
Chiasmocleis shudikarensis Dunn, 1949
|
publication ID |
https://doi.org/ 10.1206/834.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03F8878E-6F46-8422-FD0E-F9E67265F951 |
|
treatment provided by |
Felipe |
|
scientific name |
Chiasmocleis shudikarensis Dunn, 1949 |
| status |
|
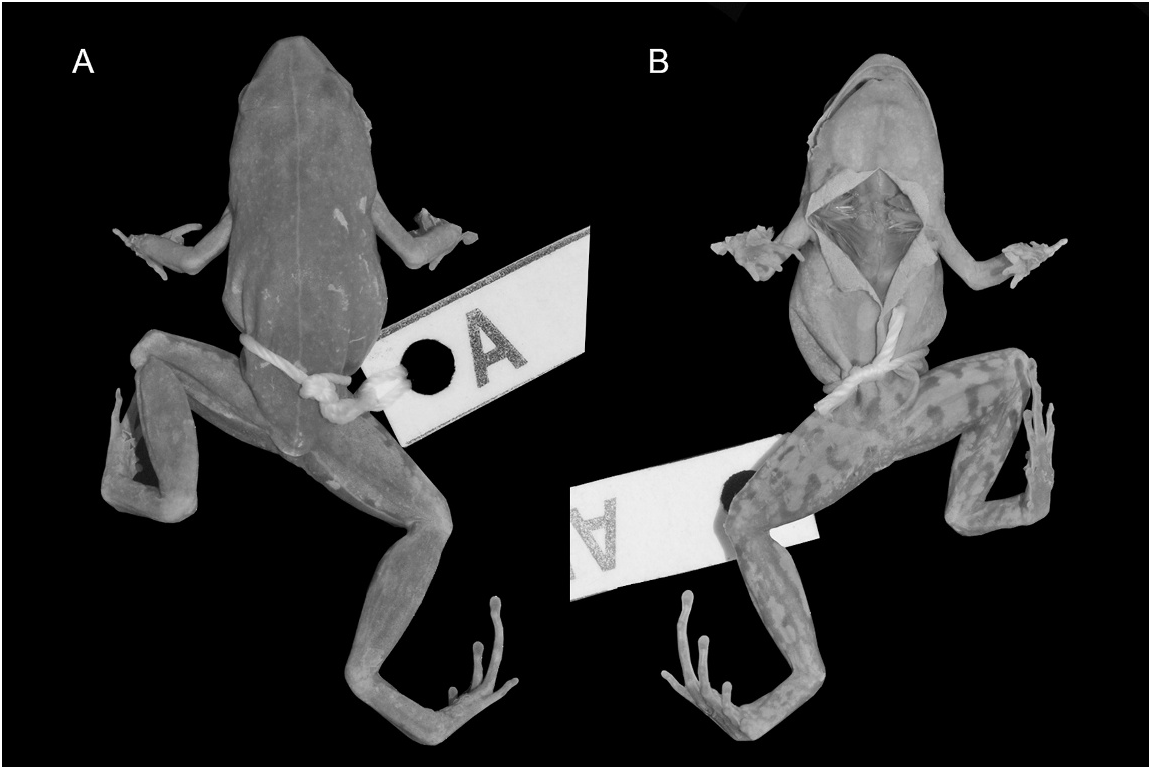
Chiasmocleis shudikarensis Dunn, 1949 View in CoL Figures 33 View Fig , plates 7, 14
HOLOTYPE (fig. 33): AMNH 43674 About AMNH ; adult, probably female specimen. The chest is cut open. Both hands are partially destroyed and so is the dorsal portion of the left foot. Other than that, the specimen is well preserved and some details of color pattern are still clearly visible.
TYPE LOCALITY: Shudikar-Wau, Upper Essequibo River, British Guiana (now the Co-operative Republic of Guyana). No geographic coordinates were given in the original description.
DIAGNOSIS: A medium-sized species for the genus; SVL in males 18.7–25.5 mm (N 5 22); in females 20.9–29.7 mm (N 5 11). Body ovoid, robust; head triangular, snout round- ed in dorsal and lateral views. Four distinctive fingers, all but FI fringed in males, much less fringed in females; fingers not webbed; FI well developed with a distinct well-developed subarticular tubercle present between the proximal phalanges; distinctly visible subarticular tubercles present on all fingers; adpressed FI reaches or extends beyond the subarticular tubercle of FII; adpressed FIV does not reach or barely reaches distal tubercle of FIII; palmar tubercles protuberant, divided; relative finger lengths I,IV,II,III. Five distinctive and well-developed toes present; toes fringed, less distinct in females; toes usually extensively webbed in males (fig. 5C) and only basally webbed in females (fig. 5B); TI with a distinct welldeveloped subarticular tubercle; adpressed TI does not touch or barely touches subarticular tubercle of TII; adpressed TV does not touch or reaches only the middle of subarticular tubercle of TIV; TII–V with terminal discs; relative toe lengths I,II, V,III,IV. Males with many dermal spines on fingers and toes; both sexes may show dermal spines on dorsum, fingers, and toes, much more numerous and developed in males; males with many spines on anterior portion of chin, lacking in females. Femoral line is always present (see pl. 7E).
VARIATION: In almost all populations where large series of both males and females were examined (e.g., FLONA do Amapá, Amapá, and Manaus, Amazonas), marked sexual dimorphism in fringes on hands was observed, with males having conspicuously more developed fringes than females. The fringes in males are usually ornamented with numerous dermal spines. Males from Petit Saut, French Guiana, also show more developed fringes in fingers than females, but the difference is not as conspicuous as in other populations (e.g., Manaus and Amapá).
In life, the dorsum of males is usually reddish or grayish but is commonly marbled in shades of yellow or orange (pl. 7). Dorsum of females is usually more uniformly colored, gray or blackish. Snout follows the same general pattern of the dorsum, or is whitish (pl. 7A, C). A dorsal line may be present, varying in extent (as shown in fig. 10). Hind limbs of males and females usually yellow, orange, or reddish. Forelimbs grayish, yellow, or reddish, being uniform in females and usually marbled (as in the dorsum) in males. Venter color pattern varies in much the same way as it does in C. avilapiresae (illustrated in fig. 20). An inguinal blotch is invariably present, although variable in shape and size.
CALL AND TADPOLE: The advertisement call of Chiasmocleis shudikarensis was described by Zimmerman and Bogart (1988) from Manaus, Amazonas, Brazil; and by Lescure and Marty (2000) from French Guiana. Both calls presented distinct acoustic parameters: Manaus, Brazil —‘‘frequency range’’ 5890–7510 Hz; note duration 10–60 ms; interval between notes 10–30 ms ( Zimmerman and Bogart, 1988); French Guiana —call rate 360 notes/min; dominant frequency 3380– 3750 Hz; note duration 98 ms ( Lescure and Marty, 2000). The difference in dominant frequency is striking and deserves further scrutiny as it suggests the possible that at least two distinct taxa are present.
The tadpole of Chiasmocleis shudikarensis was supposedly illustrated but not described by Hero (1990). The illustrations were later reported to belong to another microhylid, Ctenophryne geayi ( Menin et al., 2011) .
TABLE 10 Uncorrected pairwise distances between 16S sequences of Chiasmocleis shudikarensis
Tadpoles of C. shudikarensis were recently described in detail by Menin et al. (2011).
REMARKS: Ávila-Pires et al. (2010) referred to a presumably undescribed species from the portion of the state of Pará north of the Amazon River. P.L.V.P. was present on a fieldtrip where two of the specimens were collected (Estação Ecológica Grão Pará: CN2112, CN2185—MPEG 28327, MPEG 28328, respectively; see pl. 7A) and we have carefully examined some additional specimens from the series mentioned by Ávila- Pires et al. (2010) at the MPEG collection. We disagree with Ávila-Pires et al. (2010) and consider the populations to be unambiguously assignable to either C. shudikarensis or C. hudsoni (see appendix 1 for identifications). The C. shudikarensis specimens from ‘‘Calha Norte’’ do have a smaller SVL than specimens from French Guiana and Amazonas, but they agree with the variation we observed in larger samples of C. shudikarensis , and also agree with the original description of the species ( Dunn, 1949) and the general morphology with the holotype (AMNH 43674).
Despite distinct advertisement call parameters, we have decided not to name an additional species of Chiasmocleis allied to C. shudikarensis at this time for the following reasons: we have examined specimens of the population assigned to the ‘‘Manaus call type’’ (INPA 245, 277–78, 289, 320), but have not examined the vouchers for the ‘‘ French Guiana call type.’’ The specimens from Manaus agree with the original description and with the holotype C. shudikarensis . We have examined a series of specimens from Petit Saut, French Guiana (MPEG 5106– 5107, 5121, 5029–5033, 5060) and the only morphological differences are that male specimens from Sinnamary River show considerably less fringed fingers than those from Manaus, Amazonas, Brazil, and females from French Guiana are slightly more robust. Therefore, the population from French Guiana is also unambiguously assignable to C. shudikarensis . In this particular case, examination of specimens and advertisement calls from the vicinities of the type locality ( Essequibo River) is advisable before nomenclatural actions are taken. Genetic distances between 16S sequences included in our analysis very low and are given in table 10.
DISTRIBUTION (fig. 34): Restricted to the Guiana Shield in Guyana, Surinam (we have not examined any specimens from Surinam), French Guiana, and Brazil, north of Rio Amazonas (Amapá, Amazonas, Pará, and Roraima). Rodrigues et al. (2004b) provided a map that likely overestimates the distribution of the species, extending south of Rio Amazonas (Acre, Mato Grosso, and Rondônia). All of the specimens previously identified as Chiasmocleis shudikarensis in collections from southern Amazonia were identified by us as either C. avilapiresae or C. bassleri .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |