
Panoploscelis scudderi Beier, 1950
|
publication ID |
https://doi.org/10.5252/zoosystema2019v41a17 |
|
publication LSID |
urn:lsid:zoobank.org:pub:FAF29578-704B-48ED-A426-A345C49DEFB3 |
|
DOI |
https://doi.org/10.5281/zenodo.3729218 |
|
persistent identifier |
https://treatment.plazi.org/id/03F687D8-FFF8-FFDD-1D76-FB34ADC5F52F |
|
treatment provided by |
Felipe |
|
scientific name |
Panoploscelis scudderi Beier, 1950 |
| status |
|
Panoploscelis scudderi Beier, 1950 .
Panoploscelis scudderi Beier, 1950: 112 .
Panoploscelis angusticauda Beier, 1950: 113 , n. syn.
TYPE MATERIAL. — “Coll. Br. V. W. [Printed], Rio Branco, Peru [manuscript; in reality Brazil], Staudinger [manuscript]; 24.875 [manuscript]; Type [printed, red label], Panoploscelis armata Scudd. [manuscript] determ. Karny [printed]; Panoploscelis scudderi n. sp. Type ♂ [manuscript]; NHM, Wien, Austria, high quality pictures examined. — Type [printed, red label]; Natt: 12; Rio-Negro [manuscript]; Panoploscelis armata Scudd. [manuscript]; Panoploscelis angusticauda n. sp. Type! ♀ [manuscript] det. Beier [printed], NHM, Wien, Austria, high quality pictures examined.”
TYPE LOCALITY. — Rio Branco, Brasil.
OTHER MATERIAL EXAMANED. — French Guiana. Planète revisitée Guyane 2015, Monts Tumuc-Humac , Massif du Mitaraka , Layon D, 54.4509°O, 2.2357°N, 280 m a.s.l. GoogleMaps – 54.4517°O, 2.2338°N 293 m a.s.l., 23.II-10.III.2015, nuit, 1 ♂, 1 ♀, MNHN. GoogleMaps — Planète revisitée Guyane 2015, Monts Tumuc-Humac, Massif du Mitaraka , vers sommet en Cloche , 54.4541°O, 2.2349° N, 370 m GoogleMaps – 54.4646°O, 2.2329°N 470 m a.s.l., 23.II-10.III.2015, nuit, 1 ♀, MNHN. GoogleMaps These specimens were reared, and F1-F2 generations were also examined for the present work to assess the variability of diagnostic characters.
DISTRIBUTION. — South America: North Brazil, Guiana, French Guiana.
DESCRIPTION
Male
Head. Fig. 2A, B View FIG . All as in female. Fastigium verticis spineshaped, reaching antennal sockets. Fastigium frontis spineshaped. Carina lateralis interna distinct at the basis, carina lateralis externa complete. Scapus with one distal inner spine and one distal dorsal tubercle.
Thorax. Fig. 2B View FIG . Anterior margin with a distinct median tubercle; in addition to the two transverse sulci, sagittal sulci at least distinct at the level of posterior transverse sulcus (as in female); posterior margin broadly rounded with an inconspicuous median emargination of the rim. Metazona elevated.
Legs. All as in female, anterior tibiae and femora longer than mid tibiae and femora. Fore femur ( Fig 2C View FIG ): distal spine of inner ventral margin perpendicular to femur axis; dorsally with 4-5 spines. Posterior femora dorsally with 5-7 spines ( Fig. 2D View FIG ); mid femora unarmed dorsally. Inner genicular lobe of fore femora unarmed. Wings. Left forewing shorter than right forewing (this very unusual asymmetry is possibly a generic character). Left mirror area triangular, right mirror somewhat oval. Anal field posterior margin of left forewing with a concavity; posterior margin of both forewing with an emargination distal to the anal field. File with c. 66 teeth. Abdomen. Suranal distal margin variable, usually with an angle (as in female), sometimes very obtuse. Cercus short, stout, with a single inner ventral spine-shaped projection.
Subgenital plate broad at basis, moderately tapering distally; with a more or less rounded V-shaped emargination between the styli.
Measurements. See Table 1 View TABLE .
Female
Female has been described under Panoploscelis scudderi and Panoploscelis angusticauda in Beier (1950, 1960 ). Variation in characters are illustrated Fig. 1 View FIG . Forewings of a female specimen from Mitaraka are illustrated in Fig. 3A, B View FIG . Female forewings of P. scudderi lack the scraper lobe pointed by Montealegre-Z et al. (2003) in females of Panoplscelis specularis Beier, 1950 .
BIOACOUSTICS
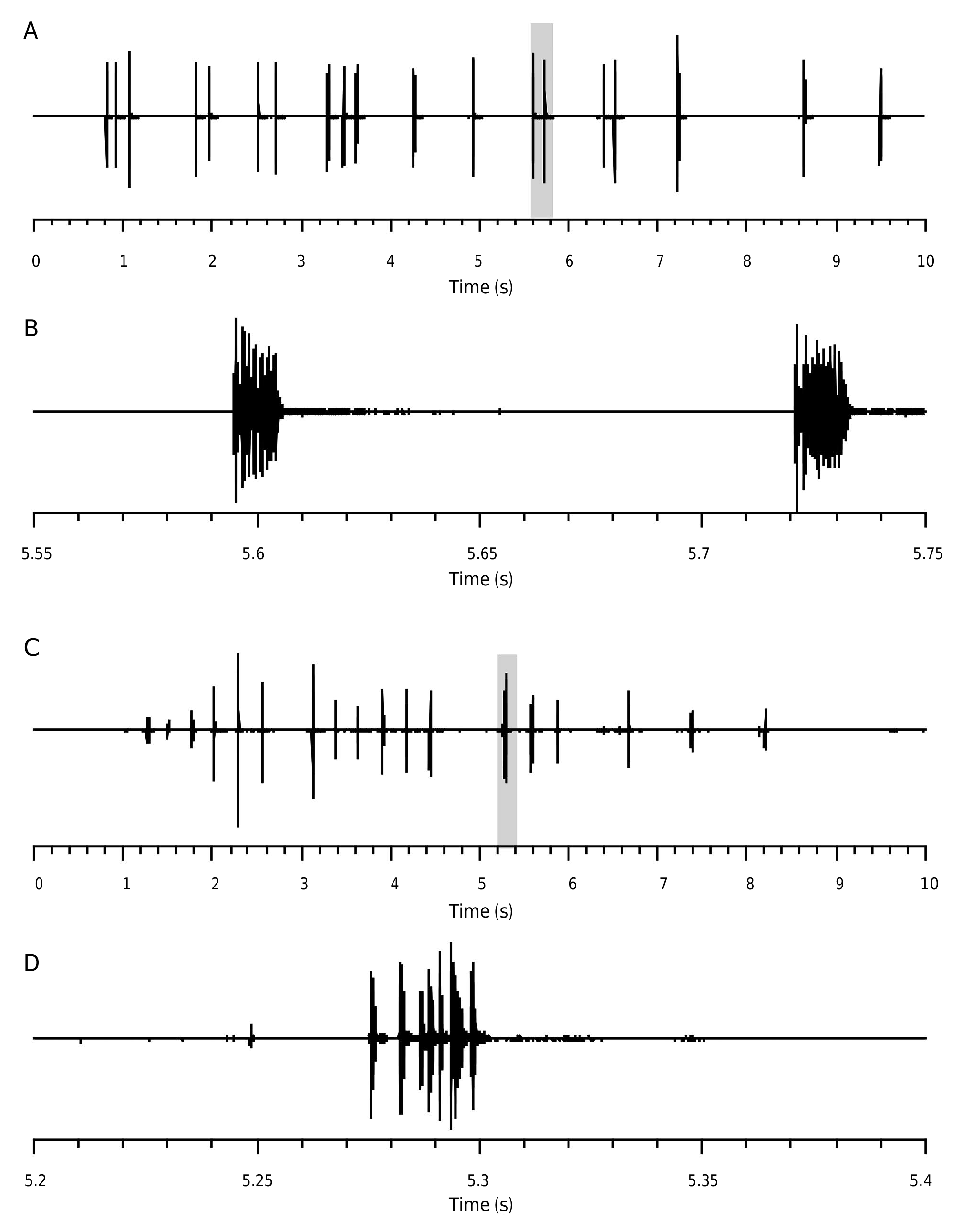
Male call ( Fig. 4 View FIG ).
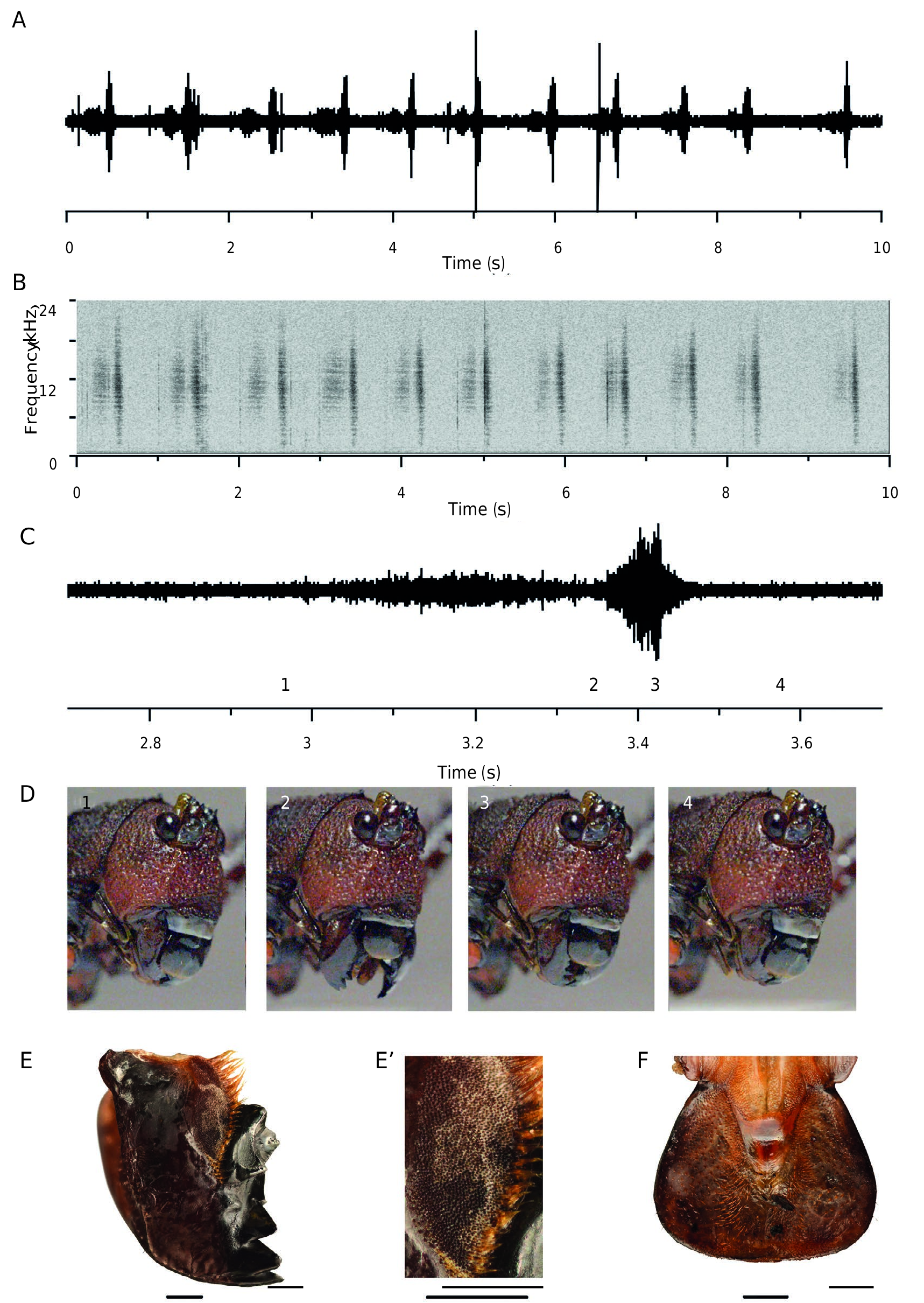
In rearing conditions, with many males and females together, the call of Panoploscelis scudderi males is very rarely heard. Single males sing only by night hours. At 24 °C, the song is made of 3-12 verses repeated every 31.7 ± 3.1 s (min: 18.44 s; max: 43.60 s). The first verse is usually softer than the subsequent ones. The duration of one verse is 7.5± 0.6 s (min: 4.4 s; max: 10.0 s). Verses are made of 29.4±1.9 phonatomes (min: 18; max: 34) repeated every 251±2 ms (min: 233 ms; max 271 ms). The duration of one phonatome is 60±2 ms (min: 30 ms; max: 87 ms). At 24°C, a first peak was recorded at c. 10.3-11.0 kHz, and a secondary broad peak was recorded at 30.5 kHz.
Isolated males seldom produce isolated verses (1-3 per nights).
Female call
In our initial rearing conditions (30 adult pairs in a 0.3 m 3 cage at 24°C by night), we failed to record any call from females (at F3 generation, a total of 50 adult females have been reared). Since males only rarely sing in the same conditions, this lack of spontaneous calls might simply be linked to specimen density. Therefore at F4 generation, subadult males and females were separated and kept in isolation for two months after their last molting. Under these conditions, males were calling very frequently (at least one call every 10 min for 25 males together in a cage). We were nevertheless unable to record any song from females, even with overnight recordings (8 full night recordings). Since females may only respond to male call, we play-backed male calls (one call every 10 min, during the whole night): no responses were recorded (two overnight recordings). We than checked whether females responded to males during the mating process: we put virgin females together with males in one single empty cage and imaged/recorded them during the whole night. Under these conditions, all females mated within the first 30 minutes, and neither males nor females produced any song (Supplementary video 3).
These data suggest that call of P. scudderi females is not required in the reproductive behavior of that species. It cannot be ruled out that P. scudderi females do not call at all since the only tegminal sounds we recorded were produced as protest signals.
Interstingly, P. scudderi females lack the scraper lobe pointed by Montealegre-Z et al. (2003) in P. specularis ( Fig. 3A View FIG ). This might speculatively suggest that intraspecific female signaling might be less developed in P. scudderi .
Male and female protest signals
Both males and females respond to air puff, water spraying or simple touch by lifting their forelegs and repeated protest signals ( Fig. 3C View FIG ). These protest signals are produced by tegminal stridulatory organs as well as mandibles/labrum.
At 24 °C, male tegminal protest signal ( Fig. 5 View FIG AB) is made of phonatomes isolated or grouped (2-3) repeated every 420±78 ms (min: 108 ms; max: 1335 ms). The frequency peaks at 9.4 kHz, a frequency significantly lower than male call. At 24°C, female tegminal protest signal ( Fig. 5 View FIG CD) is made of verses repeated every 348±56 ms (min: 246 ms; max: 836 ms). Verses are made of 5±1 phonatomes (min: 2; max: 12) of irregular duration and spacing. The power spectrum
of female tegminal protest is broad and displays a double peak: one at 10 kHz and another at 14 kHz.
Male and female mandibular protest signal ( Fig. 6 View FIG , Supplementary video 1)
Mandibular protest signals are similar in males and females. These are the first protest signals produced when a specimen is stimulated, the tegminal signal requiring more stimulation. This signal has two phases: the first phase is produced while opening the mandibles and up lifting the labrum by clypeus retraction, the second is produced by the return to the resting state. The total duration of a cycle is very variable, lasting c. 0.5 s, with a first phase usually longer than the second one. These signals are repeated every 2.3± 0.2 s (min: 0.7; max: 3.6). The power spectrum of this signal displays a broad peak between 10.3 kHz and 14.6 kHz
Protest signals appear broader and emitted at lower frequencies than male call. This might speculatively be linked to the correlation between signal valence and signal frequency in intraspecific communication of some rodent species were lower frequencies are associated to unpleasant situations ( Knutson et al. 2002).
Comparison of P. specularis and P. scudderi male call
Whereas the call of P. specularis males is made of two phonatomes separated by c. 130 ms and regularly repeated every c. 3 s, the song of P. scudderi is more complex in structure, with phonatomes organized in verses (see above). Alternatively, since P. scudderi sometimes produces isolated verses, one cannot rule out that P. specularis could do the same, and the full call of this species with organized verses may speculatively have been missed. The frequency of the maximally intense peak is relatively lower in P. scudderi than in P. specularis (10- 11 kHz vs. 11-13 kHz).




| NHM |
United Kingdom, London, The Natural History Museum [formerly British Museum (Natural History)] |
| MNHN |
France, Paris, Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Pseudophyllinae |
|
Genus |
Panoploscelis scudderi Beier, 1950
| Hugel, Sylvain 2019 |
Panoploscelis scudderi
| BEIER M. 1950: 112 |
Panoploscelis angusticauda Beier, 1950: 113
| BEIER M. 1950: 113 |