
Clytia gracilis (M. Sars, 1850)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3908.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:D6AD2B49-170B-4D9C-84AA-DBE0FEEAD8BE |
|
DOI |
https://doi.org/10.5281/zenodo.6107190 |
|
persistent identifier |
https://treatment.plazi.org/id/03F887DE-FF7D-FF35-9CD6-0C0DD41CF80A |
|
treatment provided by |
Plazi |
|
scientific name |
Clytia gracilis (M. Sars, 1850) |
| status |
|
Clytia gracilis (M. Sars, 1850) View in CoL
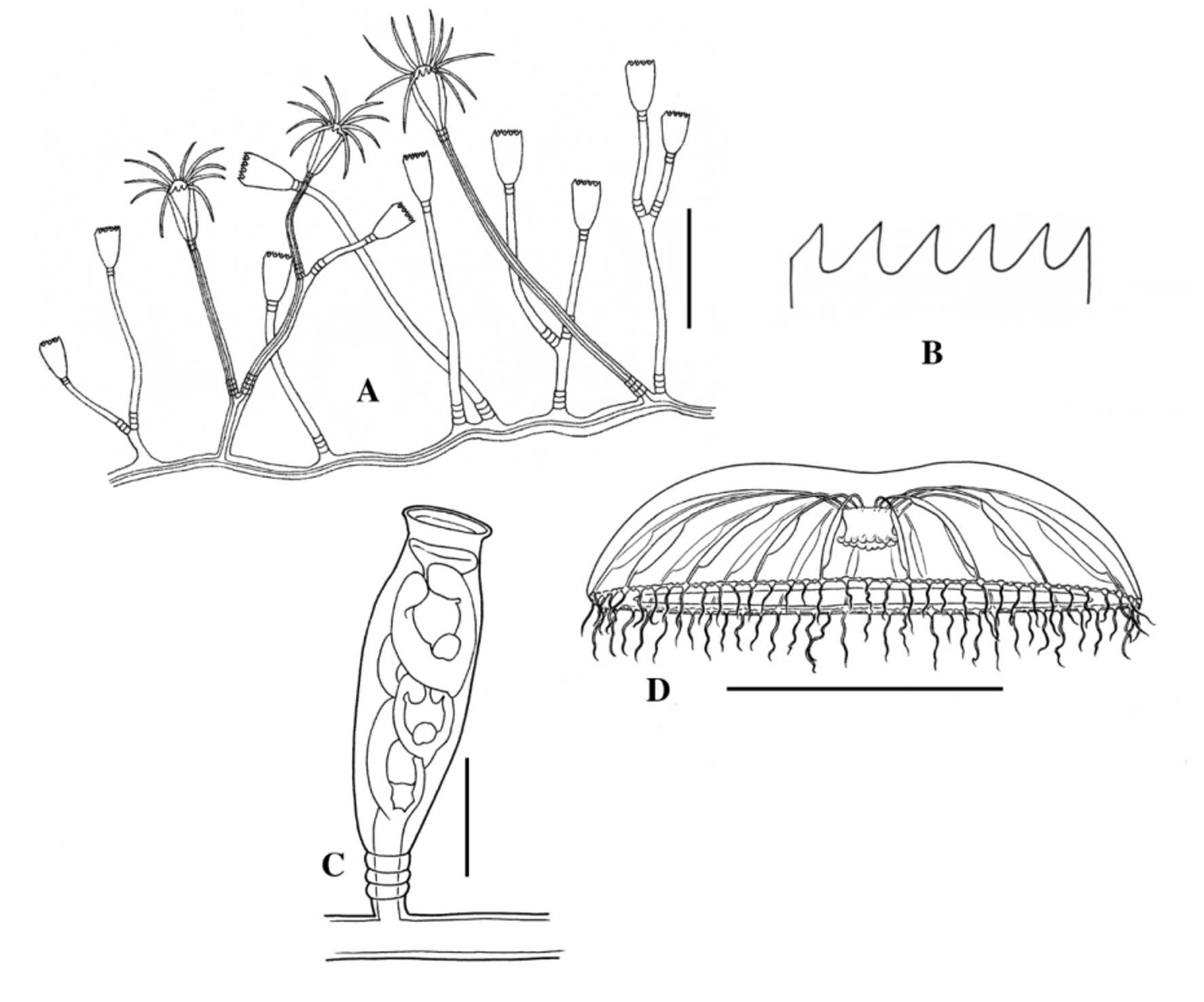
Fig. 100 View FIGURE 100 A–D
See Peña Cantero & García Carrascosa (2002) for a complete synonymy.
Material examined. HCUS-S 107p and HCUS-S 107m (Hydrozoa Collection, University of Salento—fauna of the Salento Peninsula)—polyp and medusa stages.
Description (based on our own observations; Ramil & Vervoort 1992; Cornelius 1995; Schuchert 2001a):
Hydroid. Hydrorhiza as creeping stolon system; colonies erect, delicate, up to 20 mm high; hydrocauli forking 2–3 times, with long internodes and pedicels, annulated; hydranth highly extensile, with 18–20 amphicoronate tentacles; hydrothecae deep-campanulate, with 8–12 large pointed cusps inclined to one side, diaphragm thin, pedicel base incurving. Colour: hydrorhiza yellowish. Gonothecae long, urn-shaped, constricted abruptly below rim and below this more gradually, smooth walled, wide mouthed, arising from hydrorhiza on short, slender pedicels; up to 4 developing medusae per gonotheca.
Habitat type. Littoral and shelf species recorded in the Mediterranean from 5 to about 200 m depth (Gili 1986; Ramil & Vervoort 1992).
Substrate. Algae, bio-concretions, and rock, hydroids, sponges, bryozoans, mollusc shells, polychaete tubes, and cirripedes.
Seasonality. April–December (Puce et al. 2009) in the Ligurian Sea; January–December (De Vito 2006; this study) in Salento waters.
Reproductive period. January (De Vito 2006; this study), June ( Stechow 1919), July ( Peña Cantero & García Carrascosa 2002), August ( Peña Cantero & García Carrascosa 2002; C. Gravili unpublished observations), September (Gili 1986; Puce et al. 2009), October (De Vito 2006; this study) and November ( Broch 1933; De Vito 2006; this study) in several localities of the Mediterranean Sea.
Medusa. Adult. Umbrella nearly hemispherical, up to 8 mm wide, mesoglea thin; manubrium small, quadrate to cruciform; mouth with 4–8 simple lips; up to 22 radial canals; up to 14 gonads oval to round, with few eggs, along the third distal quarter of the radial canals; up to 40 marginal bulbs with spherical base; each marginal bulb bearing one tentacle; 1–2 statocysts between successive tentacles.
Developmental stages. Newly released medusa with umbrella 0.7 mm wide; manubrium small quadrate to cruciform; mouth with 4 simple lips; 4 straight radial canals; 4 marginal bulbs with spherical base; 4 tentacles; 1–2 statocysts between successive tentacles. After one day the marginal bulbs are 8 and all tentaculate. In the 4-day-old medusa the radial canals are 4; marginal bulbs are 12 with 8 tentacles. In the 12-day-old medusa the umbrella is hemispherical; the mouth has 8 lobes; there are 8 radial canals (only 4 are complete until the umbrellar margin; 24 tentaculate bulbs. Twenty-one-day-old medusa with 8 radial canals; 32 tentaculate bulbs; 8 gonads near the umbrellar margin.
Cnidome. b-mastigophores 1 type (polyp), isorhizas (medusa).
Distribution. Atlantic, Indo-Pacific, Mediterranean for the hydroid stage, planktonic colonies have been described in Atlantic waters ( Medel & López-González 1996; Migotto 1996; Peña Cantero & García Carrascosa 2002; Bouillon et al. 2004; Gravili et al. 2008a; Morri et al. 2009; Soto Ãngel & Peña Cantero 2013).
Records in Salento. Common at: Grotta del Ciolo ( Moscatello & Belmonte 2007); Il Chiapparo (C. Gravili unpublished observations); Otranto (De Vito 2006; Gravili 2006; Andreano 2007; Gravili et al. 2008a; Ventura 2011; Piraino et al. 2013; this study).
Remarks. The newborn medusa is often confounded with other Clytia medusae, mainly with C. hemisphaerica , although the discrimination between different species may perhaps be possible by the use of cnidocyst micromorphology ( Östman 1979a, 1983). It seems to be a circumglobal species, reaching sub-Arctic ( Naumov 1969) and sub-Antarctic waters (Stepanjants 1979), although previous confusion with the polyp stage of C. hemisphaerica makes it difficult to establish an exact distributional pattern for this species (see Lindner et al. 2011). Moreover, the medusa described in this paper, having multiple radial canals, shows several differences from the description of the medusa reported in Cornelius (1995): probably they are different species. The hydroid stage and the newborn medusa are clearly referable to Clytia , but the adult medusa is not part of the classical definition of the genus (see Bouillon et al. 2006) and calls for a re-assesment of its diagnosis.
References. Broch (1912, 1933) as Laomedea (Clytia) gracilis View in CoL ; Stechow (1919) as Gonothyraea gracilis View in CoL ; Leloup (1934) as Laomedea gracilis View in CoL ; Picard (1958a), Rossi (1961), Naumov (1969), Ramil (1988), Riedl (1991), Ramil & Vervoort (1992), Vervoort (1993), Altuna (1994), Medel & López-González (1996), Medel & Vervoort (2000), Schuchert (2001a), Peña Cantero & García Carrascosa (2002), Bouillon et al. (2004), De Vito (2006), Gravili (2006), Gravili et al. (2008a), Moscatello & Belmonte (2007), Morri et al. (2009), Puce et al. (2009), Lindner et al. (2011), Ventura (2011), Piraino et al. (2013).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.