
Paleopiquitinga brasiliensis, De, Francisco J., 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.194942 |
|
DOI |
https://doi.org/10.5281/zenodo.6200377 |
|
persistent identifier |
https://treatment.plazi.org/id/03F787AF-FFAC-8911-D582-FDD87D6AC7CC |
|
treatment provided by |
Plazi |
|
scientific name |
Paleopiquitinga brasiliensis |
| status |
comb. nov. |
† Paleopiquitinga brasiliensis , comb. nov.
( Figures 1–9 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 )
Selected synonymy
† Knightia brasiliensis Woodward, 1939: 450 –451, Pl. 16, figures 1 e 2. † Knightia brasiliensis: Santos, 1946: 1 (quotation).
† Knightia brasiliensis: Schaeffer, 1947: 20 (quotation).
† ‘Knightia’ brasiliensis : Grande, 1982: 15 (comments on taxonomic position). †‘ Knightia View in CoL ’ brasiliensis: Grande, 1985: 319 (comments on taxonomic position).
Diagnosis. as for genus.
Referred material. DGM 32, DGM 33, DGM 45–59.
Remarks. Woodward (1939) figured only two specimens (DGM 32 and 33) in his original description, but he did not designate a holotype. It is not possible to determine which specimens he effectively used in the original description. Therefore, DGM 33 is considered herein as lectotype and all remaining specimens collected by the late geologist Josalfredo Borges, in 1936, as paralectotypes.
Locality and stratigraphy. The precise site where the specimens were collected is uncertain. According to Woodward (1939), it was on left bank of Parnaíba River at Vila de Nova Iorque, State of Maranhão. However, Oliveira (1936) furnished more complete information based on field notes of the geologist Josalfredo Borges who collected all specimens available. He described the outcrop as having an extension of approximately 35m and composed of a sequence of five distinct beds. In a lower bed (ca. 0,8m), corresponding to the third (top to bottom) bed of that sequence, composed of greenish shales, many smallsized fishes were found, mainly † Paleopiquitinga brasiliensis .
Oliveira (1936) reported “lots of specimens” were obtained from the Nova Iorque beds. So far, 65% of this collection have been assigned to the clupeid † Paleopiquitinga brasiliensis ( Melo et al., 2005) .
Although Melo et al. (2005) claimed MCTer/DNPM housed all material coming from the Nova Iorque beds, according to Grande (1982, 1985) eight specimens of † Knigthia brasiliensis are currently deposited in the paleontological collection of the Natural History Museum of London. This may be due to the fact that the material was donated to institution by the late Eusebio Paulo de Oliveira (Director of DNPM in the 1930s) in a reward for valuable contributions by Arthur Smith Woodward to Brazilian paleontology, particularly in describing taxa coming from Maranhão. More recently, some researchers have obtained stratigraphic data from pollen indicating a Pliocene age for the Nova Iorque beds ( Lima, 1991; Góes & Feijó, 1994).
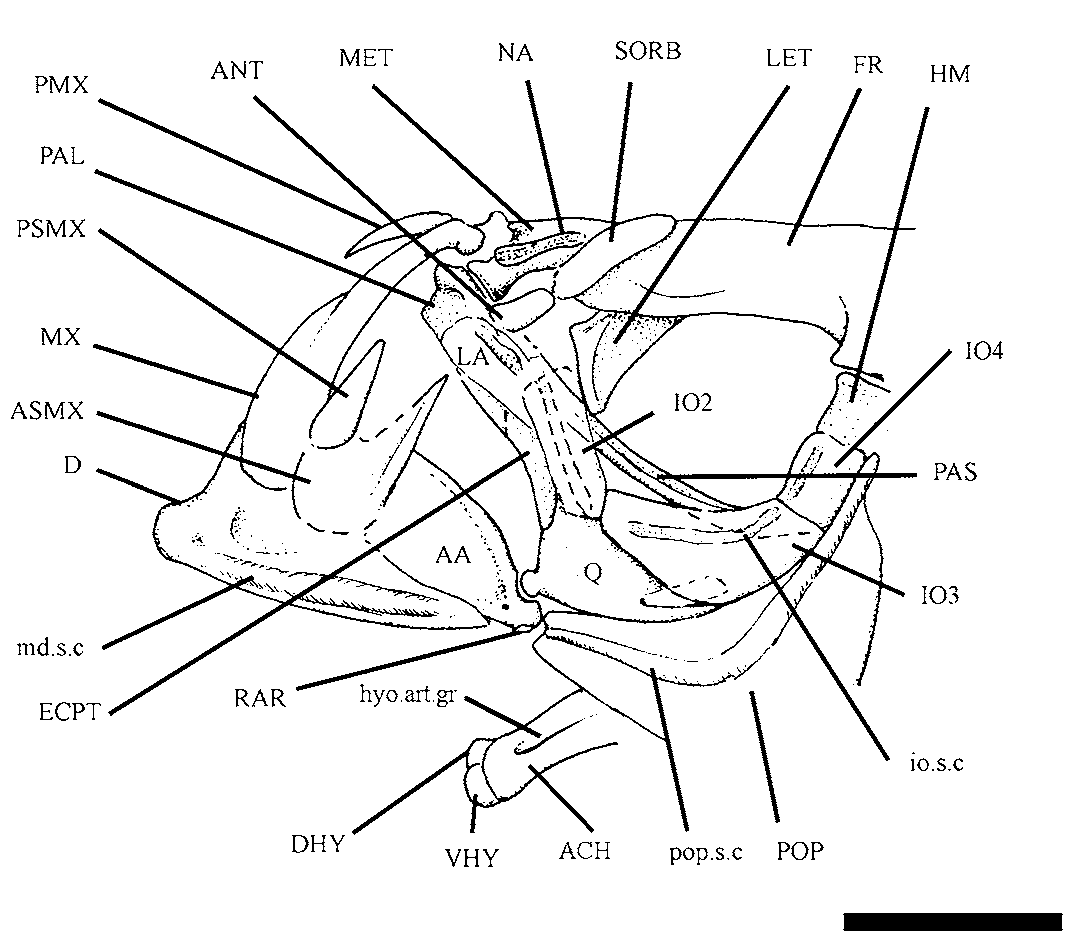
Descriptive morphology. Cranial bones. Although many specimens were examined, DGM 38, 44 and 50 show the best skull preservation ( Figs. 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). The ethmoidal region comprises a relatively elongate mesethmoid bone ( Fig. 3 View FIGURE 3 ) bearing a pair of prominent bell-shaped lateral processes forwardly inclined to meet the palatine. There is a short and prominent anterodorsal spine-like process separating deep lateral fossae for lodging the ascending process of premaxilla and articular head of maxilla.
The vomer is partially known in lateral view ( Fig. 2 View FIGURE 2 ). It is edentulous. The sharp posterior process is lodged in an anteroventral groove of parasphenoid placed anterior to lateral ethmoid.
The lateral ethmoid ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 5 View FIGURE 5 ) is a delicate anvil-shaped bone corresponding to a large perichondral shield. It forms the anterior limit of the orbit and tapers in a sharp posteroventral process. The bone does not reach the level of the parasphenoid so that among mesethmoid, lateral ethmoid and vomer there is a large space probably filled with cartilage in life.
The nasal ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ) is a short and tubular bone flanking the mesethmoid along most of its extension. Practically, it is reduced to a neurodermal component, showing a thin longitudinal tubular relief containing the anteriormost portion of the supraorbital sensory canal.
The skull roof is relatively long and wide, and lacks anterior and posterior fontanels at the midline. A pair of frontals ( Figs. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ) corresponds to the main tectorial component of the orbit. Each one is narrow above the orbit, becoming widely expanded behind it to produce an extensive temporal shield, which covers a large part of the otic region and separates autosphenotic of pterotic externally. The midline region between frontals is depressed and smooth whereas underlying parts are deeper and ornamented with delicate longitudinal ridges.
Frontal extends backwards until reach supraoccipital and parietals. Each frontal forms together with parietal the medial limit of a deeply excavated temporal fossa.
Anterior to orbit, the supraorbital sensory canal extends along the frontal in a deep groove. Above the orbit, the trajectory of canal is hardly noted on surface by an obscure longitudinal tube. Posterior to orbit, the canal is totally bone-enclosed and does not show any exit on surface.
The parietals ( Figs. 2 View FIGURE 2 , 4 View FIGURE 4 , and 5) are transversely subrectangular bones weakly ornamented of striae and grooves radiating from the centers of growth. Medially, they are separated by an outgrowth of supraoccipital and each one meets the pterotic laterally. The parietal forms, together with the pterotic, the posterolateral limit of the temporal fossa.
The posterolateral portion of parietal meets the epioccipital through a long and sinuous contact. Both contribute to the limits of a pre-epioccipital fossa whose structural pattern resembles that of the alosine Brevoortia (pers. obs.). The supratemporal commissure extends along a great part of the parietal near surface continuing into supraoccipital medially.
The autosphenotic ( Figs. 2 View FIGURE 2 , 4 View FIGURE 4 , and 5) is a large and falcate bone forming the posterodorsal limit of the orbit. Posterolaterally the bone produces a deflected autosphenotic spine. This bone contributes with a small part for the anterior hyomandibular facet. The site corresponding to the dilator fossa is very expanded and totally exposed on surface.
The pterotic ( Fig. 2 View FIGURE 2 ) is a trapezoid bone in lateral view. Medially, it meets the parietal for producing an anterior flange, which contributes to the lateral limit of the temporal fossa.
As in other clupeoids, the temporal sensory canal runs most of pterotic in a dilated bony tube near lateral border that promptly makes confluence with a large inner chamber, the so-called recessus lateralis. From this chamber ( Fig. 4 View FIGURE 4 ), there is an anterior opening for the temporal sensory canal (partially covered by frontal) that makes anastomosis with the infraorbital sensory canal. Nearby, there is a second opening, the so-called middle opening of the temporal sensory canal, which joins the preopercular sensory canal. The posterior opening of the temporal sensory canal for recessus lateralis is large and grooved anteroposteriorly.
The supraoccipital ( Figs. 2 View FIGURE 2 , 4 View FIGURE 4 , and 5) is a small subtriangular bone. It produces a very short and laminate anterior outgrowth but the supraoccipital process is unconspicuous. In DGM 35 and 50 this bone is better seen because it is slightly displaced from its usual anatomical position.
An inverted L-shaped epioccipital ( Figs. 2 View FIGURE 2 and 4 View FIGURE 4 ) meets the parietal anteriorly and supraoccipital medially. In the confluence of limbs there is a reduced articular facet for receiving the anterodorsal process of posttemporal bone. Extrascapulars were not observed in any specimen examined.
The parasphenoid ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , and 5) is only seen in the orbital region; it is rectilinear and shafted along most of its extension, except in the posterior corner of orbit where is slightly arched. This bone lacks teeth on its oral border. A low median crest for membranous interorbital septum is visible at the midline of the orbital region.
Orbital bones. The orbital region ( Fig. 2 View FIGURE 2 , 3 View FIGURE 3 , and 5) is very large, corresponding to approximately 1/3 of head length. The sclerotic ring is not preserved in any specimen examined, indicating it is composed of very delicate and flimsy structures easily loss during fossilization. Yet orbital series is complete and relatively well preserved. It is composed of supraorbital, antorbital, and six infraorbital bones.
The lachrymal (first infraorbital, Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ) is a large and spatulate bone placed anterior to orbit and meeting an anamestic and reniform antorbital ( Fig. 3 View FIGURE 3 ) dorsally.
The second infraorbital ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ) is subrectangular; it is as long as the first infraorbital. The third infraorbital ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ) is trapezoidal, with well-developed laminar portion that reaches the dorsal border of quadrate posteroventrally.
The fourth infraorbital ( Fig. 3 View FIGURE 3 ) is a small and quadrangular bony plate. Although the fifth infraorbital bone is not entirely preserved in any specimen examined, judging by the gape between fourth and sixth infraorbital bones, it was probably similar in dimensions to fourth infraorbital bone. The sixth infraorbital (=dermosphenotic) is a reduced trapezoidal bone lying on the wing-like lateral outgrowth of the autosphenotic ( Fig. 2 View FIGURE 2 ).
The infraorbital sensory canal extends along the infraorbital bones within a conspicuous bony tube near the orbital rim. There is no evidence of secondary tubules radiating from it.
The supraorbital ( Fig. 3 View FIGURE 3 ) is an elongate and fusiform bone lying on frontal laterally, in the anterodorsal corner of the orbit.
Upper jaw. The dermal bones which form the upper jaw are: premaxilla, maxilla, and two supramaxillae ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ).
The premaxilla ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ) is a small triangular bone. It lacks teeth on oral border and shows a moderate symphyseal region.
The maxilla ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ) is very similar to that of † Clupavus Arambourg, 1954 (see Taverne, 1977). It is short, with anterior half thin and elongate whereas posterior half is roughly elliptical. The anterior (cranial) process is relatively short as well as palatine condyle.
The anterior supramaxilla ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ) is a fusiform and low bone lying on anterodorsal margin of maxilla. The posterior supramaxilla ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ) is drop-like, with long and sharp anterodorsal process.
Lower jaw. The lower jaw is relatively short and deep as usual in generalized clupeoids ( Fig. 2 View FIGURE 2 and 3 View FIGURE 3 ). In lateral view, it is composed of dentary, anguloarticular and retroarticular. The dentary ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ) shows a relatively short and low symphyseal margin. Teeth are lacking on oral border. Posteriorly, the bone ascends abruptly for producing a deep coronoid process. The ventral margin of the bone is slightly convex and the mandibular sensory canal runs all its extension in a conspicuous straight bony tube.
The anguloarticular ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ) has a short anterior laminate process for fitting onto posterior incisure of dentary and a deep articular facet for quadrate. The posterodorsal margin of the bone ascends abruptly producing a deep coronoid process. An outer pore for the mandibular sensory canal is visible in certain specimens (e.g., DGM 38), a little beneath the articular facet for quadrate.
The retroarticular ( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 ) is lodged in a recess of the ventral margin of anguloarticular anterior to the short and rounded postarticular process of the mandible.
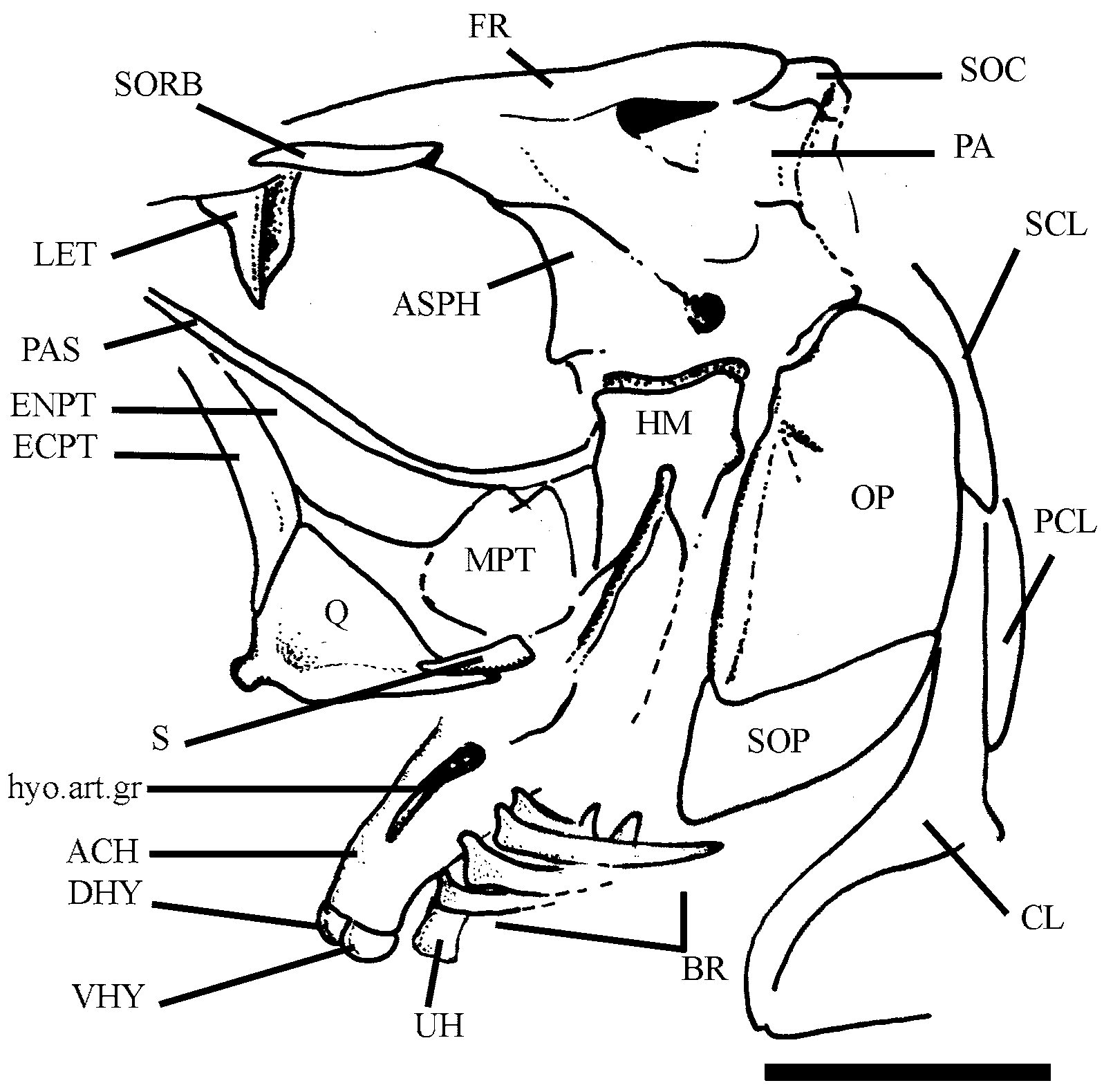
Supensorium. The mandibular suspensory is exposed in few specimens (e.g., DGM 44, Fig. 5 View FIGURE 5 ). The palatine ( Fig. 3 View FIGURE 3 ) is often hidden by the first infraorbital bone and dorsal edge of the lower jaw so that it is rarely seen. It is subtriangular, with ventral margin slightly convex and edentulous. There is a short and rounded anterodorsal process with reduced articular facet for the mesethmoid.
The endopterygoid ( Figs. 2 View FIGURE 2 and 5 View FIGURE 5 ) is a flimsy and oval shield forming the lateral floor of the orbit, flanking the parasphenoid. The underlying ectopterygoid ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , and 5) is typically boomerang-shaped. It is toothless and meets the anterodorsal margin of quadrate backwardly.
The quadrate ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , and 5) is a triangular bone, well ossified, with dorsal margin almost straight and short posteroventral process. The articular condyle of quadrate is short but prominent, and it inclines forwards a little. The quadrate-mandible joint is placed beneath the anterior half of the orbit. Dorsally, the quadrate joins the large and trapezoidal metapterygoid ( Fig. 5 View FIGURE 5 ). There is no fenestra between quadrate and metapterygoid as commonly found in alosines.
The club-like symplectic ( Fig. 2 View FIGURE 2 and 5 View FIGURE 5 ) is partially hidden by the dorsal margin of quadrate and is lodged in a short notch medially.
The hyomandibula ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , and 5) is narrow and deep, with a well-developed ventral process. It meets the otic portion of neurocranium through a unique and slightly inclined articular facet. The anterior membranous process is very reduced. The opercular process is short, stout, and placed in the upper third of the bone.
Hyoidean bones. Details of hyoidean bones ( Figs. 3 View FIGURE 3 and 5 View FIGURE 5 ) and associated structures are better observed in few specimens (i.e., DGM 44 and 47). The dorsal and ventral hypohyals are equidimensional and reniform bones, with external surface markedly convex.
The anterior ceratohyal ( Figs. 3 View FIGURE 3 and 5 View FIGURE 5 ) is approximately hourglass shaped and very compressed. It shows a deep longitudinal groove for lodging the hyoidean artery. Four short and spathiform branchiostegals, bearing ‘clupeoid projections’ (see McAllister, 1968) onto anteroventral border, insert in shallow foveae on the ventral margin of the bone.
The posterior ceratohyal (see in DGM 47) is a small and triangular bone bearing only three elongate and falcate branchiostegals. Judging by the gape between the smooth surfaces of anterior and posterior ceratohyals, the contact between those bones probably was synchondral.
A nodular interhyal (see in DGM 47) is lodged in a notch sited on the posterodorsal corner of the posterior ceratohyal.
The urohyal ( Fig. 5 View FIGURE 5 ) is a long shafted bone with low and capitate anterior end and a compressed and leaflike posterior one.
Opercular bones. The opercular series is complete ( Fig. 2 View FIGURE 2 , 3 View FIGURE 3 , and 5). The preopercle ( Fig. 2 View FIGURE 2 and 3 View FIGURE 3 ) is well developed, with subequal dorsal and ventral limbs. The preopercular sensory canal extends along the bone near the anterior margin in a conspicuous bony tube, which ends dorsally in a neurodermal spike. Secondary tubules derived from the main canal are not clearly distinguished on the surface.
The subopercle ( Fig. 2 View FIGURE 2 ) is a falcate bone; its depth is approximately one-third of the opercle depth. This bone shows an almost straight anterior border and a prominent finger-like anterodorsal process.
The interopercle ( Fig. 2 View FIGURE 2 ) is a long triangular bone partially hidden by the ventral margin of the preopercle in all its extension.
The opercle ( Fig. 2 View FIGURE 2 and 5 View FIGURE 5 ) is a large and ovoid bone, of about twice deeper than long. It shows a strong vertical crest along the anterior margin and a deep notch (incisure) on anterodorsal corner like that found in many extant clupeoid fishes (see Grande, 1985).
Vertebral column, pleural ribs, supraneurals, intermuscular bones and abdominal scutes. The body was very compressed in life, judging by the fact that all specimens examined laterally preserved. There are 38 vertebrae (in contrast with 40 pointed out by Woodward, 1939), 23 abdominal and 15 caudal, including the urostyle. The vertebral column is almost straight ( Figs. 6 View FIGURE 6 , 7 View FIGURE 7 ), except for its anteriormost region, where it is clearly slightly arched.
Each vertebral centrum is structurally similar to an elongate spool with a well-developed median constriction. A thin sheet of autocentrum covers a thicker sheet of chordacentrum leaving a large gape corresponding to the notochordal canal.
The neural spines ( Fig. 6 View FIGURE 6 ) are bifid in the abdominal region, becoming medially co-ossified in a unique piece along most of caudal region. The caudal neural spines are as long and slender as haemal spines. Each parapophyses is moderately developed and bears the large proximal end of the pleural rib. The pleural ribs are very slender and slightly arched. They contact the anterodorsal margin of the sharp abdominal scutes and almost reach the ventral midline.
There are 11 prepelvic and eight postpelvic abdominal scutes. All scutes are triangular in shape and keeled ventrally as in generalized clupeid fishes. Posteriorly, the keel presents a short and weak spine.
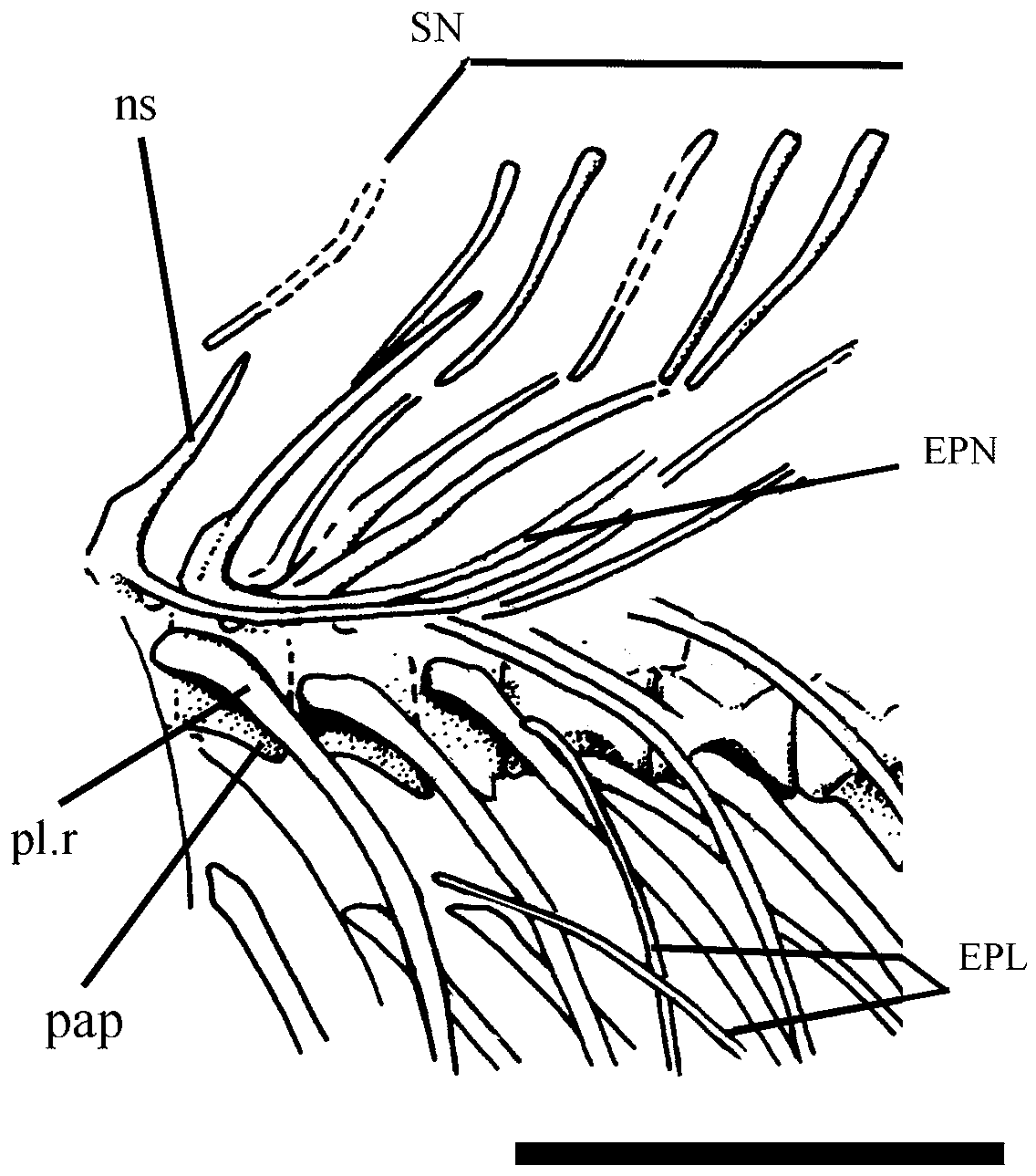
There are eight slender and curved supraneural bones positioned between the occiput and the origin of the dorsal fin. They lack anterior and posterior laminae as well as expanded distal edge.
There is a complex framework of intermuscular bones. The epineurals are thin, long, and very arched along the abdominal region. All are co-ossified to the base of neural arches.
The epipleurals are observed in the abdominal region spanning an extension from vertebrae 15 (or 16) to 35. They are long and arched and laterally attached on the pleural ribs. The intermuscular epicentral bones were not unambiguously distinguished in the specimens examined.
Detached epimeral and hypomeral intermuscular bones are very thin and hypsiloid and distributed along most of caudal region.
Paired girdles and fins. Most of pectoral girdle ( Fig. 2 View FIGURE 2 ) is preserved in specimens examined. The posttemporal ( Fig. 2 View FIGURE 2 ) is a large and drop-like bone in lateral view. The main body of the bone is rounded and scaly; it produces a long and sharp anterodorsal process for meeting epioccipital. The lateral line sensory canal extends along the main body of the bone through a short bony tube.
The supracleithrum ( Fig. 2 View FIGURE 2 ) is a spatulate bone with rounded dorsal border; it tapers downwards for meeting the spine-like process of cleithrum. There is no post-supracleithrum.
The cleithrum ( Fig. 2 View FIGURE 2 ) is a deep and narrow bone. The ventral arm is arched and projects forwards. Medially, its anterior end produces a reduced symphyseal lamina. The dorsal arm of cleithrum is long, slender, and almost straight. It is as high as the opercle and shows a shallow vertical groove on the posterior border in which the long and narrow dorsal post-cleithrum is lodged. This bone meets the middle post-cleithrum ventrally. It is a scaly and falcate bone that covers the anterior half of a long and rod-like ventral postcleithrum.
The coracoid ( Fig. 2 View FIGURE 2 ) is an expanded laminar plate in the form of an inverted “L”. The well-developed anteroventral process extends forwards to meet the cleithrum and leaving a little fenestra between them. There is a well-ossifed and oblique lateral ridge for insertion of the muscle adductor profundus.
The scapula ( Fig. 2 View FIGURE 2 ) is a very reduced trapezoidal bone showing a large scapular foramen in the middle point. Posteriorly, it shows a promiment articular facet for the uppermost fin-ray of the pectoral fin and radial associated. Dorsally, scapula produces the so-called laminate scapular process for lodging modified axilar scales and allowing insertion of the pectoral fin.
The pectoral fin is moderate-sized and shows 14 fin-rays, all segmented and distally ramified. Although badly preserved in most of specimens examined, in the site of insertion of the pectoral fin there are apparently two associated sets of pectoral radials. Four proximal radials are short and club-like whereas the distal ones are nodular.
The pelvic fin is inserted at the middle point between origin of pectoral fin and flexure region of tail (at the level of posterior end of hypurals).
There is a fin-ray segmented but non-ramified distally and six segmented and ramified. Similar to advanced clupeoids, there is no pelvic splint in association to the fin. The design of the pelvic shield obeys the structural pattern found in most of clupeoids, with long and sharp ascending branch and large base (spiniform type). The pelvic bone is often hidden by abdominal scutes so that a reliable description is not possible.
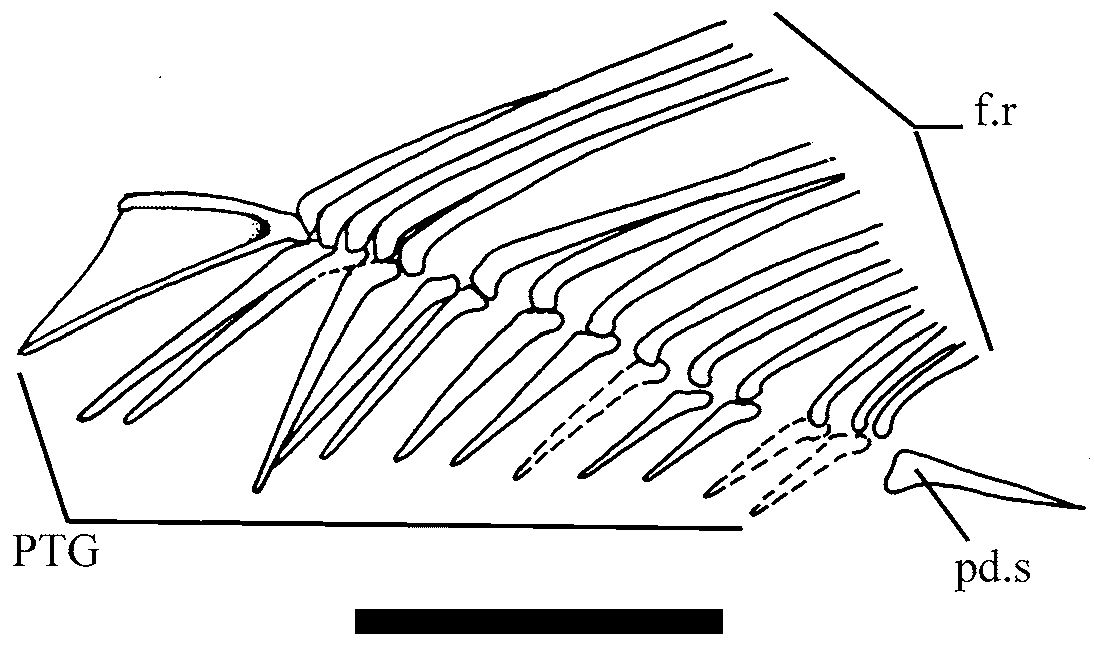
Dorsal and anal fins and endoskeletal support. The dorsal fin is short-based and relatively low ( Fig.7 View FIGURE 7 ). It origins at the level of the vertical line passing between 10 and 11 abdominal vertebrae. There is one nonramified ray followed by 13 to 14 distally ramified fin rays. The set is supported by a row of 14 long and slender proximal pterygiophores. The first pterygiophore is the largest; it shows a continuous keel supported by two well-ossified bars forming a sharp angle in the confluence of them. The last pterygiophore forms a relatively long post-dorsal strut.
The anal fin ( Fig. 8 View FIGURE 8 ) is composed of 15 segmented and distally ramified fin rays supported by 14 straight and slender pterygiophores. The two anteriormost pterygiophores are longer than the rest. This fin originates at the level of preural vertebrae 15 to 16.
Caudal endoskeleton and fin. The caudal fin is deeply forked, with upper and lower lobes subequal in size. A set composed of one non-ramified principal ray plus nine ramified rays forms the upper lobe of the caudal fin. The uppermost fin rays partially cover fifth and sixth hypurals. There is one non-ramified plus eight ramified principal fin rays below the diastema. At least four precurrent rays are visible on dorsal and ventral border of the caudal fin.
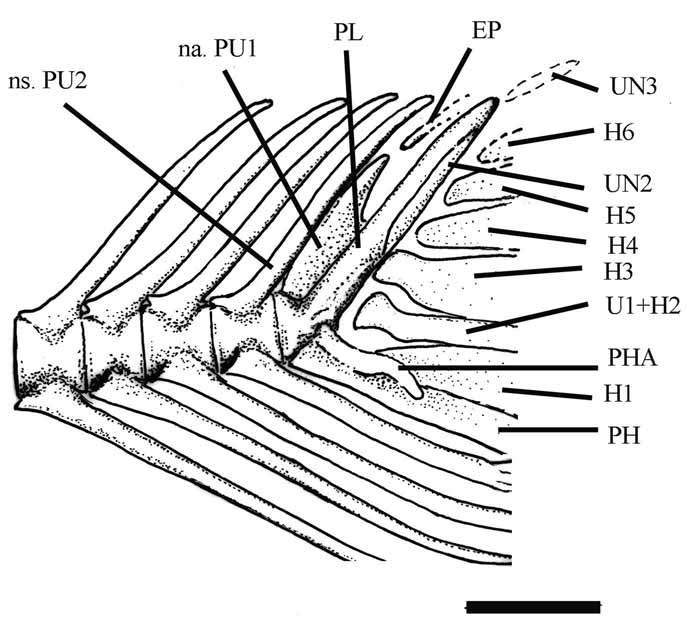
In most of specimens examined the caudal endoskeleton is relatively well preserved ( Fig. 9 View FIGURE 9 ). The neural spine and, mainly, the neural arch of first preural centrum produce an elongate leaf-like structure (short in species of † Knightia ) on which lies the tip of a long and slender epural.
The parhypural ( Fig. 9 View FIGURE 9 ) is slender and apparently not co-ossified to first preural centrum; it has a long and curved parhypurapophysis.
There are six hypurals ( Fig. 9 View FIGURE 9 ). The first hypural is a large and long triangular plate spaced from the first preural centrum and urostyle. It is followed by a long, very slender and low second hypural. Proximally, it is apparently fused to first ural centrum as indicated by the dilated proximal end resembling the condition found in Dorosoma cepedianum (see Grande, 1985, p. 347, Fig. 51).Yet the second ural centrum is conical and very reduced (visible in DGM 35). The remaining hypural bones are low and triangular, and gradually decrease in size towards end of the upper lobe following the slope of the main axis of vertebral column. It is not possible to determine presence (as derively found in pristigasteroideans) or not of interlobar notch in the third hypural.
There are three uroneurals ( Fig. 9 View FIGURE 9 ) from which the first one is longer and stronger than other of the set. It is fused to the first preural centrum forming a pleurostyle (sensu Monod, 1968) which is almost parallel to second preural neural spine. Other uroneural bones are small and fusiform. They are in a row over the second ural centrum. The second uroneural is placed over the upper third of pleurostyle and reaches the third uroneural. Even, dorsal and ventral borders of the caudal peduncle are guarnished of dorsal and ventral scutes, respectively. They are short and lenticular.
The body is covered of deep and ovoid cycloid scales showing vertically arranged circuli. They span an extension from the face to the base of tail. As noted by Woodward (1939), they are obscure, poorly preserved. There is no positive evidence of perforated lateral line scales.




| DGM |
Divisao de Geologia c Mineralogia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paleopiquitinga brasiliensis
| De, Francisco J. 2010 |
Knightia brasiliensis
| Santos 1946: 1 |
| Woodward 1939: 450 |