Forest (07°21'N, 35°13'E, 1200 m a.s.l.), the Meti Area of the Godare Forest (07°17'N, 35°16'E, 1370 m a.s.l.) and the Inegawa Forest (07°25'N, 35°24'E, 2340 m a.s.l.). Therefore, the currently known distribution range of this new species is extremely small.
Habitat. All four specimens of C. yaldeni sp. nov. were captured in the riverine variant of humid Afromontane forest on the bank of the small river (trees: Schefflera abyssinica , Croton macrostacis , Allophylus abyssinicus , Aningeria altissima, Malacanta alnifolia, Phoenix reclinata , Brucea antiderinterica , Polyscias fulva ; small trees: Vepris dainelii , Teclea nobilis , Dracaena afromontana , D. fragrans ; shrubs: Coffea arabica , Canthium oligocarpum , Galiniera coffeoides ; climbers: Embelia schimperi , Phychotria neglecta ; ferns: Pteris dentatum , Asplenium sandersoni , Pleopeltis sp.; herbs: Afromomum sp.). Three rodent species, Lophuromys chrysopus Osgood , Mus mahomet Rhoads , and Stenocephalemys albipes (Rueppell) were also collected at the same trapping site. All individuals of the new shrew were caught in Sherman live traps placed on the ground no more than 1 m from the river bank. Probably the very restricted range of C. yaldeni sp. nov. is associated with yet unknown habitat requirements, presumably more specific than 'river edge'. However, no morphological adaptations to some specific life style (including semi-aquatic adaptations) were observed.
Etymology. The new species is named in honor of the late Dr. Derek W. Yalden (1940-2013), who has contributed greatly to our knowledge on Ethiopian small mammals ( Fig. 11 View FIGURE 11 ). As the vernacular name for the new species we propose Beletta Shrew.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.