
Jungermanniales
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.9.1.13 |
|
DOI |
https://doi.org/10.5281/zenodo.5076293 |
|
persistent identifier |
https://treatment.plazi.org/id/03F587D3-FFD4-B945-B597-4CB4FD26FCA8 |
|
treatment provided by |
Felipe |
|
scientific name |
Jungermanniales |
| status |
|
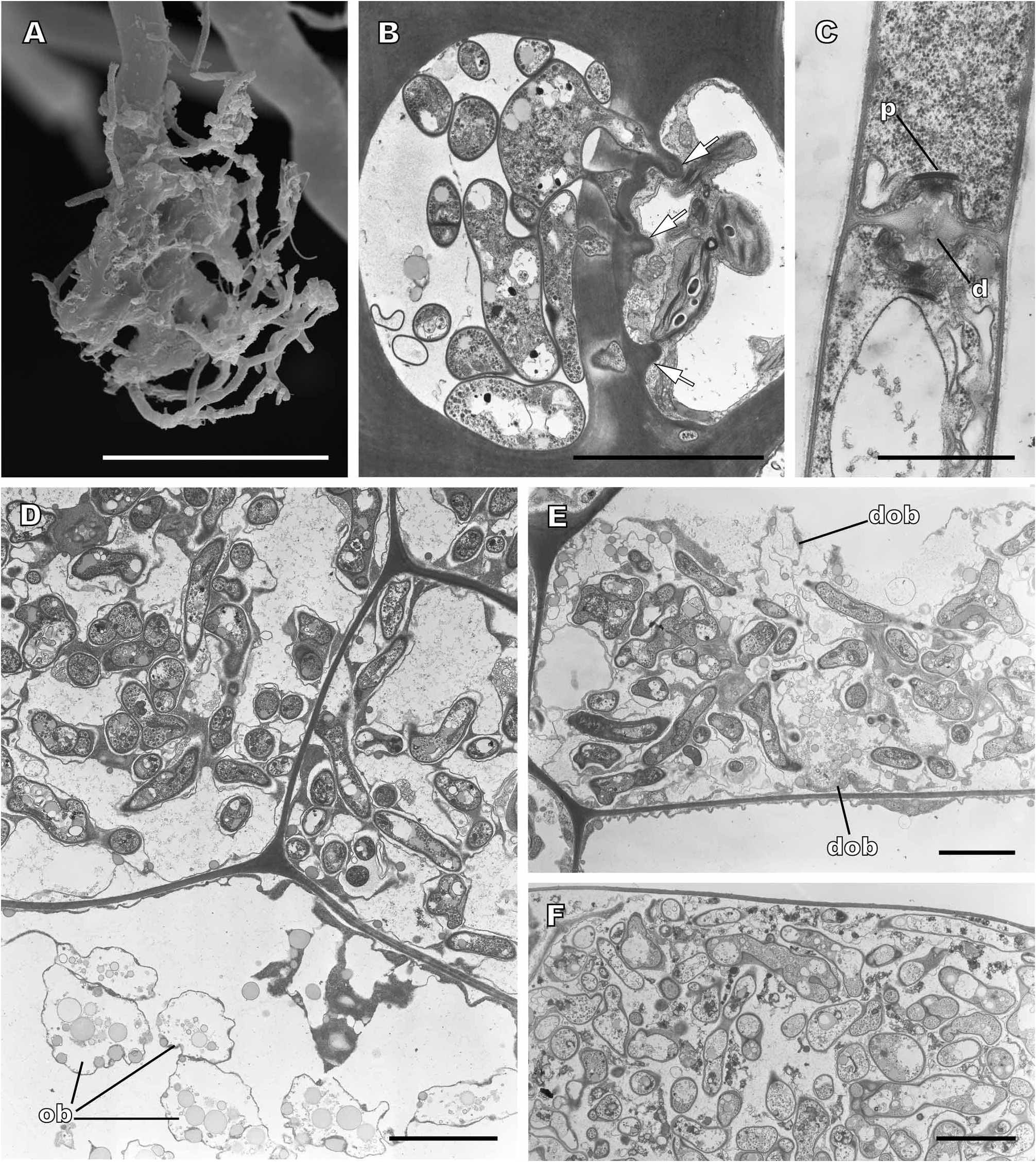
Electron microscopy and molecular studies ( Read et al. 2000; Kottke et al. 2003; Kottke & Nebel 2005; Nebel et al. 2004; Duckett et al. 2006b; Bidartondo & Duckett 2010; Newsham & Bridge 2010) have shown that eight or nine leafy liverwort genera; Barbilophozia Loeske , Diplophyllum (Dumort.) Dumort. , Lophozia (Dumort.) Dumort. , Scapania (Dumort.) Dumort. and Tritomaria Schiffn. ex Loeske in the Scapaniaceae , Nardia Gray in the Jungermanniaceae , Saccogyna Dumort. in the Geocalycaceae and Southbya Spruce , and almost certainly Gongylanthus Nees , in the Arnelliaceae , consistently and predominantly associate with members of the Sebacina vermifera Oberw. species complex, in striking contrast to Tulasnella as the predominant endophyte in the Aneuraceae . As in most liverwort-fungus associations, these endophytes enter the hosts via the rhizoids. However colonization by these basidiomycetes causes the rhizoids to become highly branched ( Fig. 5A View FIGURE 5 ) rather than eliciting the terminal swellings typical of ascomycetous infections. Within the liverwort stems two kinds of association have been described. In the Arnelliaceae the endophytes colonize a cluster of cells in the centre of the stems ( Fig. 5D View FIGURE 5 ) and there are no visible features in the host walls indicating why the fungus is so restricted. In the Scapaniaceae , Jungermanniaceae and Geocalycaceae , the central region of the stems contains a mosaic of infected and fungus-free cells. Growth of hyphae into the latter is prevented by overgrowths of host wall material ( Fig. 5B View FIGURE 5 ). In both kinds of infection there is no evidence of fungus digestion by the host and healthy hyphae persist long after the death of the host cells in older stems ( Fig. 5F View FIGURE 5 ). In fact, host cell death occurs very rapidly in the mosaic infections ( Duckett et al. 2006b). One previously overlooked effect of fungal colonization in the Arnelliaceae is that oil bodies in infected cells rapidly break down ( Fig. 5E View FIGURE 5 ) as hyphae proliferate suggesting that their contents may be a nutrient source for the endophytes.
In striking contrast to the vast host range of the rhizoidal ascomycetes, cross-infection experiments on the leafy liverwort basidiomycetes strongly suggested these to be highly host specific ( Duckett et al. 2006b). This situation has now been confirmed by a molecular study ( Bidartondo & Duckett 2010) revealing a high level of fungal specificity to the extent that where multiple liverworts co-occur they almost never share the same endophyte. This remarkable symbiotic conservatism differs fundamentally from the generalist pattern of mycorrhizas in seed plants that repeatedly evolved to form ectomycorrhizas simultaneously with a range of distantly related basidiomycete genera.
The phylogenetic position of these basidiomycete-associated leafy liverworts ( Davis 2004; Forrest et al. 2006; Heinrichs et al. 2005, 2007; Hentschel et al. 2006) and the fact that they are far more restricted than those with ascomycetes suggest that the basidiomycetes were a secondary acquisition following the loss of ascomycetes. However, the position of the Sebacinales in fungal phylogenies suggests that they may be considerably more ancient than the Tulasnella associations in the Aneuraceae ( Kottke et al. 2003; Kottke & Nebel 2005). The discovery that basidiomycetes are widespread in Scapania , Diplophyllum , Barbilophozia and Lophozia is congruent with their close evolutionary relationship ( Schill et al. 2004; Yatsentyuk et al. 2004; He-Nygrén et al. 2006) whilst the placement of basidiomycete-associated liverworts ( Lophozia sudetica (Nees & Huebener) Grolle , Barbilophozia barbata (Schmieder ex Schreb.) Loeske , B. lycopodioides (Wallr.) Loeske and B. hatchery (A.Evans) Loeske ) sister to other members of the Lophoziaceae ( de Roo et al. 2007) suggests ancestral presence of fungi in this family.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.