
Zatavua, Huber, 2003
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2003.00046.x |
|
persistent identifier |
https://treatment.plazi.org/id/03F987F6-FFFA-FFB8-FC21-0CA53629F8A3 |
|
treatment provided by |
Carolina |
|
scientific name |
Zatavua |
| status |
gen. nov. |
ZATAVUA View in CoL View at ENA , NEW GENUS
Type species. Zatavua griswoldi , new species.
Etymology. Named for Zatavu, a great magician in Malagasy mythology who created himself and was therefore allowed to marry a goddess. Gender feminine.
Note. Millot (1946) described several species belonging to this genus (under Spermophora ). Unfortunately, most of the material studied by him has not been available to me (it could not be found at the MNHN in Paris; C. Rollard, pers. comm.). Therefore, the diagnosis and description given below may not perfectly cover all the species included. Nevertheless, judging by Millot’s excellent illustrations, some of his species seem very closely related to some of the species newly described herein (see Specific Relationships below), so the diagnosis and description may in fact be tolerably accurate.
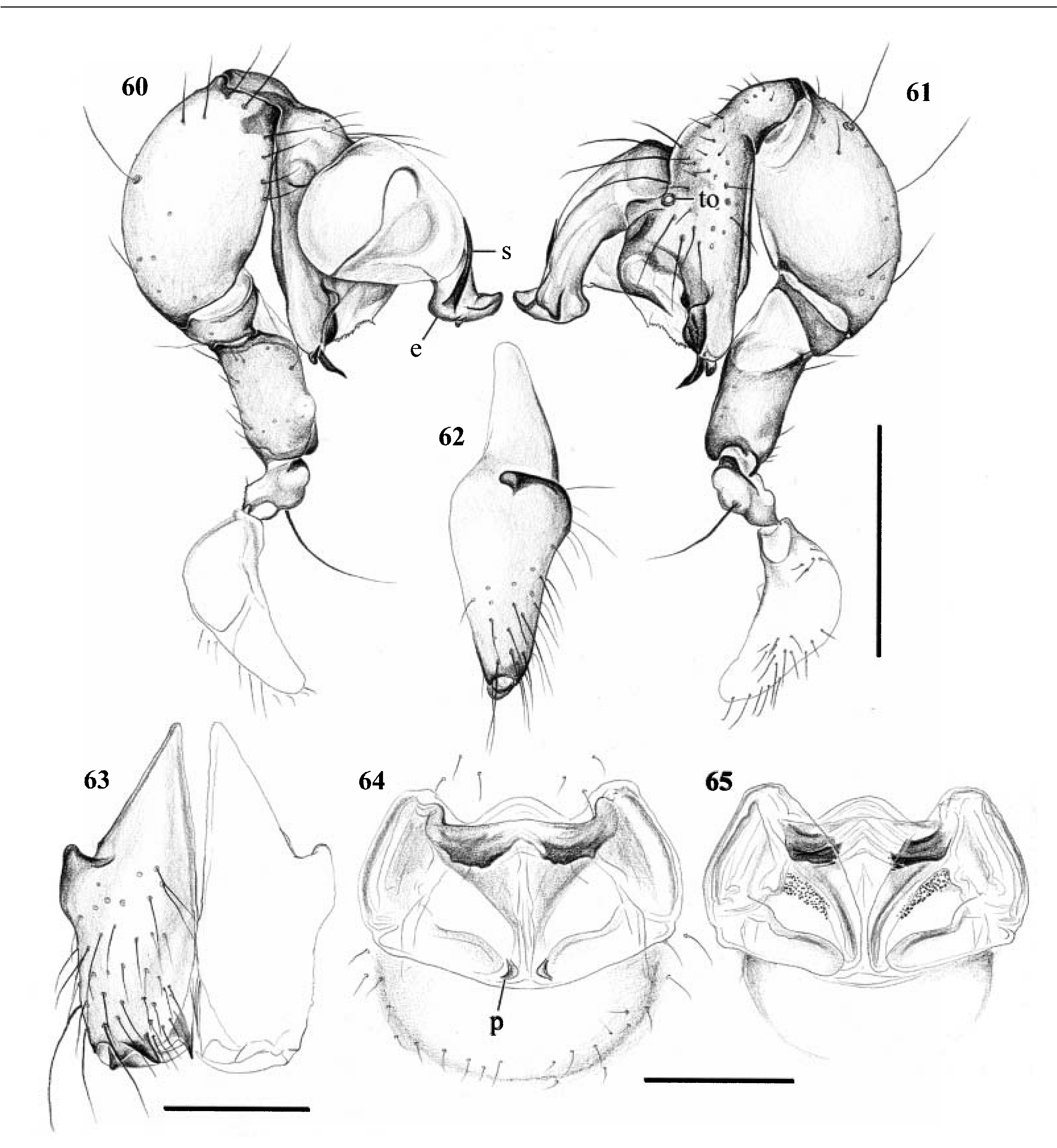
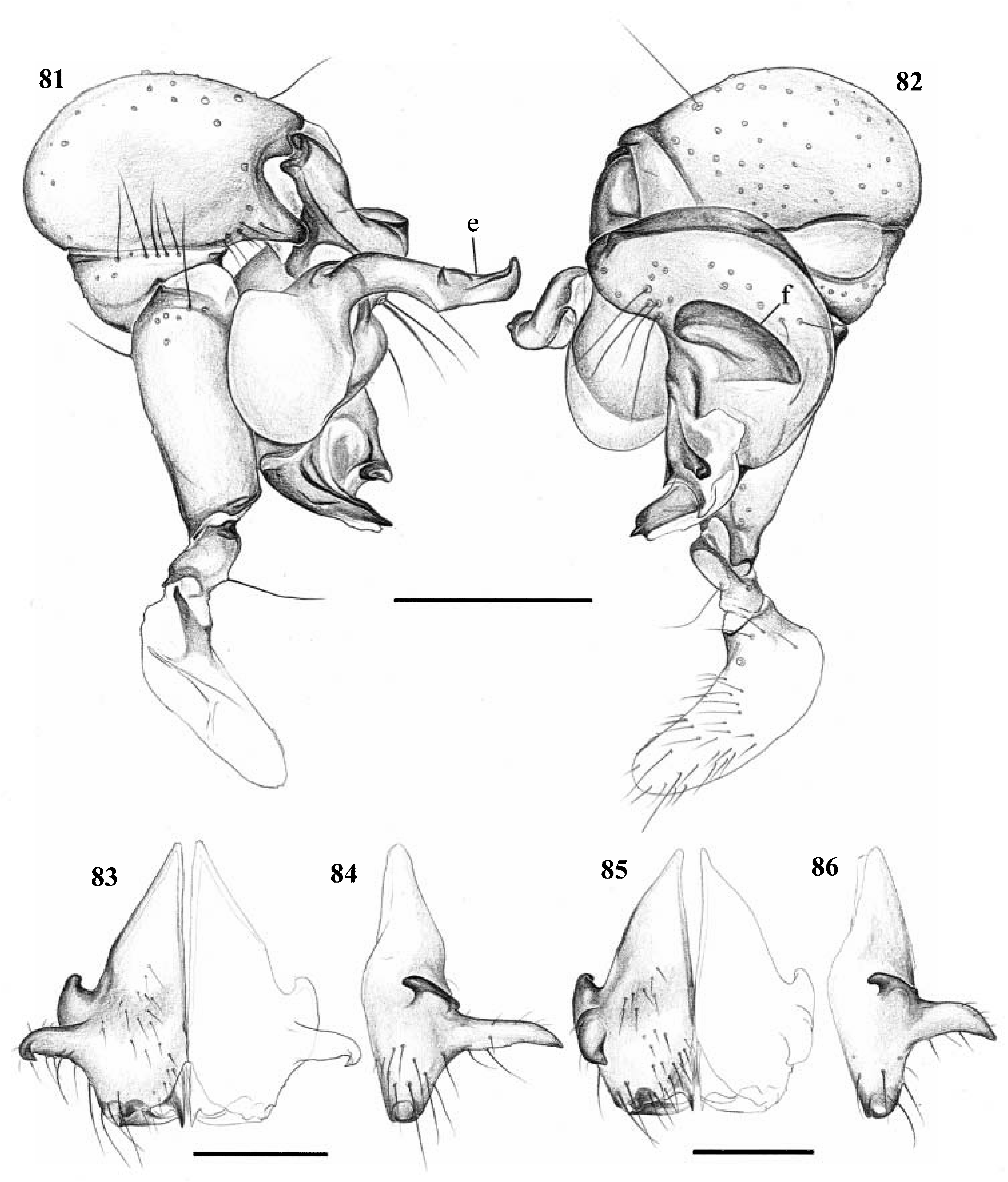
Diagnosis. Long-legged, six-eyed pholcids with globular or elongate opisthosoma and variable size (total length ~ 2–8 mm). Distinguished from Paramicromerys and other six-eyed genera by the lateral cheliceral apophyses pointing backwards ( Figs 62 View Figures 60–65 , 84, 86 View Figures 81–86 and 98 View Figures 96–101 ), the presence of a retrolateral notch on the cymbium ( Figs 34 View Figures 33–37 , 48 View Figures 47–50 and 57 View Figures 56–59 ), and by the shift of the tibia-tarsus joints, resulting in a dorsal position of the retrolateral joint and in a ventral position of the prolateral joint ( Figs 29, 30 View Figures 29–32 , 33 and 34 View Figures 33–37 , etc.; two latter characters missing in Z. kely , mahafaly ). Distinguished from Paramicromerys also by the closeness of the triads (e.g. Fig. 67 View Figures 66–77 ; PME-PME <180% of PME diameter, vs.>230% in Paramicromerys ), and by the presence of several spigots on the ALS in addition to the basic set of two ( Figs 39, 40, 46 View Figures 38–46 , 74, 75 and 77 View Figures 66–77 ).
Description. Total length in males usually ~ 3–5 mm; only Z. zanahary and punctata (Millot) up to 8 mm, Z. kely only ~ 1.5 mm. Carapace oval or round, with or without thoracic groove, often with dark pattern that may be distinctive ( Figs 1–18 View Figures 1–18 ). Six eyes in two triads, on moderately elevated ocular area; no trace of AME. Distance PME-ALE small (~20–40% of PME diameter), distance PME-PME also small (~90–180% of PME diameter). Clypeus in some species with paired unsclerotized projections at rim. Male chelicerae with pair of lateral apophyses pointing backwards ( Figs 62 View Figures 60–65 , 84, 86 View Figures 81–86 and 98 View Figures 96–101 ), and frontal apophyses variable in position and shape; never with modified hairs, never with stridulatory ridges. Male palps moderately large in relation to overall size; coxa and trochanter without retrolateral apophyses, femur usually cylindrical, sometimes with small ventral apophysis proximally; tibia moderately to highly expanded, with two trichobothria; tibia-cymbium joints distinctively shifted (resulting in dorsal position of retrolateral joint and ventral position of prolateral joint); cymbium with distinctive notch on retrolateral side ( Figs 34 View Figures 33–37 , 48 View Figures 47–50 and 57 View Figures 56–59 ); procursus usually complex but never with hinged process; with capsulate tarsal organ ( Fig. 73 View Figures 66–77 ), often in very distal position ( Figs 30 View Figures 29–32 , 33 View Figures 33–37 , 47 View Figures 47–50 , 52 View Figures 51–55 , 56 View Figures 56–59 , 61 View Figures 60–65 and 78 View Figures 78–80 ); bulb consisting of proximal globular part and embolus that is partly sclerotized and often provided with sclerotized spine. Legs usually long (leg 1 about 8– 11 ¥ body length; in Z. kely only 6 ¥), medium-thin (tibia 1 L/d ~40–90), leg 1 always longest, legs 2 and 4 about same length, leg 3 shortest. Legs usually without spines, with few vertical hairs, without curved hairs; retrolateral trichobothrium of tibia 1 usually at 5–15%, in Z. kely at 27%. Prolateral trichobothrium missing on tibiae 1, present in all others. Tarsus 1 with over 20 pseudosegments, but only ~10–20 distal pseudosegments easily visible in dissecting microscope. Opisthosoma either globular or elongate, often with posterior elongation over spinnerets ( Figs 4, 14 and 16 View Figures 1–18 ). Male gonopore with four epiandrous spigots in all species examined ( Z. zanahary , griswoldi , voahangyae ; Figs 41 View Figures 38–46 and 66 View Figures 66–77 ); ALS with several spigots in addition to basic set of two (examined: Z. griswoldi , vohiparara , voahangyae , isalo , kely ; Figs 39, 40, 46 View Figures 38–46 , 74, 75 and 77 View Figures 66–77 ); other spinnerets typical for family (e.g. Fig. 38 View Figures 38–46 ; cf. Huber, 2000).
Sexual dimorphism slight. In some species females with median conical elevation posteriorly on carapace; corresponding side of opisthosoma not visibly modified. Epigynum shape very variable, often with pair of pockets, in Z. vohiparara with short scape ( Fig. 45 View Figures 38–46 ); internally with pair of relatively large pore plates and complex system of sclerites and membranes of unknown function.
Monophyly. The monophyly of this genus is supported by the following three synapomorphies: the backward facing lateral cheliceral apophyses (shared by all species included), the shift of the tibia-cymbium joints, and the notch on the cymbium (the latter two characters are not shared by Z. kely and Z. mahafaly ).
Generic relationships. As discussed above, Zatavua is here considered the sister taxon of all other pholcines. The presence of lateral apophyses on the male chelicerae places the genus within the subfamily, but the genus lacks the retrolateral trochanter apophysis characteristic of other pholcines.
Specific relationships. Several synapomorphies support species groups within Zatavua . A core group of species is characterized by the presence of a conical elevation on the female carapace [ Z. griswoldi , tamatave , vohiparara , maybe including andrei (Millot) and ankaranae (Millot)]. This group shares with Z. analalava the projections on the male clypeus. The resulting group shares with several further species [ Z. zanahary , punctata (Millot) , voahangyae , talatakely , probably impudica (Millot)] the distal position of the male palpal tarsal organ. Within this latter group, Z. zanahary and punctata share their unusual size and the unique pattern on the carapace ( Fig. 9 View Figures 1–18 ). All species listed so far share with Z. isalo and madagascariensis (Fage) the shift of the tibia-cymbium joints and the notch on the cymbium. The latter two species have almost identical palps (not coded) and are therefore seen as closely related. Zatavua fagei (Millot) is only known from the female, but was collected near the type locality of Z. madagascariensis (Fage) and is either a junior synonym or a closely related species. Finally, Z. mahafaly and kely are very unusual in several aspects, but share with all other species the backward facing lateral cheliceral apophyses on the male chelicerae. Of these, Z. kely might be close to imerinensis (Millot) , judging by the similar shapes of the palps.
Natural history. No species has ever been studied in any detail, and little can be inferred from notes on the collection labels. Several species have been collected in caves [ Z. andrei , ankaranae, impudica, madagascariensis (Fage) , mahafaly ], two more were classified as ‘semi-cavernicole’ by Millot (1946) ( Z. fagei , punctata ), and one as ‘semi-domestique’ ( Z. imerinensis ). All new species described below (except Z. mahafaly ) were apparently collected in forests. Zatavua vohiparara was collected on Pandanus , an undescribed species from the AMNH was collected by beating vegetation.
Distribution. Known from Madagascar only. The genus is widely distributed in Madagascar (Map 1), and probably covered most of the island before human destruction of a large part of the primary vegetation. As in Paramicromerys , no species has been collected at two localities more than a few kilometres apart. A possible exception is Z. punctata (Millot) , which has been recorded from the Antonibe Peninsula and on Nosy- Komba Island (about 200 km apart), but the latter record is based on a female and a juvenile specimen and should be checked.
Composition. The genus includes a total of 17 described species. Of these, six were described by Millot (1946), and are not treated below ( Z. andrei , ankaranae, fagei , imerinensis , impudica, punctata ). The remaining 11 species [10 new species and Z. madagascariensis (Fage) ], are (re)described below. The collections studied contain four additional new species that are not described due to their poor state of preservation. Considering the facts that (1) most or all species seem to have small distributional ranges, and that (2) all areas that were subjected to intense collecting yielded between one and three species, it seems reasonable to expect several dozen additional species not yet discovered.
ZATAVUA GRISWOLDI , NEW SPECIES
( FIGS 1, 2 View Figures 1–18 , 19 View Figures 19–28 , 29–32 View Figures 29–32 , 38–44 View Figures 38–46 )
Type. Male holotype from Marojejy Res. , 8.4 km NNW Manantenina (14∞26¢S, 49∞45¢E), 700 m a.s.l., Antsiranana, Madagascar; November 10–16, 1993 (C. E. Griswold, J. Coddington, N. Scharff, S. Larcher, R. Andriamasimanana); in CAS .
Etymology. Named for the first collector, Charles Griswold from the California Academy of Sciences, who kindly made his superb collection of Malagasy pholcids available to me.
Diagnosis. Medium size species with elongated opisthosoma, distinguished from known congeners by the shape of procursus and embolus ( Figs 29, 30 View Figures 29–32 : overall shape and several unnamed details), and by the shape of the epigynum ( Figs 19 View Figures 19–28 and 31 View Figures 29–32 ). The AMNH has a male from a nearby locality (10.5 km NW Manantenina) that differs slightly by having a shorter procursus and more profoundly by the shape of the embolus. The MRAC has a further close relative from











No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.