
Mystacides rivularis, Kuhara & Nozaki & Zhang & Zhou, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5306.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:F7C3E432-3FCF-4A3A-8972-C4012346D338 |
|
DOI |
https://doi.org/10.5281/zenodo.8061284 |
|
persistent identifier |
https://treatment.plazi.org/id/03F7F326-302D-FFA0-DDCB-0E8905C55E6B |
|
treatment provided by |
Plazi |
|
scientific name |
Mystacides rivularis |
| status |
sp. nov. |
Mystacides rivularis n. sp.
Figs 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3
Mystacides sp. : Nozaki & Tanida 2007: 252.
Diagnosis. This species can be distinguished from M. azureus , which is occasionally collected sympatrically with M. rivularis n. sp., by shape of the male segment IX that is strongly extended posteroventrad. Among species of the M. azureus Species Group, this character state is similar to that of M. schmidi Morse & Yang 2002 from Sri Lanka, but distinguishable from the latter by the long sigmoid spine of segment X. The female of this species is characterized by the manta-ray-shaped plate of the spermathecal sclerite.
Adult. Forewing length: male 6.4–7.6 mm (mean = 7.2 mm, n = 10); female 6.8–7.6 mm (mean = 7.3 mm, n = 9). Vertex, thorax, and wings in ethanol brownish black. General morphology typical for the genus. Relative eye size smaller than that in M. azureus for both male and female ( Fig. 1 View FIGURE 1 ).
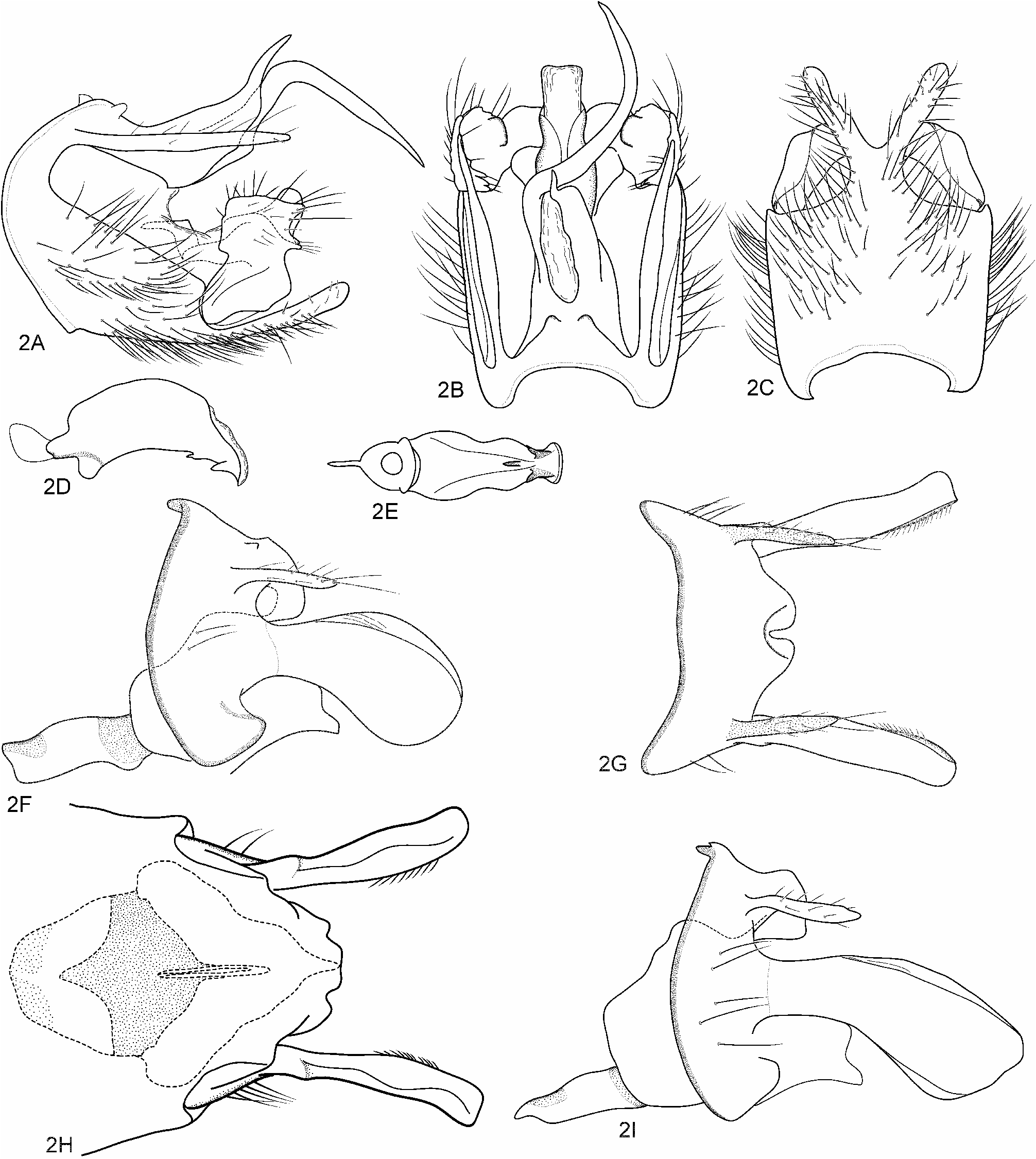
Male genitalia ( Figs 2A–2E View FIGURE 2 ). Segment IX 1/5 as long dorsally as tall laterally, 1 1/5 as long ventrally as tall laterally; in ventral view apicoventral process of sternum 3/4 as long as rest of sternum, with stout base and slender V-shaped lateral arms. Preanal appendages very long and slender. Tergum X antisymmetrical, produced into two sinuate spines: one spine sigmoid in dorsal aspect, long, extending far beyond apical process of sternum IX; other spine weakly sinuate and shorter than former. Inferior appendages each with dorsomesal lobe oriented vertically, somewhat longer than mid-width in lateral view, its anterior and posterior margins subparallel; posterior margin with three caudal projections: upper caudal projection with acute apex; middle caudal projection narrow and often longest among three projections, with acute apex; lower caudal projection with round apex. Phallus without evident paramere spines or phallicata, with ventromedian spine directed anteroventrad at one third distance from apex and pair of sharp, triangular flanges subapically, apicoventral lip of phallobase constricted subapically in ventral aspect.
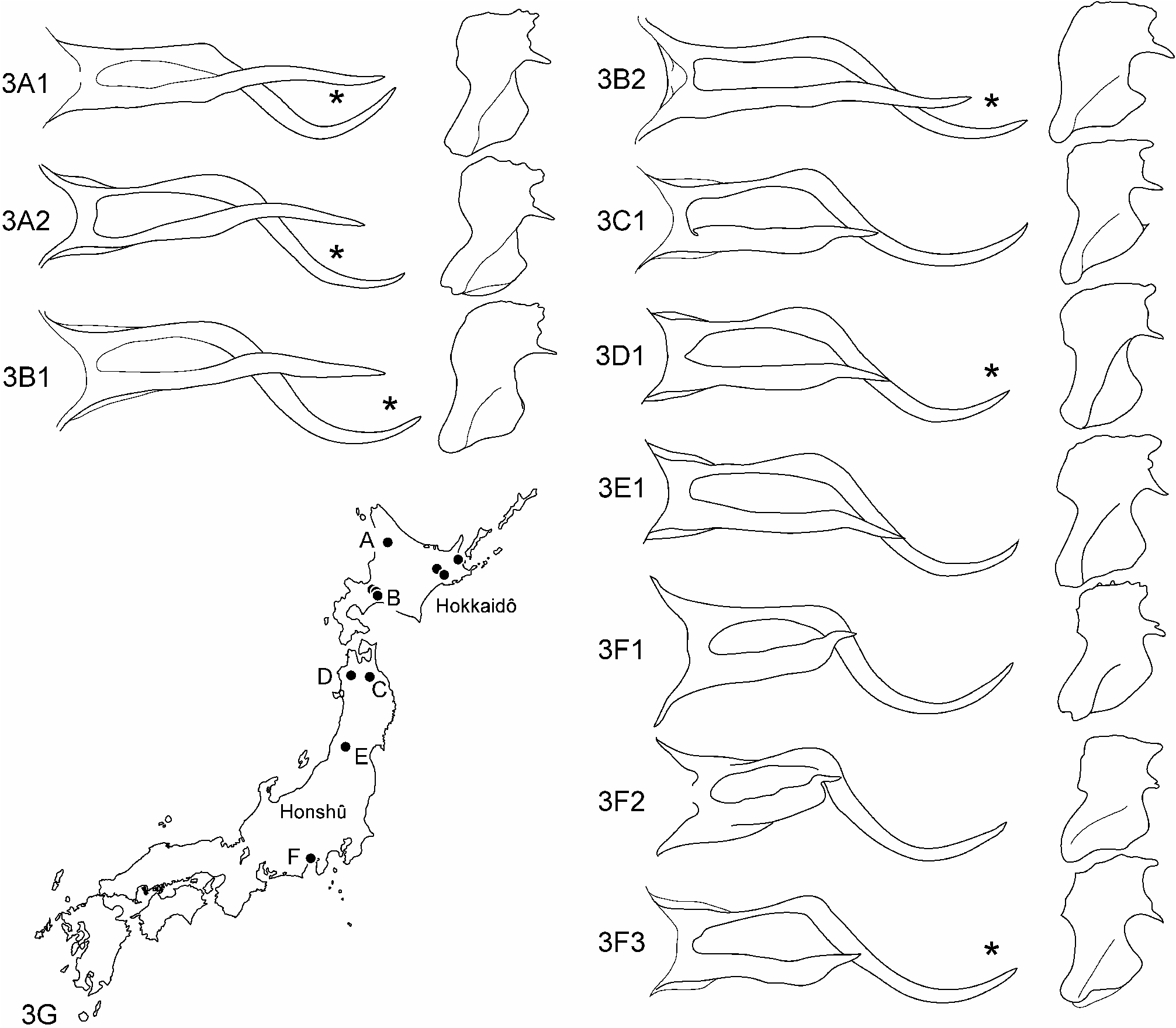
Variation ( Figs 3A1–3F3 View FIGURE 3 View FIGURE 1 View FIGURE 2 ): The shapes of tergum X and inferior appendages are variable geographically. The shorter process of tergum X is about half the length of the longer process in individuals from central Honshû ( Figs 3F1–3F3 View FIGURE 3 View FIGURE 1 View FIGURE 2 ), but the former is prolonged in those from northern Honshû ( Figs 3C1–3E1 View FIGURE 3 View FIGURE 1 View FIGURE 2 ) and almost reaches the tip of the latter in those from Hokkaidô ( Figs 3A1–3B2 View FIGURE 3 View FIGURE 1 View FIGURE 2 ). The middle projection of each inferior appendage tends to be more strongly developed in individuals from Hokkaidô and northern Honshû ( Figs 3A1–3E1 View FIGURE 3 View FIGURE 1 View FIGURE 2 ) than those from central Honshû ( Figs 3F View FIGURE 3 1 View FIGURE 1 , 3F View FIGURE 3 2 View FIGURE 2 ) with an exception ( Fig. 3F3 View FIGURE 3 ).
Female genitalia ( Figs 2F–2I View FIGURE 2 ). Segment IX short in lateral view. Segment X shorter than preanal appendages, narrowly incised ventromedially in dorsal view. Preanal appendages slender, straight, and setose. Lamellae very long, constricted basally, shape in lateral aspect variable geographically; dorsal margin inflated; apicodorsal corner protruded outward; outer surface slightly concave. Gonopod plates triangular in ventral aspect with pair of posterolateral processes short, rounded and apex extending far beyond apex of segment X. Spermathecal sclerite with pigmented manta-ray-shaped plate, anterior and posterior margins extended anterad and posterad mesally and acute.
Variation: Lamellae in lateral aspect are each subelliptical and rounded apically in individuals from the type locality Kakida-gawa, central Honshû ( Fig. 2F View FIGURE 2 ), but more nearly parallel-sided, gradually heightened caudally, and obliquely truncate apically in those from Hokkaidô ( Fig. 2I View FIGURE 2 ) .
Holotype: ³, Kakida-gawa , Shimizu-chô, Shizuoka, Honshû, Japan, 35.1031°N, 138.9028°E, alt. 13 m, 27.iv.2002, TN. GoogleMaps
Paratypes: 3³, same data as holotype; 5³, type locality, 22–23.xi.2002, NK, 2³ (pinned), type locality, 5.iv.2006, TN.
Other specimens examined. HOKKAIDÔ: Nemuro : 1♀, Shibetsu-chô, Shibetsu-shitsugen , 12.viii.1996, K. Kuribayashi, 1³ 1♀, same location, 21.viii.2013, M. Nakatani ; 1♀, same location, 2–3.viii.2013, M. Nakatani; 1³ 1♀, same location, 3.ix.2013, M. Nakatani. Kushiro: 1³, Kushiro-shi, Akan-chô, Ibeshibetsu-gawa R ., 6.viii.1990, NK; 1³, same location, 6.viii.1990, NK; 3³ 1♀, Shibecha-chô, Gojikkoku, Shirarutoroetoro-gawa R ., 16.vii.2009, TI ; 2³ 7♀, same location, 28.vii.2012, TI ; 1³ 2♀, Shibecha-chô, Kayanuma, Shirarutoroetoro-gawa R ., 16.vii.2009, TI ; Kamikawa: 1³, Horokanai-chô, small tributary of Shumarinai-gawa R ., 10.viii.1999, TI & A. Ohkawa; 1³, Horokanai-chô, small tributary of Shumarinai-gawa R ., 7.vii.2007, NK. Ishikari: 1³, Chitose-shi, Bibi, Bibi-gawa R ., 5–26.ix.1993, TI ; 1³ 1♀, same location, 4.viii.2007, TI ; 1³ 3♀, Eniwa-shi, Izari-gawa R ., Eniwa-ôhashi , 11.vii.1999, TI ; 1³, same location, 22.vii.1999, TI ; 1³, same location, 17.vii.2015, TI ; 1♀, Sapporo-shi, Hitsujigaoka, 24–31.vii.2009, K. Konishi; 1³, Sapporo-shi, Nopporo-shinrin-kôen, Osawa-guchi , 17.vi.2002, M. Sakurai. Iburi: 3³ 5♀, Tomakomai-shi, Misawa, Bibi-gawa R ., 29.vii.1989, NK; 5³ 1♀, same location, 17.vii.1990, NK ; 1♀, same location, 16.viii.1990, NK ; 4♀, same location, 18.viii.1991, TI ; 2³, same location, 12.ix.1993, TI ; 1³, same location, 2.viii.1998, NK; 1³, same location, 20.viii.2007, TI & A. Ohkawa; 5³ 8♀, same location, 31.vii.2009, NK; 2³, Tomakomai-shi, Misawa , small stream, 1.viii.1992, NK; 1³, same location, 15.viii.1992, NK; 5³ 2♀, Tomakomai-shi, Uenae, Bibi-gawa R ., 22.vii.2001, TI & A. Ohkawa; 1³, same location, 25.vii.2001, TI ; 1³, same location, 29.vii.2001, TI ; 1³, same location, 30.vii.2001, TI ; 3³ 2♀, same location, 23.viii.2007, TI ; 2³ 3♀, same location, 16.vii.2008, TI . HONSHÛ: Aomori: 1³, Nishimeya-mura, Anmon-gawa R ., alt. 240 m, 16.ix.2010, TI ; 1³, Takko-machi, Natsuzaka, Kumahara-gawa R ., 18.viii.1996, Suzuki. Yamagata: 3³, Nishikawa-chô, Shizu , Buna-no-izumi, 11.ix.2003, TI . Shizuoka: 1♀, type locality, 11.iii.1984, reared and emerged on 7.iv.1984, TN; 3³ 1♀, same data as holotype; 1³ GoogleMaps 1♀, type locality, 27.iv.2002, TN; 1♀, type locality, 31.viii.2002, TN; 2³ 1♀, type locality, 14.xi.2002, TN; 6³ 1♀, type locality, 22–23.xi.2002, NK.
Etymology. The specific epithet (Latin adjective, rivularis = of a brook or small stream) refers to the habitat of this species.
Distribution. Japan (Hokkaidô, eastern Honshû).
Habitat. Adults are often found beside slowly flowing streams. They are sometimes collected with M. azureus sympatrically, but the habitat preference of the new species is narrower than that of the latter.
| R |
Departamento de Geologia, Universidad de Chile |
| TI |
Herbarium of the Department of Botany, University of Tokyo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Mystacides rivularis
| Kuhara, Naotoshi, Nozaki, Takao, Zhang, Ao & Zhou, Xin 2023 |
Mystacides sp.
| Nozaki, T. & Tanida, K. 2007: 252 |