
Metania madagascariensis Manconi and Pronzato
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3918.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:03928490-DD7C-4DB2-9470-C0417D05AF27 |
|
DOI |
https://doi.org/10.5281/zenodo.6102613 |
|
persistent identifier |
https://treatment.plazi.org/id/03F79750-B506-FFA2-FF6D-FCEFFCFBFEAB |
|
treatment provided by |
Plazi |
|
scientific name |
Metania madagascariensis Manconi and Pronzato |
| status |
sp. nov. |
Metania madagascariensis Manconi and Pronzato sp. nov.
( Figs 1–8 View FIGURE 1 View FIGURE 2 View FIGURE 3. A View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 )
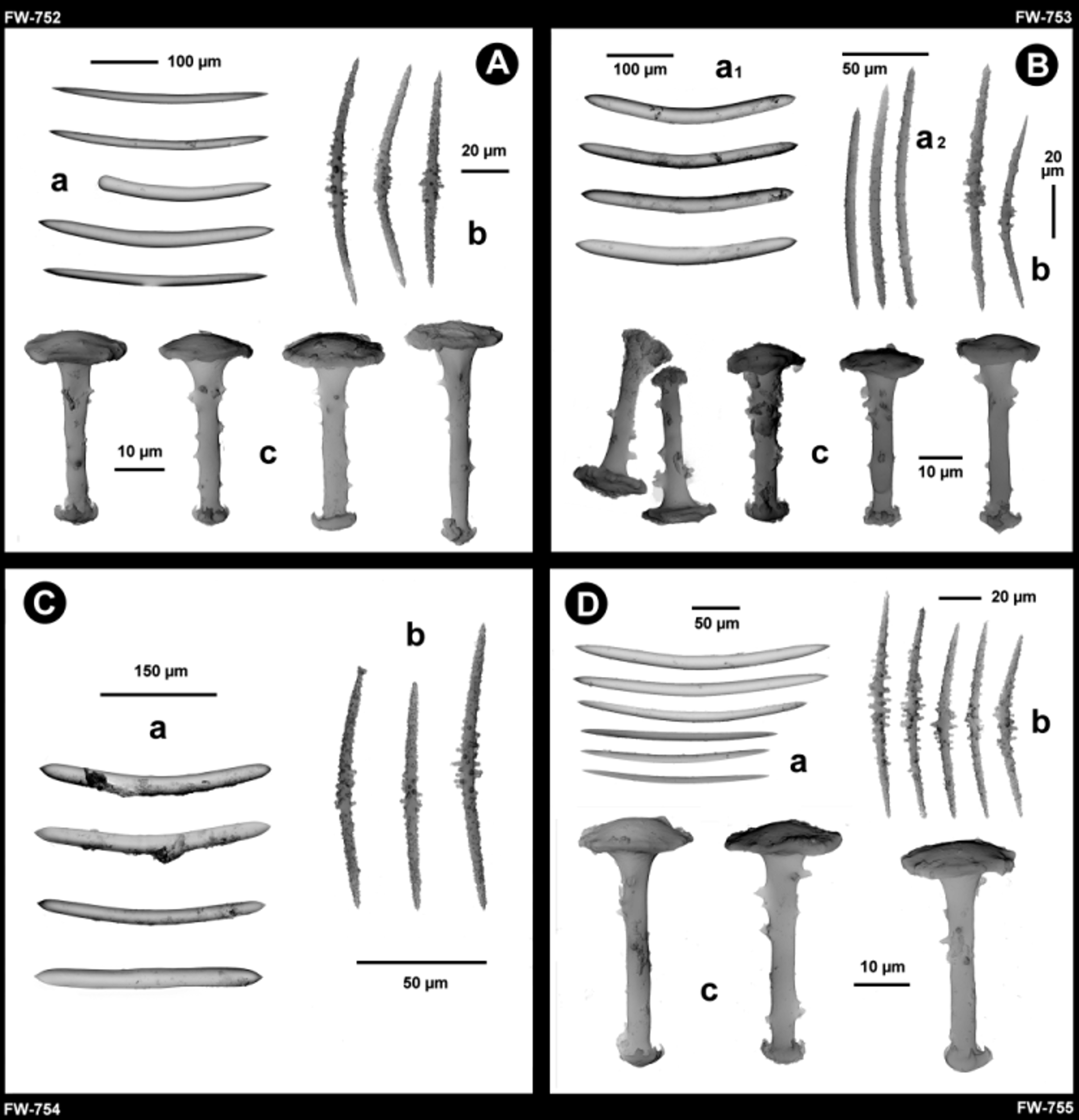
Holotype. MSNG 57788 (schizotype DISTAV-FW 752), MADAGASCAR, Haute Matsiatra Region, River Matsiatra, 21°25'36.1092"S, 47°9' 23.7054"E, tributary of the higher River Mangoky basin, Central High Plateau, between Fianarantsoa North and Ambalakely, under a bridge along the highway Route Nationale n° 7, station MAD 28, R. Manconi leg., 25/ix/2011. P aratype. MSNG 57789 (schizoparatype DISTAV-FW 753), same data as holotype. Other material. DISTAV-FW 754, DISTAV-FW 755, same data as holotype.
Comparative material. Tubella rhodesiana Lake Ishiba Ngandu (ex-Lake Young, Rhodesia), C.K. Ricardo leg. 13.viii.1936, 5 m depth, (holotype BMNH 1938.2.25.1, schizotype DISTAV-FW 432); ibid., 13.i.1937, 2.5–3 m depth (paratypes BMNH 1938.2.25.5, BMNH 1938.2.25.7, schizotypes DISTAV-FW 609, DISTAV-FW 610 respectively); ibid. (topotypes BMNH 1938.2.25.2, BMNH 1938.2.25.3, schizotypes DISTAV-FW 632, DISTAV- FW 611 respectively). Metania innominata L. Ishiba Ngandu, C.K. Ricardo leg., 9.v.1936, (Holotype BMNH 1938.2.25.8, schizotype POR- FW 608). Spongilla brieni Congo, Lake Upemba, P. Brien leg., viii.1937, Burton 1938 det. (holotype MRAC 201, schizotype DISTAV-FW 490). Metania brieni Lake Upemba , Congo ( BMNH 1938.2.1.2–13. IIIA, fragment DISTAV-FW 614). Metania lissostrongyla Lukula River ( BMNH 1938.2.3.3, fragment DISTAV-FW 633); Leopoldville, H. Schouteden leg. ( BMNH 1938.2.3.7, fragment DISTAV-FW 612); Metania schoutedeni E. Dartevelle leg. ( MRAC 1006, fragment DISTAV-FW 372); Metania schoutedeni Leopoldville, 1937 , A. Tinant leg. (paratype MRAC 142, schizotype DISTAV-FW 373); Metania vanryni Kinshasa, E. Dartevelle leg. (holotype MRAC 1008, schizotype DISTAV-FW 481); Metania vesparia Angola, coll. Mission A. Powell-Cotton ( BMNH 1938.5.10.1, fragment DISTAV-FW 430); E. Dartevelle leg. ( BMNH 34.9.12.1, fragment DISTAV-FW 443). Parametania godeauxi , Katanga, Luapula River at the mouth of the Kafubu River, J. Godeaux leg., viii.1963, (schizotype MRAC 1312, DISTAV- FW 480 slides and stubs).
Etymology. The specific epithet madagascariensis is derived from the name of Madagascar this being the only species of the genus Metania reported from the island.
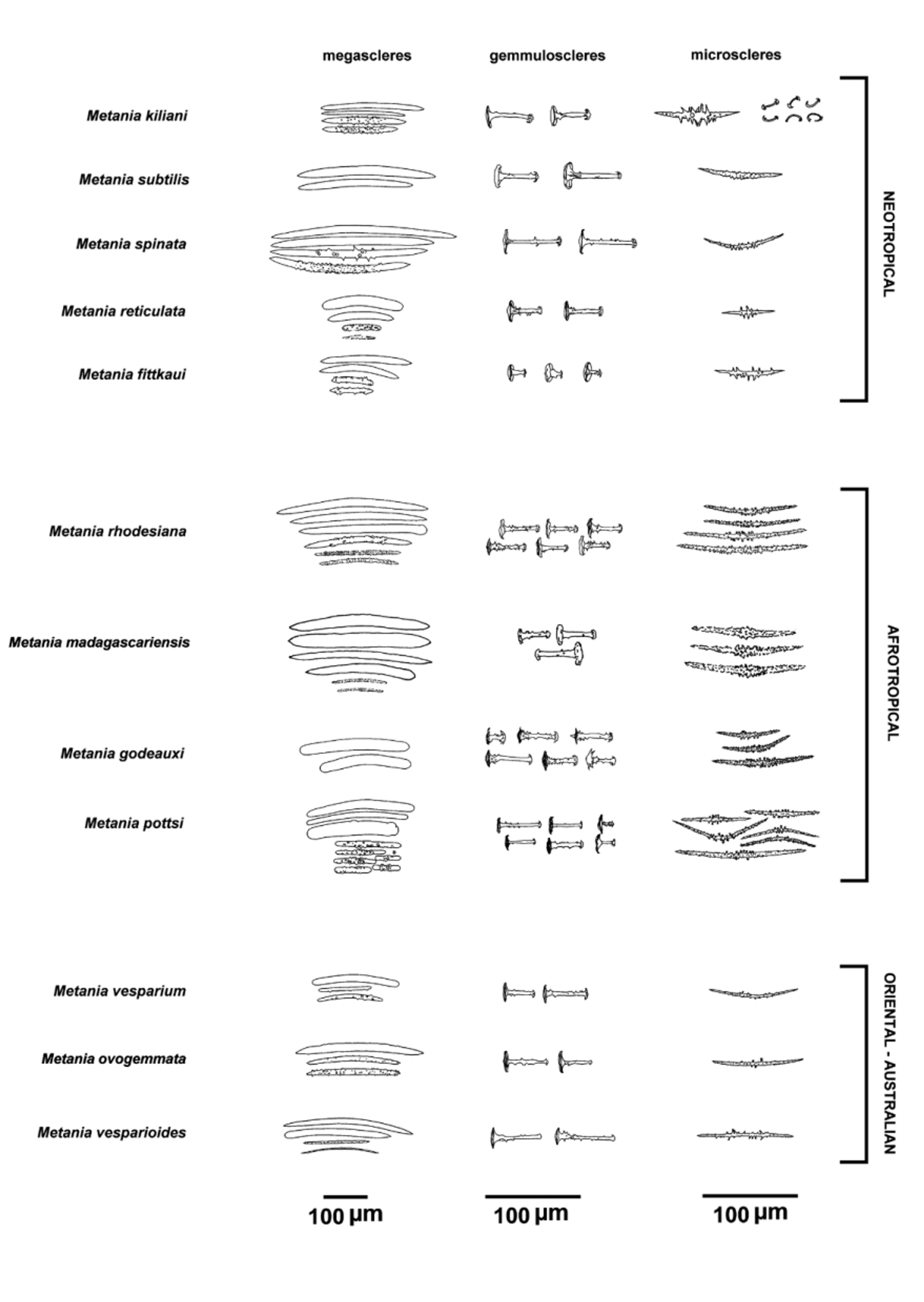
Diagnosis. Smooth oxeas as main megascleres (α), slender and short oxeas as β megascleres, gemmular cage of α megascleres present to absent.
Description. Growth form encrusting to cushion-like. Consistency hard and fragile both in vivo and dry condition. Colour whitish to dark brown both in vivo and dry condition. Surfac e conulose with acute (1–1.5 mm in height), dense conules (0.5–1.5 mm apart) with apical tufts of oxeas to support more or less rounded areas of the translucid dermal membrane. Oscules scattered, circular, small (0.5–2 mm in diameter). Inhalant apertures scattered in the dermal membrane. Ectosomal skeleton densely armed by microscleres tangentially arranged in the dermal membrane, supported by tips of ascending pauci-spicular fibres, arranged in a superficial network of irregular polygonal, mono- to pauci-spicular meshes (300–600 µm in diameter). Subectosomal skeleton with short pauci- to multi-spicular ascending fibres up to the ectosome to support conules. Choanosomal skeleton irregularly alveolate network of pauci- to multi-spicular (stout smooth oxeas) polygonal meshes (ca. 0.5 mm in diameter) supported in some areas by extremely dense assemblages of microscleres. Spongin scanty, except for the gemmular theca and the basal spongin plate. Basal spongin plate well developed, tangentially armed by megascleres as stout, smooth oxeas and by microscleres as spiny oxeas. Megascleres oxeas of two types smooth and spiny, abruptly pointed to acerate. Type 1 (α and β megascleres) abundant smooth oxeas (255–387 x 10–30 µm), stout, straight to notably bent, with acute, abruptly pointed tips; rare styles also present, probably freak. Type 2 (β megascleres) less abundant acanthoxeas (120–178 x 5–8 µm) entirely spiny by scattered small spines of same dimension; spines in the central portion of the shaft with a few microspines (2–5) in rosettes, and simple spines sometimes more dense towards the abruptly pointed tips (more abundant in paratype MSNG 57789). Rare, slender, smooth oxeas (155–345 x 5–18 µm) slightly bent, with acerate tips also present. Very thin, smooth oxeas (53–103 µm in length) probably belong to larvae. Microscleres acanthoxeas (55–128 µm in length) ranging from 8–12 µm in thickness with tubercles/spines, to 3–5 µm in thickness not considering spines height, bent to straight with sharp, abruptly pointed tips, and less frequently with blunt tips; entirely spiny by dense, small spines, and with in the central portion of the shaft variably long, stout, straight, less dense tubercles/spines bearing rosettes of dense microspines at their tips. Gemmular cage absent or as an assemblage of stout, smooth oxeas (megascleres); microscleres of the skeleton frequently present around gemmules. Gemmules singly scattered in the skeletal meshes, very rarely grouped, subspherical, large (450–500 µm in diameter), sometimes collapsed in dry condition. Foramen (50–70 µm in diameter) well evident with a simple collar surrounded sometimes by an undulated circular lamina to with a short vase-like tubule. Gemmular theca trilayered (ca. 50 µm in thickness). Outer layer of compact spongin variably developed with emerging distal knob-like tips (pseudorotule) of radially embedded gemmuloscleres. Pneumatic layer fibrous, not chambered, as a network of thin spongin fibres/trabecules arranged as more or less rounded meshes with a variable diameter ranging from large at the distal portion of the theca to smaller at the proximal one towards the inner layer. Inner layer of compact sublayered spongin strictly in contact with proximal rotules of partly overlapping gemmuloscleres. Gemmuloscleres arranged in a single layer with proximal rotule strictly adhering to the inner layer and distal pseudorotule emerging from the theca, tubelliform/ trumpet-like (32–55 µm in length) with straight shaft (4–5 µm in thickness), variably spiny (up to 10 spines/ tubercles with microspines) to less frequently smooth, with proximal true rotules (18–23 µm in diameter) with irregular blunt margins, and distal, knob-like pseudorotules (5–10 µm in diameter) sometime umbonate, with a few hooks at the margins.
Habitat. M. madagascariensis sp. nov. inhabits the Central High Plateau at an altitude ca. 1000 m asl under a bridge crossing the River Matsiatra, within the higher River Mangoky hydrographic basin tributary of the Mozambico Canal. The new species matches the tropical rain forests range, although forests are now relictual in the area. The survey of ca. 50 pebbles performed in the watercourse (ca. 10 m wide) at the end of the dry season (late September) resulted in the discovery of scattered but not abundant sponges at 5–70 cm of depth under submerged boulders and pebbles in brown, silty shallow water along the seasonally flooded riverside. Active sponges, both young and small to old and large bear gemmules in late September. Specimens of the new species were associated, also on the same substratum, with almost two other presently unidentified species of Spongillina (Manconi et al. in prep.). Bryozoans and aquatic insects (larvae and adults) were also present.
Geographic distribution. M. madagascariensis sp. nov. belongs to the Afrotropical group of Metania and is known until now from the type locality.
Remarks. We consider the differences in the microtraits of the spicular complement displayed by the studied specimens from Madagascar as intraspecific phenotypic variability. Diverging diagnostic traits of the new species vs. all the other species of Metania are clearly shown in Fig. 2 View FIGURE 2 .






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.