
Ebertichthys ettlingensis, Arratia, 2016
|
publication ID |
https://doi.org/ 10.5194/fr-19-31-2016 |
|
DOI |
https://doi.org/10.5281/zenodo.11045874 |
|
persistent identifier |
https://treatment.plazi.org/id/03F68791-BA22-2F16-FFFF-FB02FACCD0AC |
|
treatment provided by |
Felipe |
|
scientific name |
Ebertichthys ettlingensis |
| status |
sp. nov. |
Ebertichthys ettlingensis n. sp.
Figures 2–13 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 View Figure 9 View Figure 10 View Figure 11 View Figure 12 View Figure 13
2008 Undescribed small teleost. – Ebert and Kölbl-Ebert, fig. 6.
2011 Ascalabos -like. – Ebert and Kölbl-Ebert, fig. 8.
2015 Teleost sp. 1. – Ebert et al., p. 21, fig. 13b; p. 22, fig. 14a.
Diagnosis: Same as generic diagnosis.
Derivation of name: The specific name ettlingensis refers to the locality of Ettling and its rich and beautifully preserved fossils, from which the fishes were recovered.
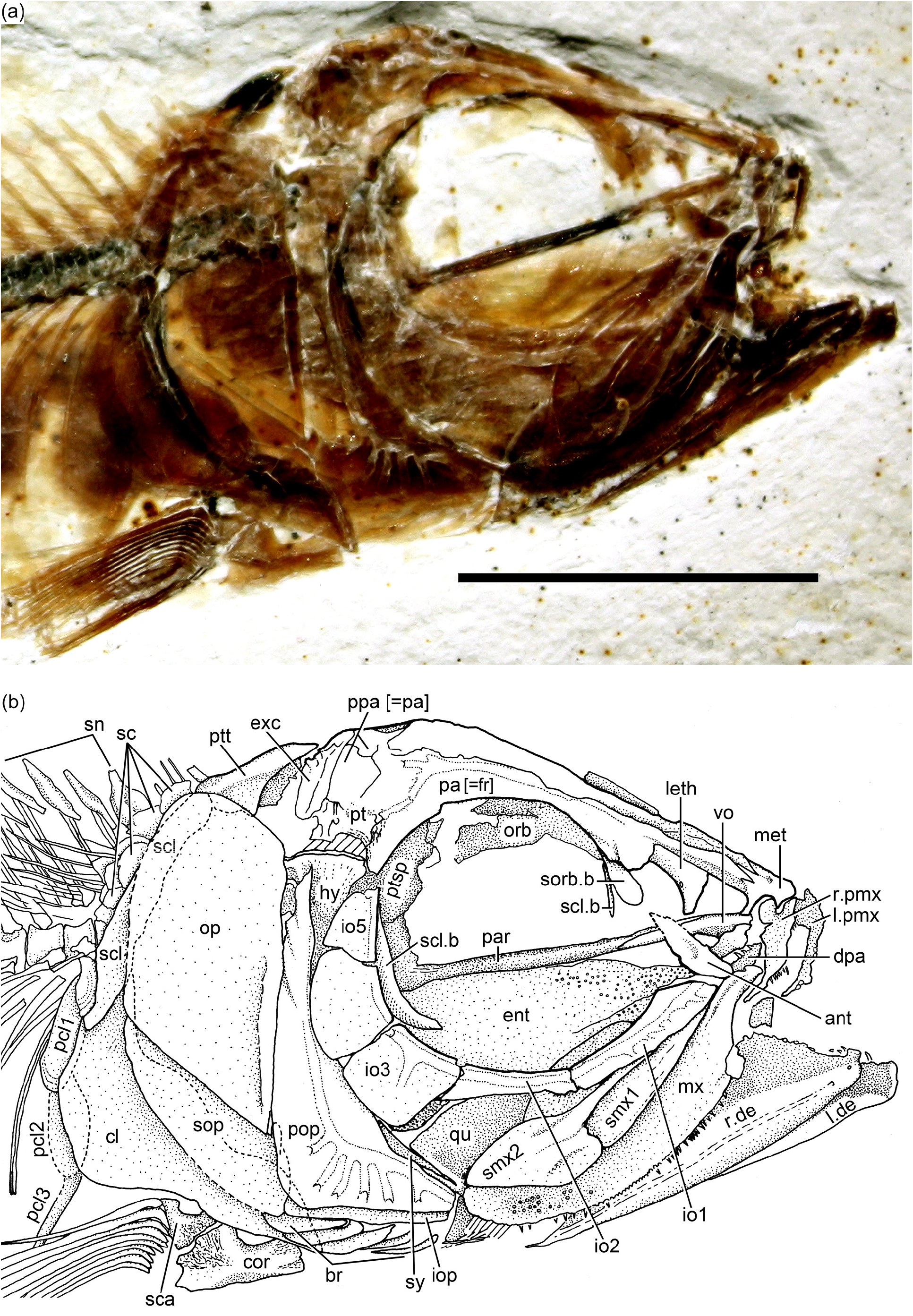
Holotype: JME ETT 108 a and 108b is preserved in part and counterpart. It is a complete, beautifully preserved specimen of about 75 mm total length including soft anatomy preservation (part of digestive system) and squamation ( Fig. 2a View Figure 2 ). The specimen is heavily ossified, so that it is interpreted as an adult although the presence of unfused hypurals 1 and 2 and haemal arch of preural centrum 1 with their respective centrum.
Paratypes: JME ETT 11 , JME ETT 24 ( Fig. 2c View Figure 2 ) , JME ETT 60 , JME ETT 61 , JME ETT 64 a, JME ETT 132 a, b ( Fig. 2b View Figure 2 ), JME ETT 148 , and JME ETT 847 a.
Type locality and age: Ettling (Plattenkalks I– III), Bavaria ( Arratia and Tischlinger, 2010; Ebert et al., 2015). Upper Jurassic, probably upper Kimmeridgian ( Ebert et al., 2015) to lower Tithonian. However, the age of the locality is approximated due to the fact that well-preserved ammonoids have not been recovered.
Description
General description: The fish is small, below 90 mm total length, slightly fusiform, and with dorsal, pelvic, and anal fins producing a triangular outline, with the dorsal fin almost equidistant to the pelvic and anal fins ( Fig. 2a–c View Figure 2 ). Dorsal fin insertion placed posterior to the midpoint of standard length (61 to 64 %). Pelvic fin insertion about at the midpoint of standard length (about 52 to 55 %). Anal fin insertion 69 to 73 % of standard length (SL). Caudal peduncle deep, about half of the maximum body depth. The head is proportionally large, about 33 % of standard length. In a young individual (JME ETT 61) of 34.7 mm total length, the head is comparatively larger (37 % of SL). Eye large, 34 to 36 % of head length.
Skull roof and braincase: The skull roof is incompletely preserved or partially damaged in the studied specimens. Additionally, the skull is laterally compressed from burial, so that it is difficult to describe particular elements. All bones of the skull roof have smooth surfaces and are unornamented.
The main element of the skull roof ( Fig. 3a–b View Figure 3 ) is the parietal bone (i.e., frontal bone of traditional terminology) that occupies most of the preorbital region and part of the postorbital region, which is short. The postorbital region is about one-third of the preorbital length. Anteriorly, the parietal bones suture with a broad and short mesethmoid ( Figs. 3b View Figure 3 and 4b View Figure 4 ). The latter bone presents two short and narrow lateral processes. Posteriorly, the mesethmoid sutures with the parietals throughout several interdigitating projections of different lengths. The parietals apparently leave a long, narrow space between them as a cranial fenestra, a condition that also has been observed in Ascalabos voithii ( Arratia, 1997) . A nasal bone is laterally placed to the anterior part of the parietal bone. The bone seems to be narrow and elongate, mainly carrying the anterior section of the supraorbital canal. Due to poor preservation, the limits between the parietal, postparietal, pterotic, and autosphenotic are not discernable, but the postparietal bones seem to be small and sutured to each other medially. The pterotic is very short and is the main element that articulates with the hyomandibula. The pterotic sutures with a small, triangular, and well-ossified autosphenotic anteriorly and with the parietal and postparietal medially. Its posterior region is covered by an elongate, narrow extrascapula. A short section of the middle pit line is observed, but it is unclear whether the pit line crossed the pterotic or not. The supraoccipital seems to be very small, with a very low crest. Boundaries between these bones ( Fig. 3a–b View Figure 3 ) are difficult to discern due to the fragility and transparency of the bones.
Anteriorly and below the parietal, the autosphenotic sutures with a well-developed and ossified chondral bone, the pterosphenoid ( Fig. 3b View Figure 3 ). Anterior to the pterosphenoid is another chondral bone that has a median position, the orbitosphenoid. The latter extends anteriorly and ventrally, producing an incomplete interorbital septum. The lateral ethmoid is well developed, but its preservation does not allow a proper description. A section of the parasphenoid is visible throughout the large orbit. It has a well-developed ascendent process suturing with the pterosphenoid. Anteriorly, the parasphenoid sutures with the well-developed posterior process of vomer ( Fig. 3b View Figure 3 ); however, the suture between both bones is unclear in other specimens ( Fig. 4a–b View Figure 4 ). There are numerous, minuscule teeth scattered below the parasphenoid, vomer, and entopterygoid ( Figs. 3b View Figure 3 and 4b View Figure 4 ). Because of the angle at which some of those teeth are preserved in JME ETT 132a, it is possible to assume that not only the entopterygoid carried teeth but also the vomer and parasphenoid. This assumption should be tested when more specimens become available. It is unclear whether a basipterygoid process was present or not.
The supraorbital canal is a simple canal with a few tubules. The short parietal branch ( Fig. 3b View Figure 3 ) is rudimentary and does not extend posteriorly into the postparietal bone. The supraorbital canal does not produce a lateral tubule near the autosphenotic; thus, a connection between supra- and infraorbital canals is absent. Due to condition of preservation, the otic canal was not observed in the studied material.
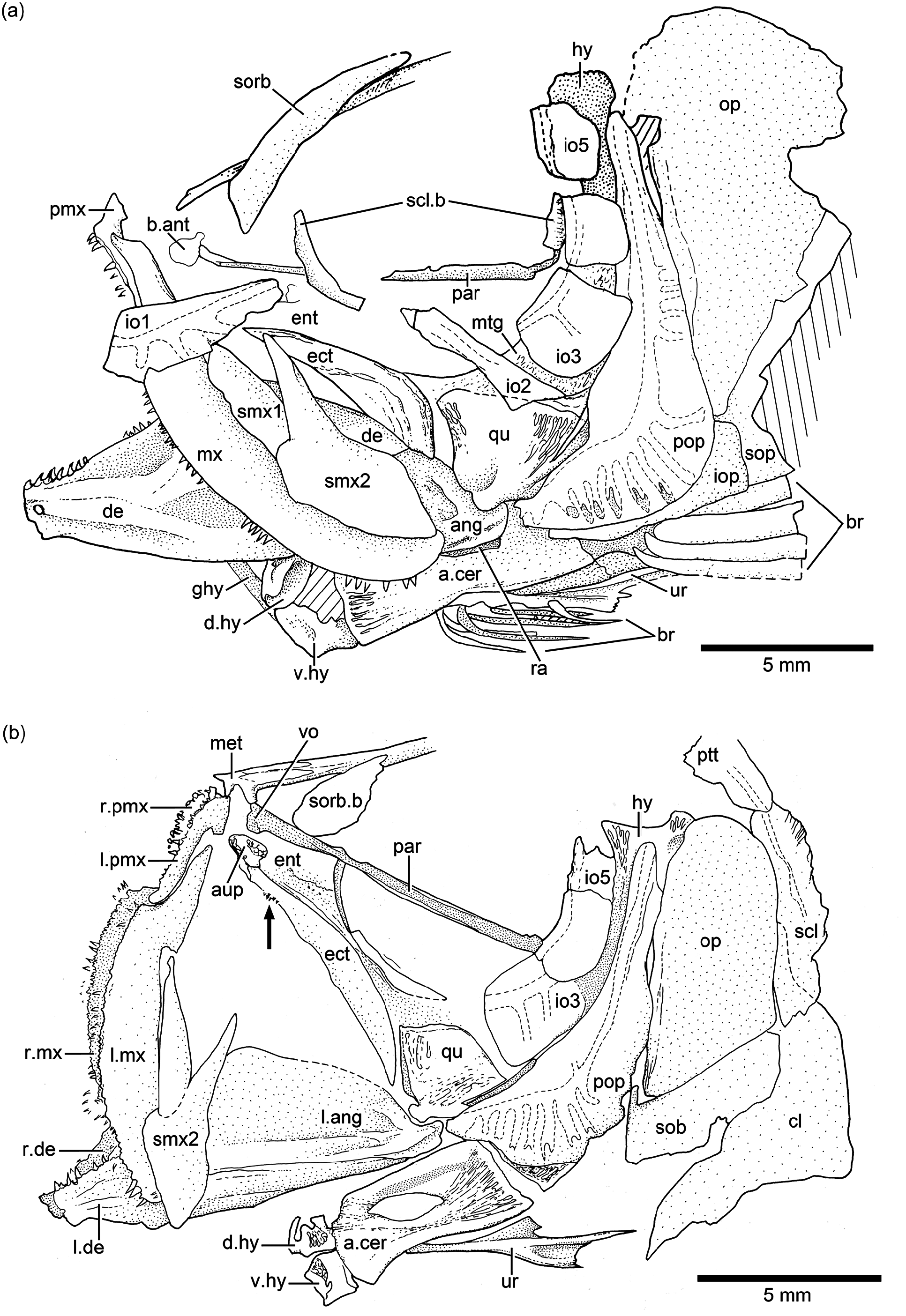
Orbit and circumorbital series: The fish has a large orbit ( Figs. 2a–c View Figure 2 and 3a–b View Figure 3 ), so that the postorbital distance between the margin of the orbit and the preopercle is very narrow. This space is partially covered by the narrow and small infraorbitals 4 and 5, which do not reach the preopercle posteriorly, so that the dorsal part of the hyomandibula is exposed laterally ( Figs. 3b View Figure 3 , 4a–b View Figure 4 , and 5 View Figure 5 ).
The series of circumorbital bones apparently encloses the large orbit completely, although a dermosphenotic is not preserved in the studied specimens. However, given the space between the most posterodorsal infraorbital and the autosphenotic, it is assumed that the bone was small. The series includes a supraorbital, antorbital, and five (occasionally six?) infraorbitals ( Figs. 3b View Figure 3 , 4a–b View Figure 4 , and 5 View Figure 5 ). Specimen JME ETT 148 seems to have six infraorbitals ( Fig. 5 View Figure 5 ); however, and by comparison with other specimens, I prefer to interpret this 6th bone as result of a fracture of infraorbital 5, and not as a dermosphenotic, because the infraorbital canal does not turn posteriorly to join the otic canal. There are two large sclerotic bones occupying anterior and posterior positions.
The supraorbital bone ( Fig. 4a–b View Figure 4 ) is large, well ossified, and expanded anteriorly. Only its anteriormost portion is preserved in the holotype ( Fig. 3b View Figure 3 ). Because of its size and it being displaced in all specimens, it is interpreted here as a bone that covers most of the dorsal margin of the orbit.
The antorbital ( Fig. 3b View Figure 3 ) is a slightly elongate, triangular bone that is comparatively larger than antorbitals present in other Jurassic teleosts. An antorbital branch of the infraorbital canal has not been observed.
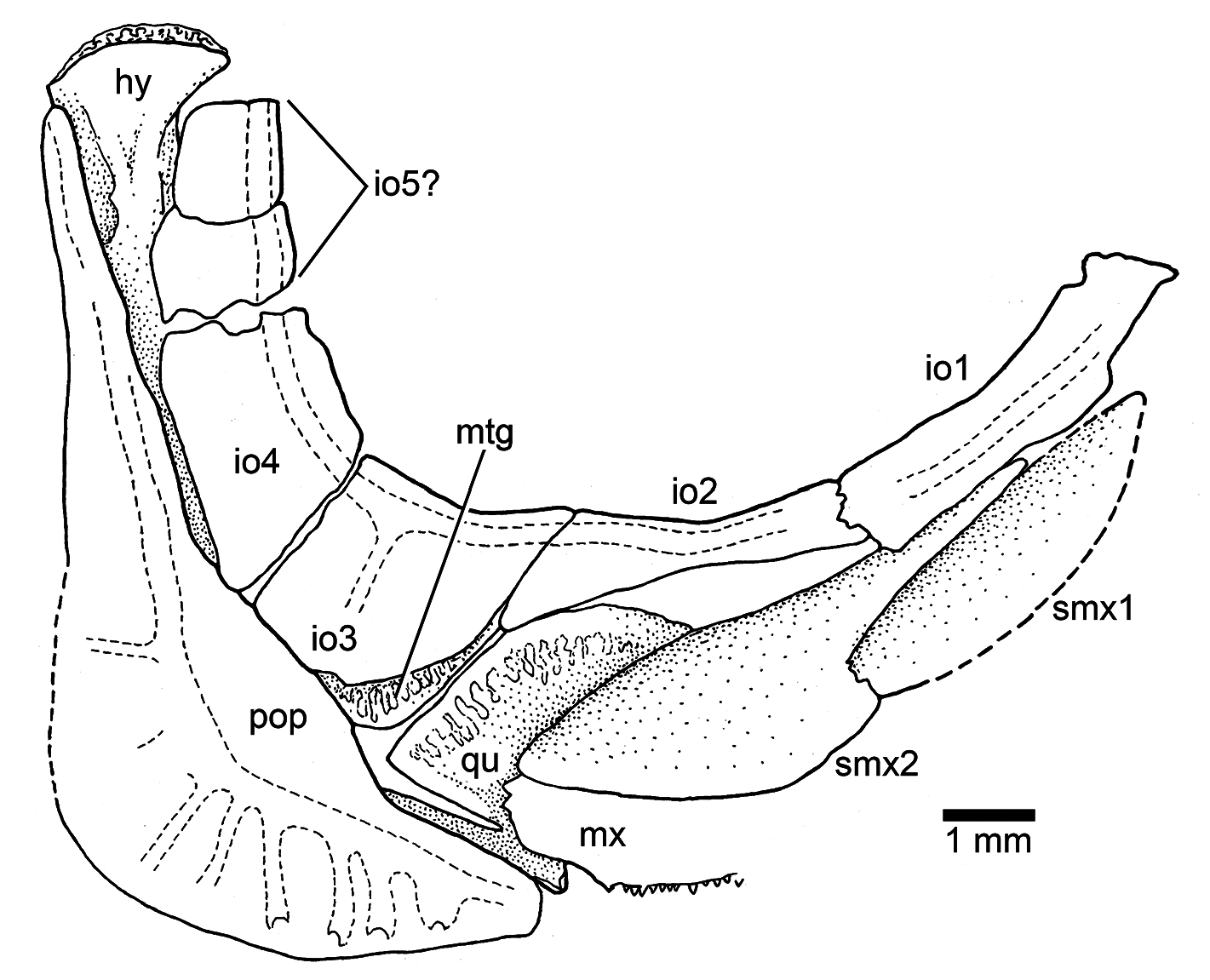
Infraorbital 1 ( Figs. 3b View Figure 3 , 4a View Figure 4 , and 5 View Figure 5 ) is an elongate, narrow bone that is slightly broader at its anterior margin than posteriorly. Infraorbital 2 ( Figs. 3b View Figure 3 , 4a View Figure 4 , and 5 View Figure 5 ) is long and narrow, but becomes broader at its posteroventral corner, joining infraorbital 3. The bone is so long that it is the main element forming the ventral margin of the orbit. Infraorbital 3 ( Figs. 3b View Figure 3 , 4a–b View Figure 4 , and 5 View Figure 5 ) is a small, slightly square bone at the posteroventral corner of the orbit. Its posterior margin just reaches the anterior margin of the preopercle or both bones are separated by a short distance. Due to the small size of infraorbital 3, the quadrate is partially exposed laterally. Infraorbitals 4 and 5 form the posterodorsal margin of the orbit.
The infraorbital canal ( Figs. 3b View Figure 3 , 4a–b View Figure 4 , and 5 View Figure 5 ) is enclosed by thin bone and is of simple type. The main canal is relatively broad, with a few branching sensory tubules. At least three tubules are counted in infraorbital 1. They open close to the ventral margin of the bone. No sensory tubules branch off in infraorbitals 2, 4, and 5. Infraorbital 3 has only one or two sensory tubules that end at the middle region of the bone. Upper jaw: Premaxilla, maxilla, and two supramaxillae form the upper jaw. The premaxilla ( Figs. 3b View Figure 3 and 4a–b View Figure 4 ) is a slightly triangular bone, with a short, broad ascendent process and an elongate oral margin, which is as long as the elongate anterior articulatory region of the maxilla. The oral margin bears small, conical teeth. It is unknown how many rows of teeth were present.
The maxilla ( Figs. 3b View Figure 3 and 4a–b View Figure 4 ) is elongate, partially covering the lateral aspect of the quadrate and extending close to the posterior margin of the orbit. Its articulatory anterior region is about a third of the maxillary length. The ventral margin is slightly convex, as is its posterior margin. A row of small, conical teeth that increase in size posteriorly is present. The teeth are comparatively larger in younger states than in older (larger) specimens. The posterolateral side of the maxilla – which is well preserved in the holotype – presents small, rounded tubercles ( Fig. 3b View Figure 3 ), an uncommon feature in Late Jurassic teleosts, which lack ornamentation.
Two supramaxillae ( Figs. 3b View Figure 3 , 4a–b View Figure 4 , and 5 View Figure 5 ) lie on the dorsal margin of the maxilla. Both bones together occupy the whole length of the maxillary blade. Supramaxilla 2 has a broad, expanded body and a narrow, long anterodorsal process that almost covers the whole dorsal margin of supramaxilla 1, which is slightly ovoid.
Lower jaw: The jaw ( Figs. 3b View Figure 3 and 4a–b View Figure 4 ) is moderately long, with its articulatory region for the quadrate placed at the level of the posterior half of the orbit. The jaw is formed laterally by two bones, the dentary and the angular. The suture between both bones is not visible because of preservation, although discontinuous sections are observed in some specimens. From a narrow mandibular symphysis, the dentary expands posteriorly, producing together with the angular a massive and high coronoid process. A “leptolepid” notch has not been observed, because the maxilla covers the region where the notch would be placed. A retroarticular has not been observed in any specimen, so that it is assumed that the angular, articular, and retroarticular are fused in the medial side of the jaw. The postarticular process is elongate and well ossified. A narrow section of a chondral bone is medial to the ventral margin of the angular in specimens JME ETT 60 and is interpreted here as part of a branchial arch.
A surangular is not present at the posterodorsal corner of the jaw. Coronoid bones are not present either.
The mandibular canal is positioned near the ventral margin of the jaw. Pores have not been observed in the posteroventral region of the dentary and angular, so that it is assumed here that the mandibular canal exits medially.
Palatoquadrate, suspensorium, hyoid arch, glossohyal, and urohyal: Most of these bones are partially hidden by other bones, so that a complete description is restricted to a few elements. A small, somewhat square autopalatine ( Fig. 4b View Figure 4 ) is placed lateral to the vomer, and slightly dorsal to a small dermopalatine. The latter bears very small, conical teeth. The autopalatine and quadrate are separated by a thin, elongate entopterygoid ( Figs. 3b View Figure 3 and 4b View Figure 4 ) that is covered medially with tiny conical teeth, at least at the anterior part of the entopterygoid and below the orbit. The ectopterygoid ( Fig. 4b View Figure 4 ) is lateral to the entopterygoid and in front of the quadrate. The bone is elongate, boomerang-like, and has a small patch of tiny, conical teeth close to the palatine region. The quadrate ( Figs. 3b View Figure 3 , 4b View Figure 4 , and 5 View Figure 5 ) is slightly triangular, with its dorsal margin straight or slightly rounded, and a comparatively small, slightly rounded condyle for articulation with the lower jaw. Its posterodorsal margin sutures with the metapterygoid. The length of the posterodorsal process of the quadrate is unknown because the process is covered by the anterior margin of the preopercle. A small portion of the metapterygoid ( Figs. 3b View Figure 3 , 4a View Figure 4 , and 5 View Figure 5 ) can be observed below the small infraorbital 3. Only a portion of the upper region of the hyomandibula ( Figs. 3b View Figure 3 and 4b View Figure 4 ) and a portion of the ventral region of the symplectic are observed, because those bones are covered laterally by the posterodorsal infraorbitals and the preopercle. The dorsal portion of the hyomandibula ( Figs. 3b View Figure 3 , 4a–b View Figure 4 , and 5 View Figure 5 ) is narrow and apparently has one elongate articular region with the braincase.
The lower part of the hyoid arch has preserved the hypohyals, anterior ceratohyal, and a small portion of the posterior ceratohyal. The posterior ceratohyal is partially observed below the preopercle in specimen JME ETT 24. The dorsal and ventral hypohyals ( Fig. 4a–b View Figure 4 ) are slightly rectangular, and the ventral one is slightly larger than the dorsal one. The anterior ceratohyal ( Fig. 4a–b View Figure 4 ) is rectangular, with an ovoid “beryciform” foramen in its middle region, below the groove for the hyoidean artery. The anterior and posterior borders of the anterior ceratohyal are almost straight with no articular interdigitations.
The glossohyal ( Fig. 4a View Figure 4 ) is a small and elongate narrow bone extending anterodorsally to the dorsal hypohyals. An elongate, triangular urohyal ( Fig. 4a–b View Figure 4 ) is preserved in specimen JME ETT 24. The bone is narrow anteriorly and expands posteriorly producing three processes, the longest being the middle one.
Opercular and branchiostegal series and gular plate: The preopercle ( Figs. 3b View Figure 3 , 4a–b View Figure 4 , and 5 View Figure 5 ) is triangular-shaped, with its posterior and ventral margins forming an angle of about 90 ◦. The bone is not expanded posteroventrally, but it has a characteristically slightly rounded flange just anterior to the curvature of the preopercular canal. Its dorsal arm is long, almost reaching the posterolateral margin of the pterotic, whereas the ventral arm is short. All margins are smooth.
Due to conditions of preservation, the preopercular canal and its tubules ( Figs. 3b View Figure 3 , 4b View Figure 4 , and 5 View Figure 5 ) are incomplete in most specimens, with the exception of JME ETT 60 with sensory tubules preserved in both the ventral and dorsal regions ( Fig. 4a View Figure 4 ). Apparently, the preopercular canal gives off seven or eight tubules in the ventral arm and at least five or six tubules in the dorsal arm. The tubules are very delicate, simple, and narrow, and they open close to the margins of the bone. They are difficult to observe due to the transparency of the bone, and this may explain the differences observed among specimens illustrated herein.
The opercle ( Figs. 3b View Figure 3 and 4a–b View Figure 4 ) is large and dorsally is broadly separated from the skull roof bones and the braincase. The dorsal margin is gently rounded, whereas the anterior and posterior margins are almost straight, and the ventral margin is markedly oblique. The surface of the bone is smooth with its anterior margin thickened and heavily ossified.
The subopercle ( Figs. 3b View Figure 3 and 4a–b View Figure 4 ) is a moderately narrow bone. Its depth is 4 times less than in the opercle. Its ventral margin is gently curved, and its anterodorsal process is short and rounded. The interopercle ( Figs. 3b View Figure 3 and 4a View Figure 4 ) is covered by the preopercle (or is not preserved) in most specimens, so that a description cannot be presented. Apparently, the interopercle is as long as the ventral margin of the preopercle as shown by the holotype.
There are 10 incompletely preserved branchiostegal rays ( Fig. 4a View Figure 4 ) in specimen JME ETT 60 ; six narrow and short branchiostegals articulate with the anterior ceratohyal, and four broader ones articulate with the posterior ceratohyal. Eleven rays are counted in the holotype, the first ones narrow and partially destroyed. Nevertheless, it is possible that the number was much higher than 11. A gular plate has not been observed .
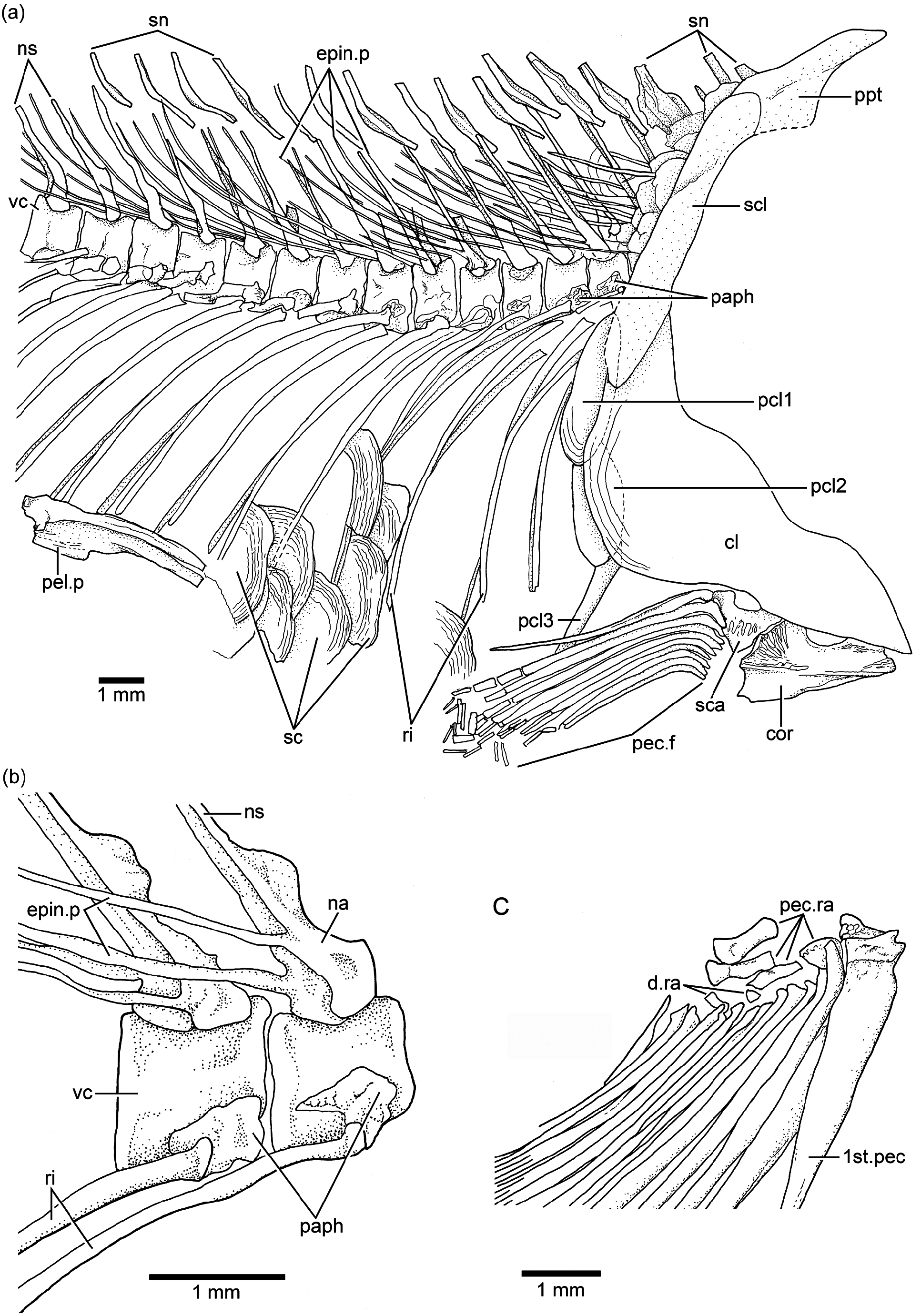
Vertebral column and intermuscular bones: There are 38 or 39 vertebrae, including preural centrum 1; from these, 21 or 22 are abdominal or precaudals, so that the caudal region is shorter than the abdominal one. The first four or five vertebrae are covered laterally by the opercle. All vertebrae are well ossified and with smooth lateral surfaces ( Figs. 2a–c View Figure 2 , 6a–b View Figure 6 , 7 View Figure 7 , and 8). The centra are almost square, as deep as long, with the exception of the preural centra that are slightly deeper than long and of the ural centra, which are reduced in size. All centra lack pre- and postzygapophyses.
The neural arches of the abdominal vertebrae are autogenous, and the halves of each arch are unfused medially. The neural arches are comparatively narrow and sit on the middle of the dorsal region of each centrum. Each neural arch ( Figs. 3b View Figure 3 and 6a–b View Figure 6 ) is slightly expanded proximally, and it has a slightly expanded anterior flange that gives the arch a bilobate aspect. Each epineural process ( Fig. 6b View Figure 6 ) emerges at the posterolateral margin of the arch. Most of the neural spines of the abdominal region are moderately inclined toward the horizontal, and they are moderately long, just reaching the ventral tips of the supraneurals. The spines are shorter than the epineural processes. The parapophyses ( Figs. 3b View Figure 3 and 6a–b View Figure 6 ) are fused to the anterolateral portion of the centrum, near its ventral margin. Each is formed by a thin chondral ossification with a prominent bony edge surrounding a small articulatory cavity placed ventroposteriorly in the parapophysis. The head of each rib articulates with the small articular cavity.
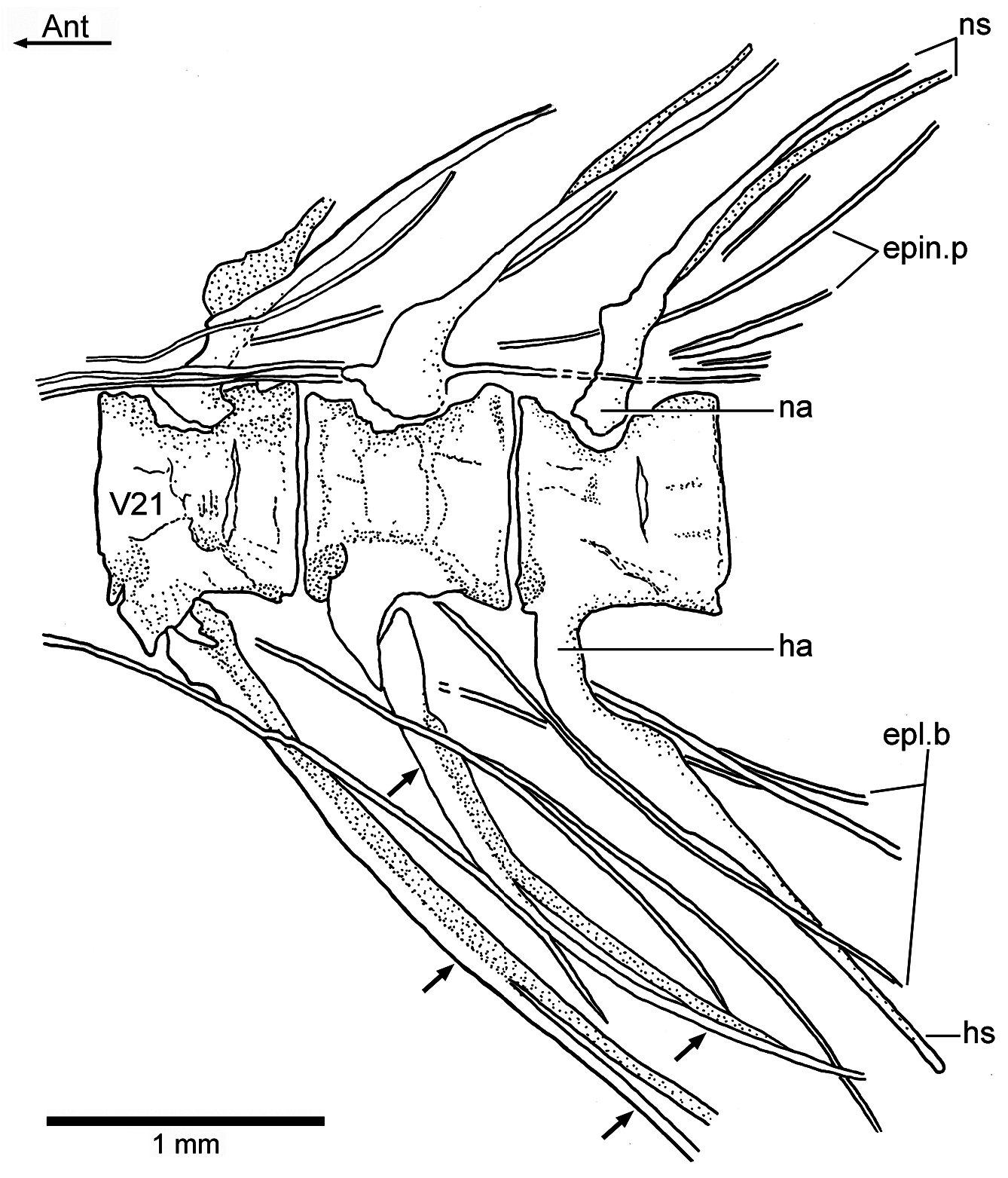
The neural arches of the caudal vertebrae ( Figs. 2a–c View Figure 2 , 7 View Figure 7 , and 8) are fused to their centra, with the exception of the first two or three caudal vertebrae (vertebrae 23 and 24) that have autogenous neural arches. All haemal arches are fused to their respective centra. The neural and haemal spines are narrow and end in an acute tip, with the exception of those of the preural centra. The neural and haemal spines are moderately inclined toward the body axis, with slightly greater inclination caudally. The haemal spines ( Fig. 7 View Figure 7 ) are short, not extending between the anal pterygiophores, except for the first two. The neural and haemal spines of the midcaudal region are completely ossified (membrane bone), lacking an internal core of cartilage; however, the condition changes in the preural region (chondral bone; see below).
There are 18 or 19 unquestionable pairs of ribs. The first two articulate with centra that are laterally covered by the opercle, and the last pair is positioned lateral to the first anal pterygiophore. All ribs ( Figs. 2a–c View Figure 2 , 3b View Figure 3 , 6a–b View Figure 6 , and 7 View Figure 7 ) are slightly curved, but the curvature is more evident in the rostral direction. The ribs are well ossified and narrow along their length, but slightly expanded at their proximal, small, articulatory heads. Ventrally, they do not reach the margin of the body. There is an unclear pattern between the last centrum bearing a pair of ribs ( Fig. 7 View Figure 7 ), which is placed anterolaterally to the first anal pterygiophore, and the first centrum with a complete haemal arch and spine that is positioned above the third anal pterygiophore ( Figs. 7 View Figure 7 and 8 View Figure 8 ). Between those centra, there are three vertebrae with ventrolateral structures that resemble parapophyses more than haemal arches. Each is associated to an elongate, median structure that is bifid at its proximal head and ends distally in two elongate processes extending between the anal pterygiophores. If these are modified ribs, then they occupy a median position, but if these are modified haemal spines, then they are unique, because they are bifid distally. I have not observed similar structures in other fossil teleosts.
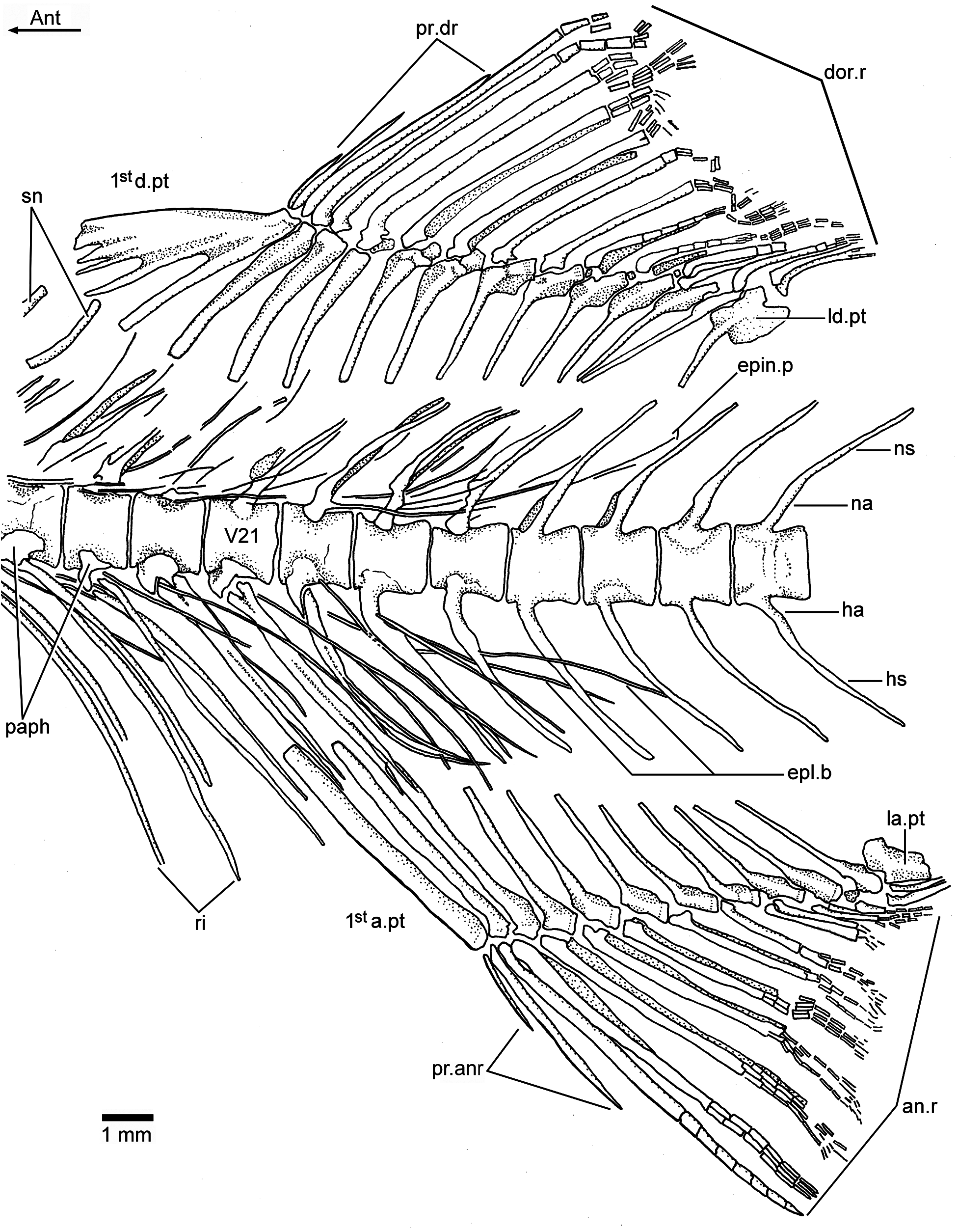
A complete series of about 16 supraneural bones ( Figs. 2a– c View Figure 2 , 3b View Figure 3 , and 6a View Figure 6 ) extends between the posterior part of the cranium and the processes of the first dorsal pterygiophore. The anteriormost supraneurals are larger and expanded anteroposteriorly than the following ones, which become narrower and smaller posteriorly. The supraneurals are slightly sigmoid or curved. They are placed above or just between the tips of the neural spines, but do not extend into the space between neighboring spines.
The epineural processes of the neural arches ( Figs. 2a–c View Figure 2 , 3b View Figure 3 , 6a–b View Figure 6 , and 7 View Figure 7 ) extend along the abdominal region ending close to the end of the dorsal fin. The last pair may arise from the neural arch of vertebra 22 or 23. The epineural processes are long, extending laterally along the space occupied by five or more centra. They are thin, heavily ossified, and curve posterodorsally, following the angle of the neural spines, with the exception of the last ones, which lie closer to the dorsal margin of the centra than to the tips of the neural spines.
A short series of epipleural bones ( Figs. 7 View Figure 7 and 8 View Figure 8 ) lies laterally to vertebrae 18 to 26, in the hypaxial musculature. The epipleurals are very thin, heavily ossified, and extend ventrocaudally to the ventrolateral surface of the last ribs, haemal arches and spines. They can be as long as the epineural processes.
Ontogenetic development of vertebral centra: Each adult abdominal centrum of Ebertichthys ettlingensis is formed by a chordacentrum surrounding and constricting the notochord and an autocentrum around the chordacentrum. In contrast, each adult caudal centrum is formed by the chordacentrum, the autocentrum, and the dorsal and ventral arcocentra that are fused to the autocentrum producing a compact vertebra. In young specimens of Ebertichthys ettlingensis , the caudal centra are unusual, because they present a pseudo-diplospondyly. Each centrum presents two small chordacentra or one large chordacentrum. The two hemicentra are separated vertically, and early in ontogeny they fuse into a single chordacentrum. In contrast, the only centrum that forms a chordacentrum in certain vertebrae presents a vertical separation that is lost early in ontogeny, so that only one centrum is observed ( Fig. 9a and b View Figure 9 ). This pseudo-diplospondyly is still observed occasionally in some autocentra showing a vertical “fracture” separating incompletely anterior and posterior regions in a centrum.
A series of caudal chordacentra ( Fig. 8b View Figure 8 ) is illustrated in a specimen of 34.7 mm standard length of Ebertichthys n. gen. A few centra show an elongate vertical gap on the lateral wall of the chordacentra. Each caudal chordacentrum is proportionally large, but its size reduces strongly caudally. Hypurals 1 to 4 are supported each by a chordacentrum, revealing a polyural condition in early ontogeny. Ural chordacentra 1 and 2 fuse during ontogeny, then are surrounded by the autocentrum producing a compound ural centrum 1 + 2 that articulates with hypurals 1 and 2. Then, the three elements become fused (see below).
Pectoral girdles and fins: The bones of the pectoral girdle and fins ( Figs. 3b View Figure 3 , 6a and c View Figure 6 ) are very well preserved in a few specimens, including the holotype, and this permits a detailed description. The posttemporal is a large bone with an almost rectangular body and a long and broad dorsal process articulating with the cranium. The ventral arm is apparently short because it was not observed in any of the specimens due to the position of the preserved bone. The main lateral line apparently is positioned along the main body of the posttemporal and exits at its posteroventral margin. No sensory tubules have been observed, only the main canal.
The supracleithrum ( Figs. 3b View Figure 3 , 4b View Figure 4 , and 6a View Figure 6 ) is a narrow, long bone extending from just at the level of the dorsal margin of the opercle to the level of the subopercle, below the suture between the opercle and subopercle and far below the level of the vertebral column. Its posterior margin is slightly crenulated in JME ETT 24. The lateral line is placed in the upper middle of the bone and exits about the level of the vertebral column.
The cleithrum ( Figs. 3b View Figure 3 and 6a View Figure 6 ) is a compact, heavily ossified bone with a short dorsal limb, a slightly expanded posteroventral corner, and a narrower, short ventral limb inclined anteroventrally. The anteromedial flange of the cleithrum is expanded, almost rounded, giving the bone a characteristic shape, unique among Jurassic teleosts.
Three postcleithra ( Figs. 3b View Figure 3 and 6a View Figure 6 ) are present. The first one is placed at the junction between the supracleithrum and cleithrum, the second one is partially covered by the posteroventral expansion of the cleithrum, and the third one is ventral to postcleithrum 2. Postcleithra 1 and 2 are ovoid shaped, whereas postcleithrum 3 is styliform.
The scapula and coracoid ( Fig. 3b View Figure 3 ) are preserved in the holotype, but not the proximal and distal radials. The coracoid is slightly L-shaped and ends anteriorly in a moderately broad and straight symphysis. Four proximal radials and a few distal radials are preserved in JME ETT 132 a ( Fig. 6c View Figure 6 ) .
The pectoral fin ( Figs. 2a–c View Figure 2 , 3b View Figure 3 , and 6b–c View Figure 6 ) has a low position on the flank, near to the ventral margin of the body. The total number of pectoral rays is unknown, but specimens JME ETT 132a and JME ETT 148 have 15 rays preserved; the first one is spine-like and thicker than all other rays ( Fig. 6c View Figure 6 ), and with scarce segmentation only at its distal end. This ray is fused with the propterygium. All other rays have very long bases and are only distally segmented and branched. The inner rays become progressively shorter.
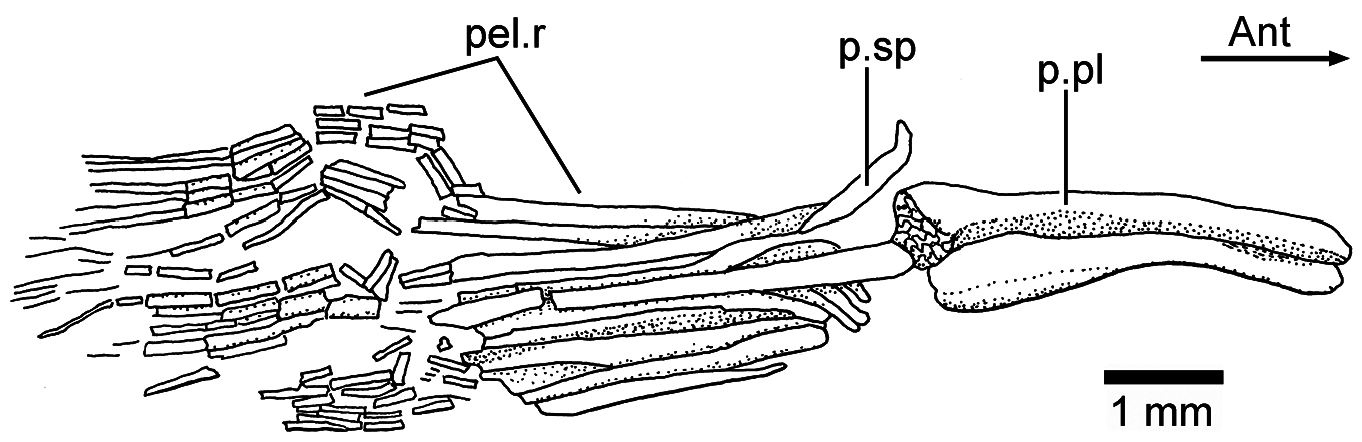
Pelvic girdles and fins: The pelvic girdles ( Figs. 2a–c View Figure 2 , 6a View Figure 6 , and 10 View Figure 10 ) are well preserved in JME ETT 132 . The basipterygium (triangular pelvic plate) presents a thickened lateral margin that expands anteriorly, while the inner region is thinly ossified bone. The posterior part of the basipterygium is thick and retains a large core of cartilage. A posterior process is apparently missing. No proximal radials have been observed. At least a short splint and nine rays are preserved in JME ETT 132 . A short splint and disarticulated rays are preserved in the holotype (JME ETT 108 a). Similar to the pectoral rays, the pelvic rays have very long bases and are only distally segmented and finely branched .
Dorsal and anal fins: The dorsal fin has three procurrent rays and 12 principal rays. The first two procurrent rays are very thin and short; the third one is almost the half of the size of the first principal dorsal ray, which is only segmented and is the largest of the series of rays. All principal rays have long bases and are finely segmented and branched distally. The first dorsal pterygiophore ( Figs. 7 View Figure 7 , 9a View Figure 9 , and 11a View Figure 11 ) expands anteroventrally and has one to three processes that are preceded by a flat, almost rectangular bony flange that gives a characteristic shape to the first pterygiophore. This element supports the procurrent rays, whereas the second pterygiophore supports the first principal dorsal ray. Pterygiophores 2–4 are of similar length, and they do not project ventrally between the neural spines. Pterygiophores 5–12 decrease slightly in size and thickness posteriorly, and the last pterygiophore is markedly expanded, with a moderately long and narrow base. As usual in teleosts, the last pterygiophore bears two small rays, but these rays articulate in different positions with the pterygiophore ( Fig. 11b View Figure 11 ). The basal portion of the pterygiophores, except the first and last ones, have a lanceolate aspect, with thin, small anterior and posterior bony flanges. They are apparently fused with the middle portion or middle radial, whereas the distal portion or distal radial is partially ossified in some of the last pterygiophores ( Fig. 7 View Figure 7 ).
The anal fin has, commonly, two or three procurrent and 11 principal anal rays that are supported by 11 pterygiophores. The procurrent rays ( Fig. 7 View Figure 7 ) are comparatively much shorter than the first principal, which is only segmented. All principal rays have long bases and are distally segmented and finely branched. The first anal pterygiophore is a long, broad, and flat rectangular bone that may support only the procurrent rays or the procurrents and part of base of the first principal ray. The next two pterygiophores are slightly narrower and shorter than the first one, and the remaining pterygiophores are much shorter and do not reach the distal tips of the haemal spines. They are thinner than the first two pterygiophores, and because of the shape of their bases, they are interpreted as formed by fusion with the proximal and middle radials. The distal radials are mostly lost – probably because they were cartilaginous. The last anal pterygiophore is broadly expanded distally and bears two minuscule rays in comparison to the first principal rays.
Caudal fin and endoskeleton: The caudal fin and endoskeleton are preserved in several specimens, so that a detailed description can be presented and intraspecific variation reported. The caudal fin ( Figs. 2a–c View Figure 2 , 12 View Figure 12 , and 13) is deeply forked with very short middle principal rays in comparison to the long, leading marginal rays.
Five preural vertebrae support the caudal rays. All these vertebrae are characterized by their smooth surfaces, and their broad dorsal and ventral arcocentra are fused to their respective centrum in larger specimens. They may retain remnants of cartilage in some vertebrae. The neural spines of preural vertebrae 2–5 are slightly expanded, and they have a central core of cartilage surrounded by a thin perichondral ossification and slightly developed anterior and posterior bony flanges. In the vertebrae that are completely preserved, it is possible to observe the anterior elongate processes at the base of neural spines 1 to 4. Neural spines 2 to 5 are long, whereas the neural spine of preural centrum 1 is shorter than the preceding ones. The haemal spines of preural centra 1–5 are broader than their respective neural spines. However, the haemal spine of preural vertebra 5 is narrower. The anterior processes located between the base of the haemal arch and spine are slightly rounded. The haemal spines of preural vertebrae are chondral elements; some retain a core of cartilage.
In most specimens the neural and haemal arches of preural vertebra 1 are fused to their centrum, but in specimen JME ETT 108a, the haemal arch is still unfused. A complete neural arch, with its spine, is present on preural centrum 1. This spine is shorter than the preceding spines. The haemal arch and its broad parhypural are fused to the centrum in most specimens (e.g., Fig. 13 View Figure 13 ). A hypurapophysis on the lateral wall of the haemal arch of preural centrum 1 was not observed in any specimen.
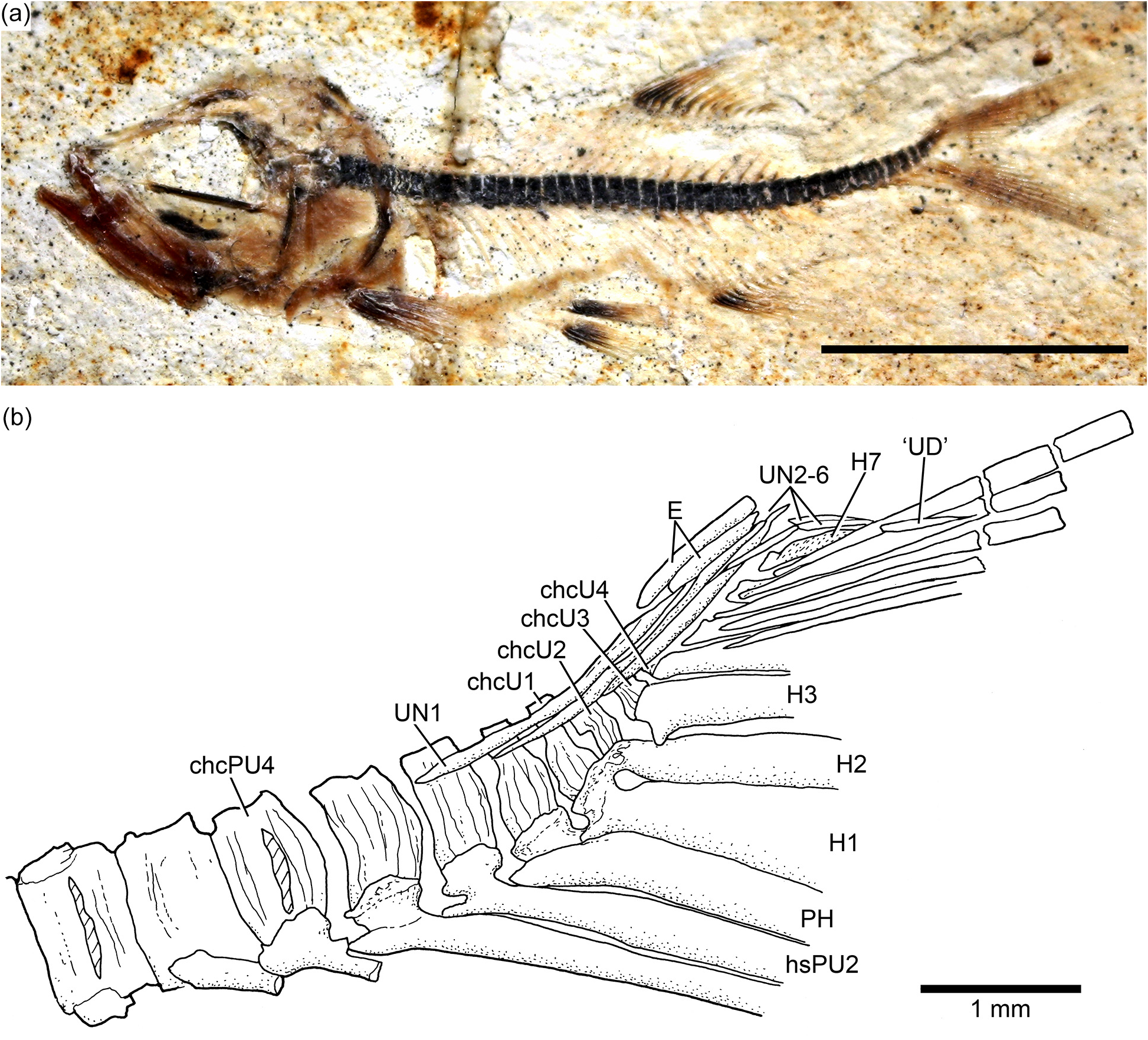
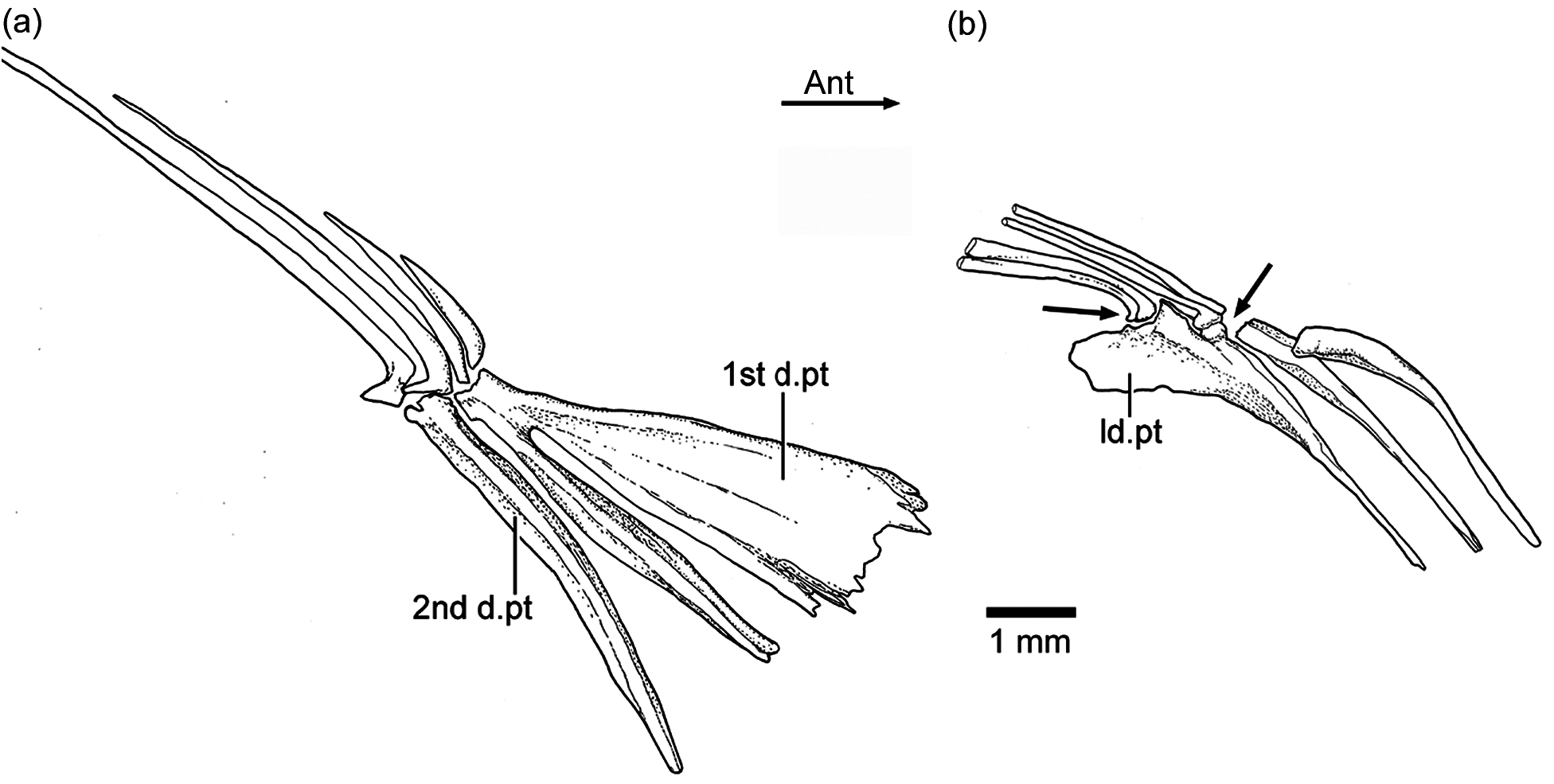
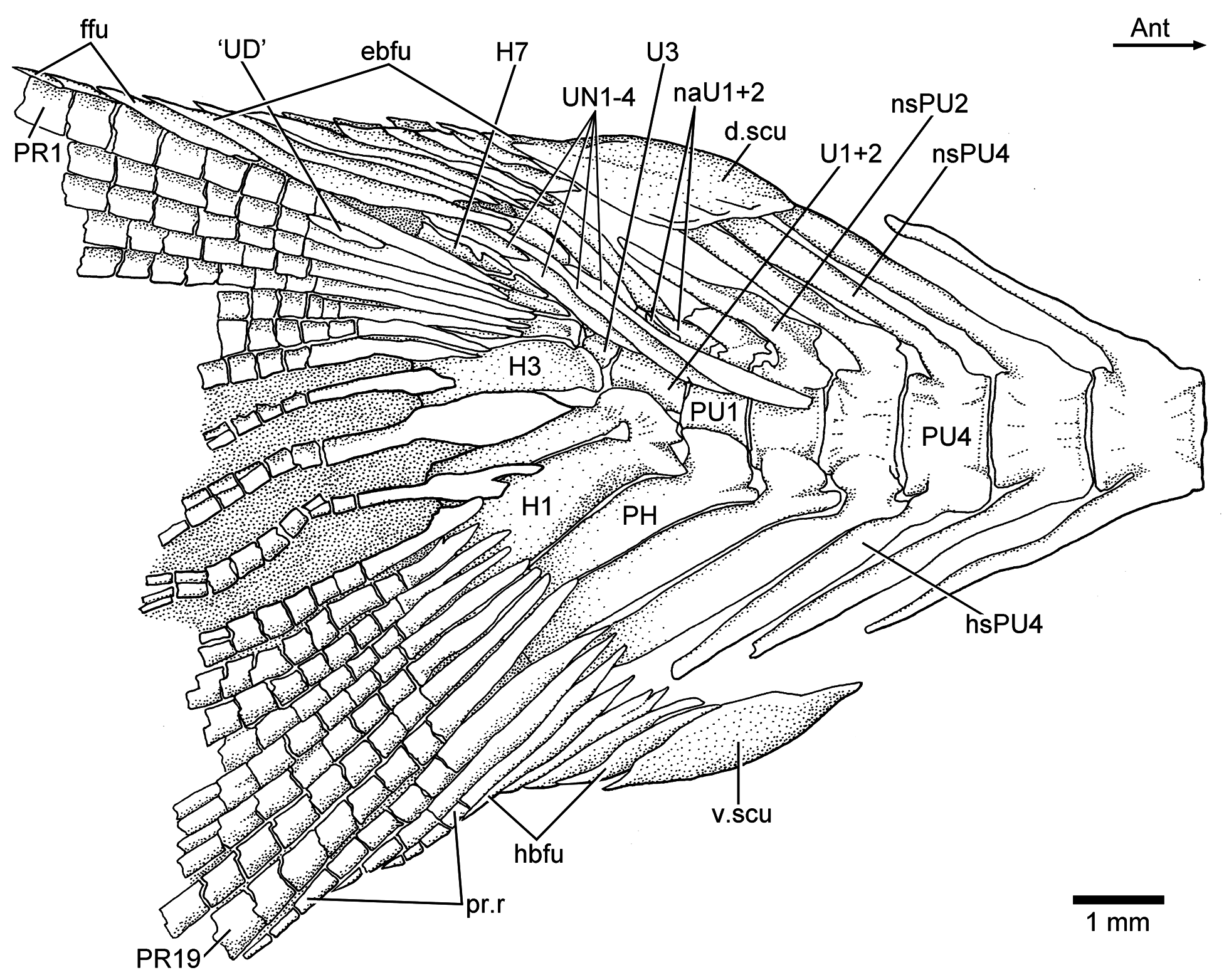
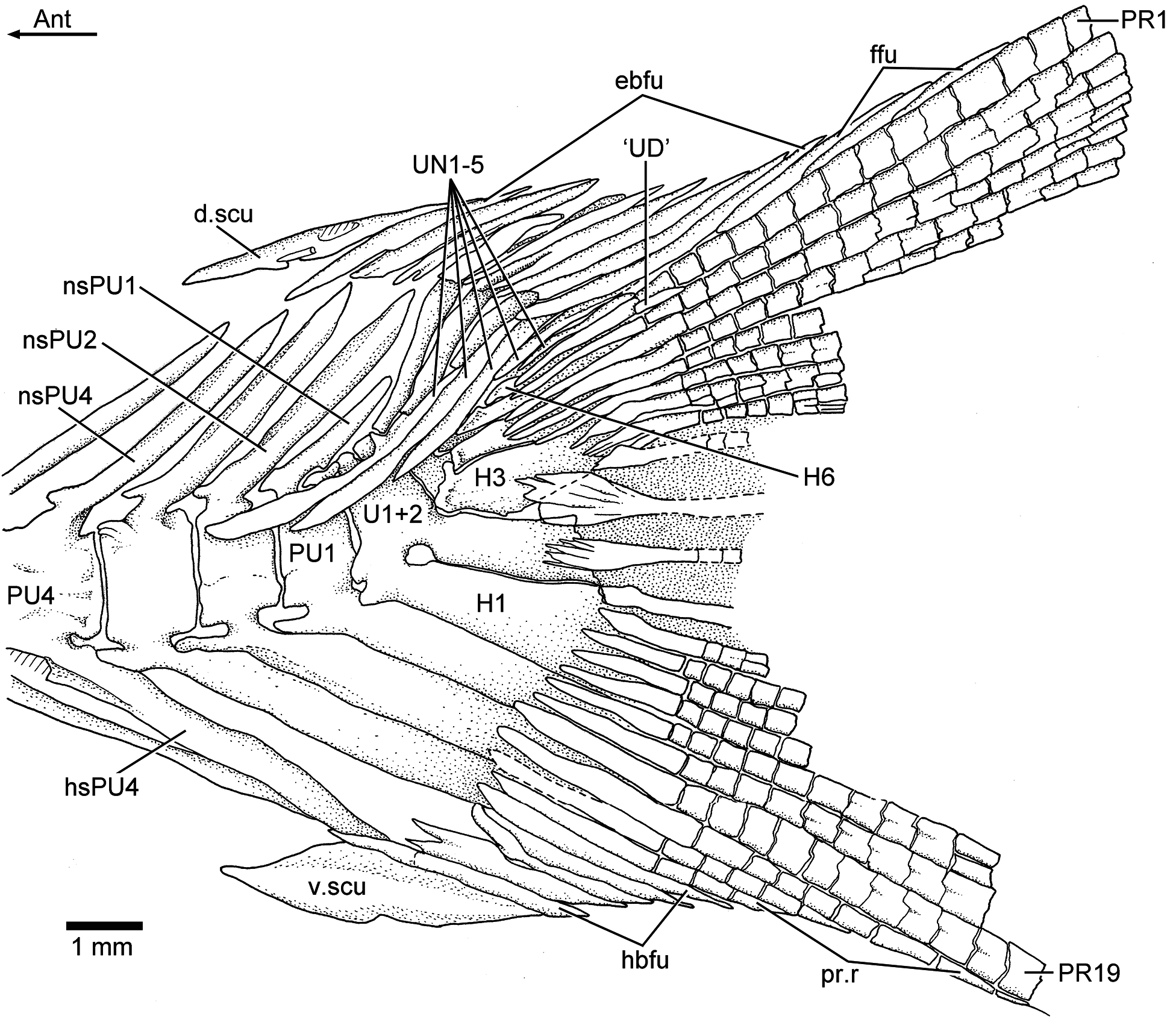
Three or four ural centra (of the polyural terminology) are associated with their respective hypurals. The first ural centrum that bears hypurals 1 and 2 results from the fusion of ural centra 1 and 2 ( Figs. 9 View Figure 9 , 12 View Figure 12 , and 13). Ural centra 3 and 4 are associated with hypurals 3 and 4, respectively. An arch and its spine and an incomplete arch are present above ural centrum 1 + 2. A third incomplete arch is present in other specimens ( Fig. 13 View Figure 13 ).
The complete number of uroneurals is unclear. Some specimens have four ( Fig. 12 View Figure 12 ) or five ( Fig. 13 View Figure 13 ) or possibly six (JME ETT 132a) uroneurals preserved. The first uroneural, the longest of the series, extends anteriorly, reaching the lateral surface of preural centrum 2. The second uroneural reaches the lateral surface of preural centrum 1, and the third uroneural reaches the lateral surface of ural centrum 1 + 2. The fourth uroneural is short, reaching anteriorly the base of hypural 6 or 7. There are other two smaller, elongate elements that I interpret as uroneurals 5 and 6. The first uroneural may have a small membranous outgrowth. There are three elongate epurals occupying the space between the neural spines 1 and 2 and the uroneurals; the first one is the longest.
At least nine hypurals are present. Hypurals 1 and 2 are continuous at their bases, and they fuse to the autocentrum of ural centrum 1 + 2 during ontogeny. Hypural 1 is the largest element of the series, and hypural 2 is comparatively narrow. Hypurals 2 and 3 are contiguous to each other, so that the hypural diastema is very narrow or non-existent. Hypural 3 is the broadest among hypurals 3–9, which decrease in size posterodorsally. Hypurals 2 and 3 are partially covered by the expanded bases of the middle principal rays (e.g., 10 and 11).
There are eight or nine epaxial basal fulcra, three fringing fulcra, 19 principal rays, three hypaxial procurrentsegmented rays, and four or five hypaxial basal fulcra ( Figs. 12 View Figure 12 and 13 View Figure 13 ). One long and slightly fusiform or ovoid dorsal scute and a ventral scute precede the epaxial and hypaxial series of basal fulcra, respectively.
The anterior epaxial basal fulcra in specimen JME ETT 108a are apparently formed by paired elements ( Fig. 12 View Figure 12 ); however, the anterior epaxial basal fulcra are unpaired but with two separate, ventral projections in other specimens. The basal fulcra are elongate, leaf-like elements that expand laterally, partially covering the next fulcrum. The three elongate fringing fulcra lie on the dorsal margin of the first principal ray. They can be elongate and of similar size ( Fig. 13 View Figure 13 ), or the first one is comparatively longer than the other two ( Fig. 12 View Figure 12 ). The bases of the posteriormost basal fulcrum and the first principal caudal ray produce an angle as described for other fishes, mainly teleosts ( Arratia, 2008).
Ten principal caudal fin rays are articulated with at least hypurals 3 to 6. Hypural 6 supports the first principal ray, and the other nine rays are supported by hypurals 3 to 5. Nine principal rays are supported by hypurals 1 and 2, parhypural, and haemal spine 2. The articulation between segments of the leading rays is mainly Z- or step-like, whereas the articulation between segments of the inner principal rays is mainly straight. Some of the bases of the middle principal caudal rays are broken, but those that are preserved show that they were expanded ( Figs. 12 View Figure 12 and 13 View Figure 13 ). Dorsal processes associated with the bases of the middle principal rays of the upper lobe have not been observed in the available material.
One elongate “urodermal” (sensu Arratia and Schultze, 1992) lies between the bases or close to the bases of the first and penultimate principal rays. The “urodermal” is a thin, oval bone ( Figs. 12 View Figure 12 and 13 View Figure 13 ).
Scales: Thin, large cycloid scales with circuli at their anterior field ( Figs. 2a–b View Figure 2 , 3a–b View Figure 3 , and 6 View Figure 6 ) cover the whole body. No radii are observed. The scales are so large that approximately six rows of scales covered the skin of the flank below the vertebral column and between pectoral and pelvic fins. No remnants of scales are observed on the fin rays.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.