
Bornella anguilla Johnson, 1984
|
publication ID |
https://doi.org/10.5281/zenodo.185130 |
|
DOI |
https://doi.org/10.5281/zenodo.5689403 |
|
persistent identifier |
https://treatment.plazi.org/id/03F98793-FF8D-722C-2BCF-FD42A527F9DD |
|
treatment provided by |
Plazi |
|
scientific name |
Bornella anguilla Johnson, 1984 |
| status |
|
Bornella anguilla Johnson, 1984 View in CoL
( Figures 1 View FIGURE 1 F, 4E, 13)
Bornella anguilla Johnson, 1984: 17 View in CoL –26, figs. 1–6.
Material examined: Papua New Guinea: CASIZ 0 65273, Papua New Guinea, north coast, near Madang, Cement Mixer reef, approx. W.N.W. of Rasch Pass in line with Ruo Island, 13 January 1988, one specimen 12 mm preserved, coll: T.M. Gosliner. CASIZ 0 65305, Papua New Guinea, northern coast, Bagabag Island, Christmas Bay, north point, 10 February 1988, two specimen 13 & 20 mm preserved, 7 m depth, coll: T.M. Gosliner & R.C. Willan. CASIZ 0 65731, Papua New Guinea, north coast, north of Madang, approx. 1km S. of Cape Croselles, north of the Quarry near Bunn Village, 12 February 1988, one specimen 40 mm preserved, dissected, 9 m depth, coll: T.M. Gosliner. CASIZ 0 65321, Papua New Guinea, north coast, near Madang, west side 'Pig Island " [Tab Island], Barracuda Point, 18 February 1988, one specimen 18 mm preserved, dissected, 9 m depth, coll: J. Mizeu. CASIZ 0 69767, Papua New Guinea, north coast, near Madang, the Pinnacle, between Pig Island [Tab Island] and Rasch Pass, 19 July 1989, one specimen 20 mm preserved, dissected, 30 m depth, coll: T.M. Gosliner. CASIZ 0 68662, Papua New Guinea, north coast, near Madang, east side 'Pig Island " [Tab Island], Barracuda Point, 27 July 1989, 9 m depth, coll: T.M. Gosliner. CASIZ 0 75820, Papua New Guinea, north coast, near Madang, east side 'Pig Island " [Tab Island], Barracuda Point, 11 November 1990, one specimen 14 mm preserved, 24 m depth, coll: T.M. Gosliner & G. Williams. CASIZ 0 75081. Papua New Guinea, north coast outer barrier reef, Bagabag Island, off New Year's Bay, 26 November 1990, one specimen 15 mm preserved, 30 m depth, coll: T.M. Gosliner. CASIZ 0 86441, Papua New Guinea, north coast, near Madang, approx. W.N.W of Rasch Pass in line with Ruo Island, Cement Mixer Reef, 0 4 June 1992, one specimen 25 mm preserved, dissected, 25 m depth, coll: T.M. Gosliner. CASIZ 0 86312, Papua New Guinea, north coast, near Madang, east side Pig Island [Tab Island], Barracuda Point, 14 June 1992, two specimens 19 & 30 mm preserved, dissected, 25 m depth, coll: T.M. Gosliner. CASIZ 109794, Papua New Guinea, Madang Province, Planet Rock ( 10 km south of Madang), 3 November 1996, one specimen 47 mm preserved, 26 m depth, coll: T.M. Gosliner. Philippines: CASIZ 175854, Philippines, Panglao, Balacasag Island, 30 May 1994, one specimen, 5-7 m depth, coll: Y. Camacho. South China Sea: CASIZ 0 71921, South China Sea, China, Hainan Dao, South of Ya Xian, Double Sail Rock, 0 7 April 1986, one specimen 65 mm preserved, dissected, 3 m depth, coll: D. Blair. Japan: CASIZ 0 70213, Japan, Ryukyu Island, Okinawa, 1 km W.N.W of Onna Village, Horseshoe Cliffs, 26 30'N, 127 50.9'E, 20 August 1989, one specimen 41 mm preserved, 49 m depth, coll: R.F. Bolland. CASIZ 0 84848, Japan, Ryukyu Island, Okinawa, 8 km N.N.E. of Kuzu-saki, off Kadena Air Force Base runway lights, 26 20'N, 127 44.7'E, 23 December 1991, one specimen 21 mm preserved, dissected, 12 m depth, coll: R.F. Bolland. CASIZ 174984, Japan, Ryukyu Island, Okinawa, Horseshoe Cliffs. 1 km W.N.W of Onna Village, 0 3 April 1994, two specimens 54 & 41 mm preserved, 58 m depth, coll: R.F. Bolland. CASIZ 174982, Japan, Ryukyu Island, Okinawa, Horseshoe Cliffs. 1 km W.N.W of Onna Village, 21 May 1994, one specimen 44 mm preserved, 68 m depth, coll: R.F. Bolland. CASIZ 174983, Japan, Ryukyu Islands, Okinawa, Horseshoe Cliffs. 1 km W.N.W of Onna Village, 18 April 1998, one specimen 40 mm preserved, 68 m depth, coll: R.F. Bolland. CASIZ 174981, Japan, Ryukyu Islands, Okinawa, Horseshoe Cliffs. 1 km W.N.W of Onna Village, 18 May 1994, one specimen 45 mm alive, 68 m depth, coll: R.F. Bolland. Pacific Ocean: CASIZ 120721, Pacific Ocean, Marshall Islands: Kwajalein Atoll, Ennubuj Island, Gea Island, ocean side of Island, in ledge at night, 24 October 1992, one specimen 20 mm preserved, 15 m depth, coll: J. Johnson. CASIZ 0 71930, Pacific Ocean, Marshall Islands, Kwajalein Atoll, Seaward side of Ennubuj- Ennylabagan Reef, November 1992, one specimen 38 mm preserved, dissected, 15 m depth, coll: J. Wedge. Australia: CASIZ 144026, Australia, Queensland, Mooloolaba, S.E. Queensland, 28 December 2000, two specimens 50 & 58 mm preserved, dissected, coll: S. Fahey. CASIZ 168860, Australia, Queensland, Mooloolaba, S.E. Queensland, outer Gneerings, Magic Mountain, 0 8 July 2004, one specimen 45 mm preserved, 23 m depth, coll: S. Fahey & T.M. Gosliner. Madagascar: CASIZ 173389, Madagascar, Iles Radama, Nosy Kalakajoro, West between Nosy Kalalajoro and Nosy Berafia, 19 November 2005, one specimen 20 mm preserved, dissected, 17 m depth, coll: S. Fahey.
Geographic distribution: Indo-West Pacific: Marshall Islands, Western Australia, Queensland ( Johnson 1984; Coleman 1989, 2001; Debelius 1998; Gosliner et al. 1996; Marshall & Willan 1999; Taylor 2001; present study), South Africa ( Johnson 1984; Gosliner 1987a; Fraser 2000), Papua New Guinea, Japan ( Okutani 2000; present study), Madagascar (present study), Philippines ( Adriance 2006; present study), China (present study), Mauritius ( Yonow & Hayward 1991), Vietnam (www.daveharasti.com), Thailand ( Beasley 2006), Indonesia ( Henke 2006; Krampf 2006), Reunion Island ( Coze 2003; Bidgrain 2005), New Zealand (www.underwater.com.au/image.php/id/6616/), Hawaii (www.slugsite.us/bow/nudwk143.htm), Fiji ( Tucker 2001), Burma ( Adams 2003; Aytur 2003) and Vanuatu (present study).
External morphology: The general body shape is elongate and limaciform with the posterior end of the foot being long and tapering ( Fig. 1 View FIGURE 1 F). This relatively large species grows up to 120 mm ( Henke 2006) but they are usually 40–50 mm long. This species has a characteristic mosaic-like color pattern on the body ( Fig. 1 View FIGURE 1 F). The general color consists in a dark brown network enclosing variable sized patches, which are whitecream, yellow, orange or light brown. This color pattern extends to the stalk of the rhinophore sheaths. There are scattered opaque white subepidermal granules. The papillae on the oral tentacles and the upper edge of the rhinophore sheaths are orange with white tips. The posterior sail of the rhinophore sheaths is bordered on each edge by an orange band and there is a central, vertical, dark brown stripe down each side. The upper paddlelike flap of the dorsolateral processes has the same coloration as the posterior sail of the rhinophore sheaths and their bases are cream-yellow with a light brown or purple vertical band on each side. There are the characteristic lobe-like oral tentacles on each side of the mouth. It consists of a flattened lobe bearing a single row of up to six elongate papillae arranged around the edge. Each rhinophore sheath is tall and stalk-like with the rhinophore protruding from the tip. The rhinophores are perfoliate with about 15 to 25 lamellae, light peach in color. Surrounding the upper edge of the rhinophore sheath, are three long and narrow anterior and anterolateral papillae and a taller, posterior, very laterally compressed sail ( Fig. 1 View FIGURE 1 F). The posterior sail is un-branched. Posterior to the rhinophores there are three pairs of dorsolateral processes, followed by three unpaired processes along the dorsal midline. The number of processes was consistent in all specimen examined. All the dorsolateral processes have a stout base topped with a paddle-like flap similar in shape and color to the posterior sail of the rhinophore sheath. There are three tripinnate, translucent white gills on each of the paired dorsolateral process. The gills are found on the inner surface. They appear to be absent from the unpaired processes in the dorsal midline, which are simple, flattened and decreasing in size towards the end of the tail. The anus is located on the right side of the dorsum between the first and second pair of dorsolateral processes, closer to the second. The reproductive opening is located on the right side, midway between the rhinophore sheath and the first dorsolateral process.
Alimentary Canal: The buccal bulb is relatively large. The labial cuticle is thick, with irregular cuticular rodlets surrounding the opening into the buccal bulb. The rodlets are elongate and numerous ( Fig. 13A View FIGURE 13. A – I ). The jaws are roughly oval in shape, without a distinct masticatory process ( Fig. 13B View FIGURE 13. A – I ). The radular formulae of some of the specimens dissected are: 94 x 14.1.14 (CASIZ 0 71930, 38 mm preserved), 57 x 17 – 19.1.17 –19 (CASIZ 0 65731, 40 mm preserved; CASIZ 144026, 50 mm preserved), 40 x 12 – 13.1.12 –13 (CASIZ 0 69767, 20 mm preserved, CASIZ 0 86441, 25 mm preserved), 37 x 10.1.10 (CASIZ 0 86312, 19 mm preserved), 37 x 13.1.13 (CASIZ 173389, 20 mm preserved), 33 x 10.1.10 (CASIZ 0 65321, 18 mm preserved) and 31 x 11.1.11 (CASIZ 0 84848, 21 mm preserved). The rachidian teeth are slightly wider than taller, with about 7 to 11 denticles on either side of a strong median cusp ( Figs. 13D,E View FIGURE 13. A – I ). The laterals are blade-like with a long basal portion, increasing in size away from the rachidian tooth, but usually the two or three outermost teeth decrease slightly in size ( Fig. 13D View FIGURE 13. A – I ). A long unpaired oral gland is found ventrally ( Fig. 13C View FIGURE 13. A – I ). A pair of yellowish, elongate-branched salivary glands is attached to the posterior side of the oesophagus. They open into the posterior buccal bulb, one on each side of the oesophageal opening. The oesophagus is relatively long and narrow, and runs back to the oval thin walled stomach. Attached to the dorsal side of the stomach are the anterior digestive glands, each of which forms a little sac and a duct into the most anterior of the dorsolateral process on its respective side. There are no branches from the anterior digestive glands to the rhinophore sheaths. The opening of the posterior digestive gland is on the lower left surface of the stomach. All pairs of dorsolateral processes, behind the first pair, receive branches from the posterior portion of the digestive gland. The posterior stomach is armed with about 30 longitudinal rows of chitinous brown spines ( Figs. 13F,G View FIGURE 13. A – I ). These spines are straight, cylindrical, and rounded at the tip ( Fig. 13G View FIGURE 13. A – I ). The spines decrease in size posteriorly. From the stomach the intestine descends to the ventral side and then bends dorsally to the anus.
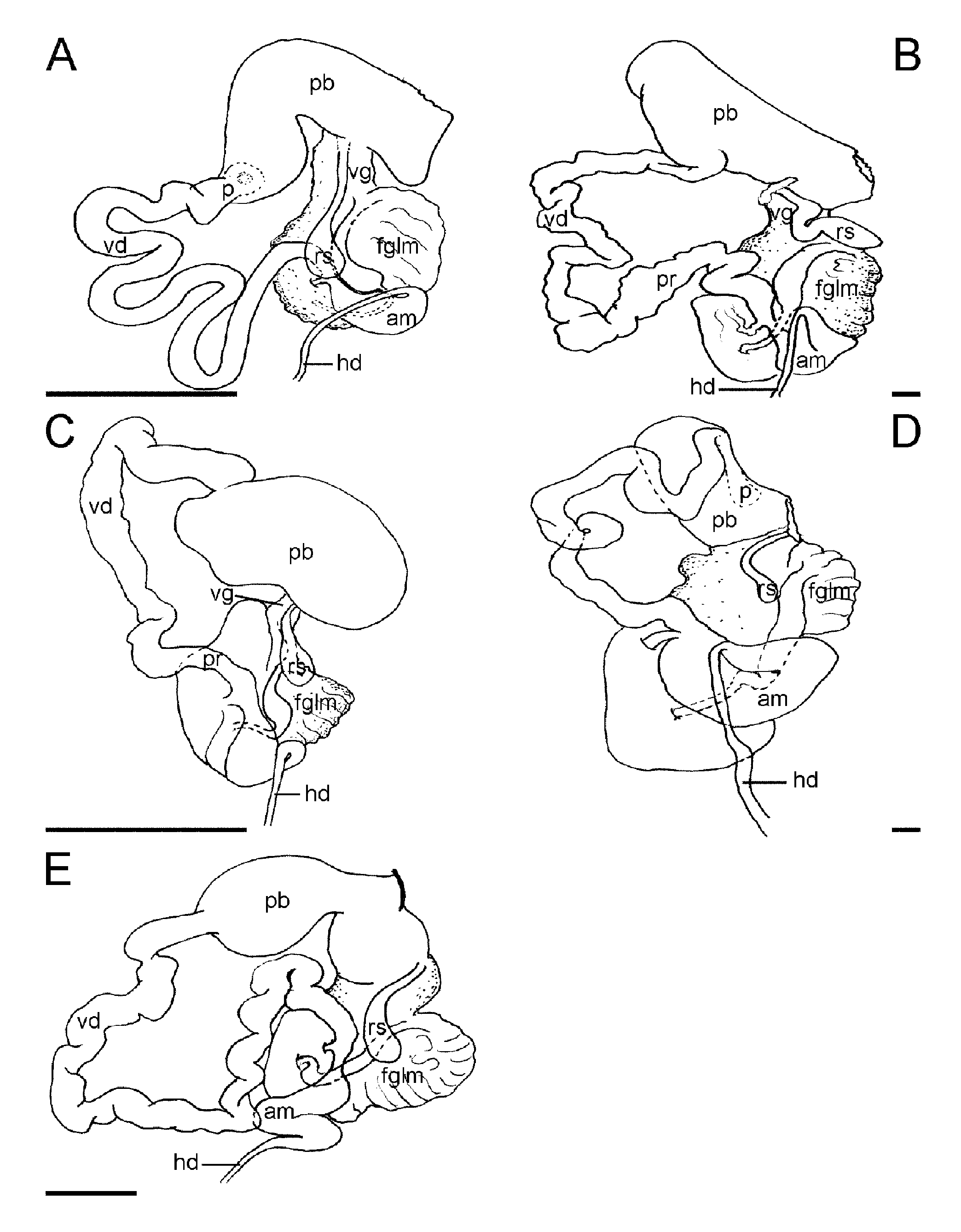
Reproductive system: ( Fig. 4E View FIGURE 4. A – E , F): The ovotestis consists of 10 to 12 rounded follicles lying over the posterior digestive gland. Each follicle has a thin duct connecting it to the hermaphrodite duct which runs around the stomach on the left side and expands into the S-shaped ampulla on the dorsal side of the female gland mass. At its anterior end, the ampulla region narrows into a short duct, which divides to form the vas deferens and the oviduct. The visible oviduct is quickly embedded in to the female gland mass, and without histological examination its path is impossible to determine with any accuracy. The vas deferens is long and folded and appears to be covered with a uniform layer of prostate gland. The penial sac contains a fleshy disclike structure, which has been described as the penial papilla ( Johnson 1984). This structure has an irregular ring of chitinous hooks around its inner edge ( Figs. 13H,I View FIGURE 13. A – I ). A medium distal pyriform allosperm receptacle opens directly at the genital aperture through a long duct.
Remarks: A key diagnostic for this species is the characteristic mosaic-like color pattern on the body. Also, the shape of the posterior sail of the rhinophore sheaths and dorsolateral processes, as well as the arrangement of the penial spines, is unique within the genus. Bornella anguilla is a large species reported to feed on hydroids of the genus Plumularia . Johnson (1984) reported it to be strictly nocturnal in the Marshall Islands but it can be found out in the open in broad daylight. The name " anguilla " meaning eel, refers to its method of swimming. Most species of Bornella can swim by a lateral flexion of their body; B. anguilla uses a muscular wave that travels down the body to produce an eel-like swimming motion ( Johnson 1984).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bornella anguilla Johnson, 1984
| Pola, Marta, Rudman, William B. & Gosliner, Terrence M. 2009 |
Bornella anguilla
| Johnson 1984: 17 |