
Limnasterias estradivariae sp., 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4712.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:97EA65A8-C055-4B4A-A07C-4AC09323FB4C |
|
persistent identifier |
https://treatment.plazi.org/id/02EA2880-8BA7-4C06-99A2-782E11AAC69C |
|
taxon LSID |
lsid:zoobank.org:act:02EA2880-8BA7-4C06-99A2-782E11AAC69C |
|
treatment provided by |
Plazi |
|
scientific name |
Limnasterias estradivariae sp. |
| status |
sp. |
Limnasterias estradivariae sp. View in CoL nov.
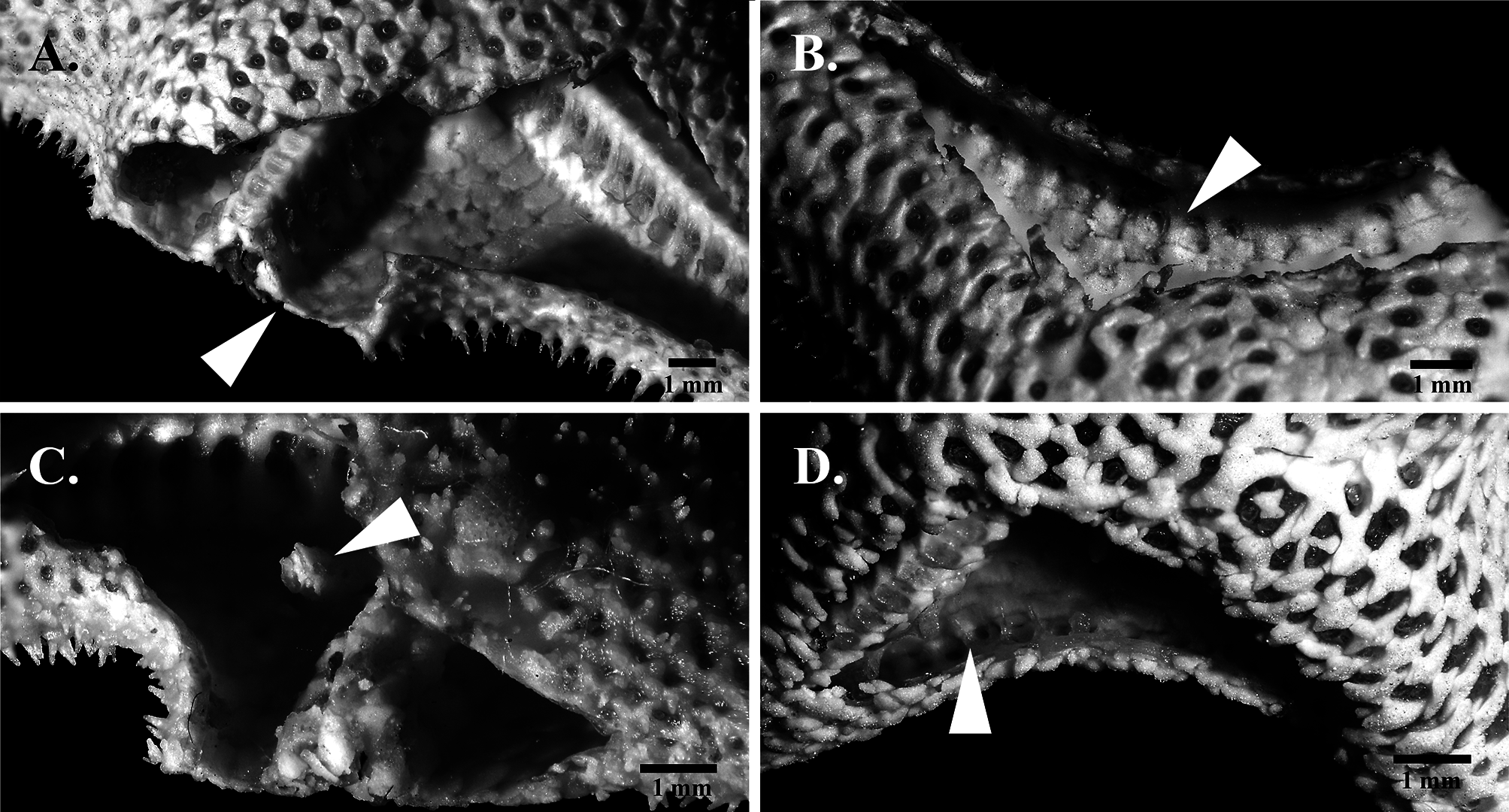
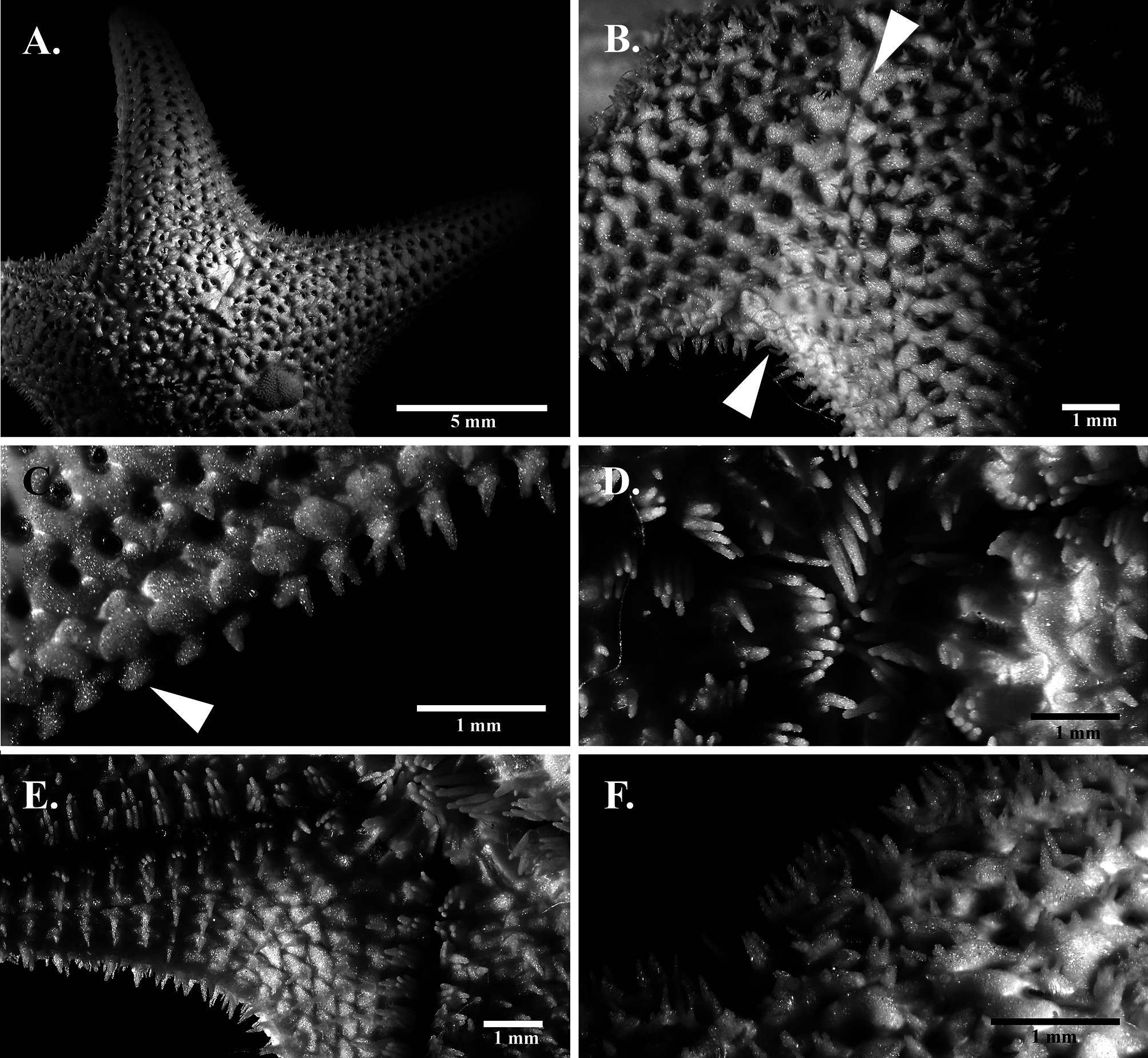
( Fig. 2C, D View FIGURE 2 ; Fig. 3 View FIGURE 3 ; Fig. 4C, E View FIGURE 4 )
Type Locality. Tanah Bamban Lake , Maratua Island, Derawan Archipelago, 1 m during low tide, on sand and macroalgae .
Material examined. Holotype. MZB. Astr. 00046, D. Kusuma, 20 February 2019, in ethanol; 3 Paratypes, MZB. Astr. 00047 (Type locality), A. Setiadi, 30 October 2018, in ethanol; 5 Paratypes, MZB. Astr. 00048 (Type locality), D. Kusuma, 20 February 2019, in ethanol.
Diagnosis. Non-fissiparous, small asterinid with R up to 12.35 mm; form five-rayed stellate, rays medium, basally rather broad with acute tips, R:r 2.0–2.6; body and rays high, semicircular in cross-section; abactinal plates with rounded keels, obscured by integument; papulae large, numerous, predominantly single; abactinal spinelets robust, digitiform to thick conical; up to 7 spinelets per plate, arranged linearly on plate apex; superomarginal plates circular, armed with up to two spinelets proximally; inferomarginal plates armed with clusters of digitiform to conical spinelets, up to 5; lengths of two or three inferomarginal spinelets often equal or subequal; oral spines digitiform, up to 5; furrow and subambulacral spines digitiform, subambulacral spines not taller than furrow spines. Color in life dark brown to dark gray abactinally, may photograph bluish gray; papulae translucent white, with pale spinelets; brown to beige actinally.
Description. Five rayed, non-fissiparous, small asterinid, R up to 12.35 mm, R:r 2.0–2.6. Ray rather broad at base, tapering acutely, with semicircular profile. Disc variably bordered by plates, with pentagonal outline to irregular or incomplete border. Single madreporite. Thick integument covers abactinal surface. Imbricating abactinal plates triangular to crescentiform, rather thick with rounded keel; plates irregularly aligned at ray median and proximal region, adjoined dorsolaterally by regular series of crescentiform plates. Extensive papular area with up to five rows of papulae at ray base, along with irregular papular array at the top of rays; papulae large, predominantly single, occasionally two per papular space; plates with papulae often separated from the superomarginals by zero to two non-papulate plates. Secondary plates rather rare, present on proximal region. Interradial non-papulate area present as opposing triangular fields, near disc borders and at the interradial margins ( Fig. 3B View FIGURE 3 ).
Robust abactinal spinelets are digitiform to thick conical, up to seven spinelets per plate, primarily 4–6 spinelets near disc ( Fig. 3F View FIGURE 3 ), 2–3 towards distal region or near margins; spinelets arranged linearly at plate apex. Glassy convexities present on plates. Pedicellariae absent. Superomarginal plates circular, armed with up to 2 spinelets proximally. Boot-shaped inferomarginal plates with single distal protrusions ( Fig. 3C View FIGURE 3 ), with clusters of up to five large, robust, conical spinelets; two or three inferomarginal spinelets are frequently equal or subequal in size, although clusters with one significantly larger spinelets are sometimes present ( Fig. 3B, C View FIGURE 3 ).
Actinal plates aligned obliquely, with up to two blunt conical spines per plate, predominantly one. Spines are sheathed with translucent tissue in live or intact animals. Digitiform furrow spines up to six, frequently three to four; subambulacral spines digitiform, up to two spines proximally. Adradial plates with single conical, blunt spines, not dissimilar from nearby actinal plates. Oral plates with five long, blunt digitiform spines; suboral spines up to two, with the proximal spine larger than the distal ( Fig. 3D View FIGURE 3 ).
Superambulacral plates absent ( Fig. 2C View FIGURE 2 ) with reduced, single series of superactinal plates at the intersection of abactinal and actinal body walls ( Fig. 2D View FIGURE 2 ); internal surface lined with resinous coating, with thickened distal edges of actinal plates. Interradial pillars are present. Sites of gonad attachment difficult to discern due to large gonadal mass crowding the small interior, and gonopores are not evident.
Color in life dark brown to dark gray abactinally ( Fig. 4C View FIGURE 4 ), which may photograph bluish gray; madreporite and spinelets pale cream, with translucent white papulae; actinal surface beige to brown.
Distribution. Tanah Bamban Lake, on Maratua Island, at depths of 1m or deeper.
Habitat. On sediment and among thalli of fleshy macroalgae ( Fig. 4E View FIGURE 4 ). Limnasterias estradivariae appears to be largely restricted to depth of one meter or beyond.
In situ observations. The depth restriction of L. estradivariae sp. nov. contrasts with L. oinops sp. nov., the latter of which are often found in very shallow microhabitats. Unlike Haji Buang Lake, Tanah Bamban Lake has markedly scarce sponges and macroalgae at intertidal depths. The shallow areas are instead covered by turf algae, mussels and cidaroid urchins Phyllacanthus sp. The fleshy macroalgae with which L. estradivariae sp. nov. appeared to co-occur are also rare from intertidal depths.
Etymology. Named after colleague Estradivari, who brought Limnasterias oinops to author’s attention in 2010 and encouraged the author to begin its description in 2017.
Remarks. Preliminary data suggest that collected Limnasterias estradivariae individuals are similarly sized thus far (mean R ± SD = 10.7 mm ± 1.2 mm, n = 13 ind.). It is unclear if this reflects smaller cohorts, ontogenetic habitat segregation, size-selective predation or other factors. This contrasts with L. oinops , wherein large adults cooccur with smaller individuals in the same habitat (mean R ± SD = 16.5 mm ± 5.3 mm, n = 17 individuals). Other characters distinguishing L. oinops and L. estradivariae are listed in Table 2 View TABLE 2 .
Discussion. This description of asterinids from two marine lakes in Indonesia relies on morphological characters, and may be further supported by future molecular studies. Some shared characters suggest possible affinities with Disasterina Perrier, 1875 and Tegulaster Livingstone, 1933 , including stellate form, integument cover, relatively sparse conical to sacciform spinelets at the abactinal, monacanthid actinal plate and median band of irregular abactinal plates. Representative molecular analyses on Limnasterias , Disasterina and Tegulaster may also help clarify the phylogenetic positions of Tegulaster- like Disasterina species, including D. ceylanica Doderlein 1888 ( Saba et al. 2012). Presently, there are no molecular data available for any of these genera ( O’Loughlin & Waters 2004).
The Haji Buang and Tanah Bamban Lakes are marine lakes with salinity range of 26 to 28.5‰ and 26‰ respectively ( Becking 2012). Limnasterias is perhaps the only known asteroid genus endemic to perpetually brackish habitats. Tolerance towards periodic hyposalinity was noted for asterinid Patiriella regularis from New Zealand fjords, which tolerates deionized water (0‰) for up to four days ( Barker & Russell 2008). A few other asteroids including Asterias rubens and Luidia clathrata are at times found in fluctuating estuarine or hyposaline habitats with salinity range of 8–27‰ ( Stickle & Diehl 1987). The extent of hyposalinity tolerance of Limnasterias oinops sp. nov. and L. estradivariae sp. nov. is not yet known; however, preliminary observation implies that individuals of Limnasterias oinops sp. nov. are unharmed by temporary freshwater lens that forms in the upper layer after rain (pers. obs.).
The Haji Buang Lake is currently threatened by encroaching developments in tourism infrastructure. An airport is located 500 m away from the lake and a number of hotels and resorts are currently under construction. Household activities and structures such as washing, lavatories and concrete buildings are likely to pollute the lake through leaching, given the porosity of the lake’s karstic boundaries and their subsurface connections ( Dawson et al. 2009; Marrack et al. 2015). In comparison, Tanah Bamban Lake is less subject to anthropogenic pressures due to its distance from human habitations. It is plausible that increasing severity of present impacts may drive future declines in Limnasterias gen. nov. populations.
Although populations of both species currently appear to be abundant, the likely endemicity of L. oinops sp. nov. and L. estradivariae sp. nov. to their respective lakes further heightens their vulnerability to extinction due to their restricted distribution. The likely recent extinction of Derwent River sea star, Patiriella littoralis (Dartnall 1970) and increasing rarity of other geographically isolated asterinids e.g. Parvulastra parvivipara (Keough & Dartnall, 1978) due to habitat degradation, isolation and possible competition against introduced species ( Liversage & Byrne 2018; O’Hara et al. 2019) highlight the threats these new species might be facing. Further exploitation by trade is also a potential threat unless protection is implemented, as Indonesia plays a major role in the supply network of ornamental marine invertebrates ( Wabnitz et al. 2003). Thus, Limnasterias oinops sp. nov. and L. estradivariae sp. nov. will benefit from an ecological and population assessment, together with protection at regional and greater scales if possible.
Further studies on marine lake asteroids will elucidate their diversity and ecology in these little-explored ecosystems. Marine lakes provide unique conditions and insularity conducive for speciation ( Dawson 2016; Dawson & Hamner 2005). Future discoveries of new forms of lake asterinids are anticipated, considering the emergence of marine lakes after the last glacial maximum around 18,000 years ago ( Dawson et al. 2009), coupled by the ability of some asterinid taxa to colonize marginal habitats ( Barker & Russell 2008; Byrne & Walker 2007) and evolve more rapidly than the timespan ( Puritz et al. 2012).

| MZB |
Museum Zoologicum Bogoriense |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.