
Harajicadectes, 2024
|
publication ID |
https://doi.org/ 10.1080/02724634.2023.2285000 |
|
publication LSID |
lsid:zoobank.org:pub:A21E263E-9B2B-4C9B-9B1B-74B53C2CF535 |
|
DOI |
https://doi.org/10.5281/zenodo.10642030 |
|
persistent identifier |
https://treatment.plazi.org/id/BE175E16-EEE6-4B1C-B08F-64A341F47C71 |
|
taxon LSID |
lsid:zoobank.org:act:BE175E16-EEE6-4B1C-B08F-64A341F47C71 |
|
treatment provided by |
Felipe |
|
scientific name |
Harajicadectes |
| status |
gen. nov. |
HARAJICADECTES ZHUMINI , gen. nov. et sp. nov.
( Figs. 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
‘rhipidistian crossopterygian’: Young, 1985:246.
‘rhipidistian osteichthyan’: Young, 1991:107.
‘osteolepid indet.’: Young, Long and Turner, 1993:248.
‘articulated sarcopterygians’: Young and Turner, 2000:107.
‘rhipidistian crossopterygian’: Clement, 2009:615.
LSID. zoobank.org:pub: A21E263E-9B2B-4C9B-9B1B-74B53 C2CF535
Holotype — NTM P6410 : partial skull and much of the body outline in dorsal view with squamation preserved.
Paratype — CPC 39948 : partial skull, right post-temporal and right lower jaw.
Referred Material — AMF 99703: pectoral fin elements; AMF 99704: crushed skull with mandibles in dorsal view; CPC 24700: section of squamation and fragmentary jaw elements. CPC 24702: denticulated tooth surface, either a partial entopterygoid or prearticular dental plate. CPC 39949: skull in palatal view (39949A) and a detached fragment of the snout tip (39949B); CPC 39950: partial palate; CPC 39952: partial skull, left pectoral fin and scales from the anterior half of body; CPC 39953: rear part of the body, comprising scales, median fins, and a fragmentary pelvic fin; CPC 39954: detached pectoral fin with basal scute superimposed over flank squamation preserved in visceral aspect; NMVP 229354: partial palate and scales from the anterior body; NMVP 228722: partial skull roof, left post-temporal, and right lower jaw in dorsal view. NMVP 229368: partial opercular-gular series, pectoral girdle, and body scales; NMVP 253964: right mandible in lingual view; NMVP 253965: fragmentary right mandible and dermopalatine (253965A), and an associated complete left maxilla in mesial view (253965B); NTM P6411: section of vertebral column and associated scales.
Etymology —“Harajica Biter.” Named for the Harajica Sandstone Member and the ancient Greek dektes (“biter”) in reference to the animal’ s large fangs and presumed predatory habits. The species honors Professor Min Zhu of the Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China, for his numerous contributions to early vertebrate paleontology.
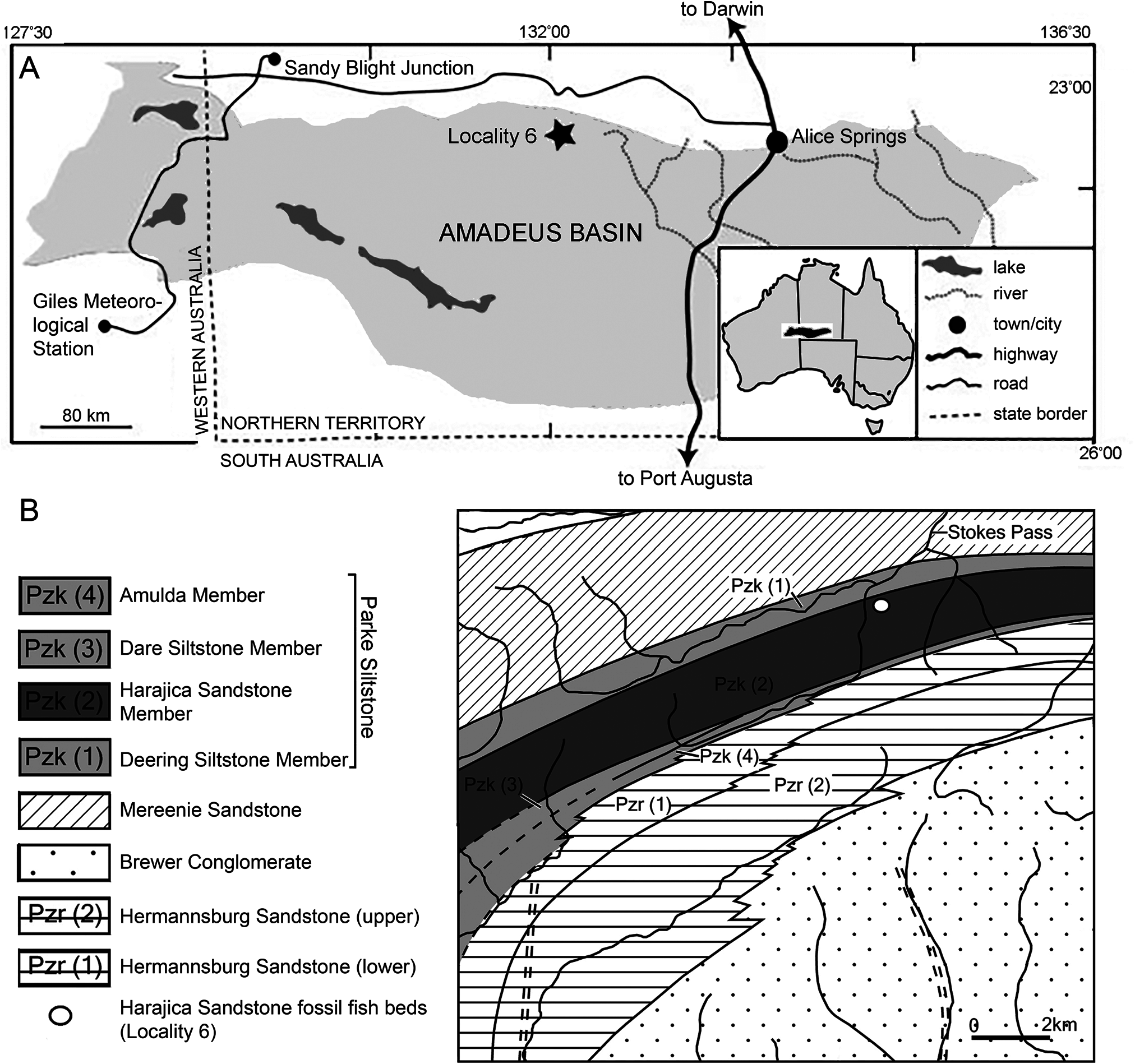
Type Locality and Horizon —Harajica Sandstone Member of the Parke Siltstone. Locality 6 of Young (1985), about 2 km southwest of the southern end of Stokes Pass, Amadeus Basin , Northern Territory. Givetian–Frasnian in age ( Fig. 1 View FIGURE 1 ).
Diagnosis —Tetrapodomorph fish with greatly enlarged spiracular openings, comprising over 20% of the total length of the skull-roof, bordered by the tabular, extratemporal, and squamosal. Parietal and postparietals of roughly equal length. Elongate intertemporal that widens anteriorly. Posterior nasals narrower than the anterior and posterior supraorbitals. Median extrascapular tapers anteriorly into a V-shaped recess formed by the large rounded lateral extrascapulars. Elongate lanceolate parasphenoid with denticles larger on the anterior part of the bone. Scales display ridged ornamentation and lack cosmine. Anterior squamation cycloid, abruptly shifting to a rhombic shape on the rear flank.
DESCRIPTION
General Features
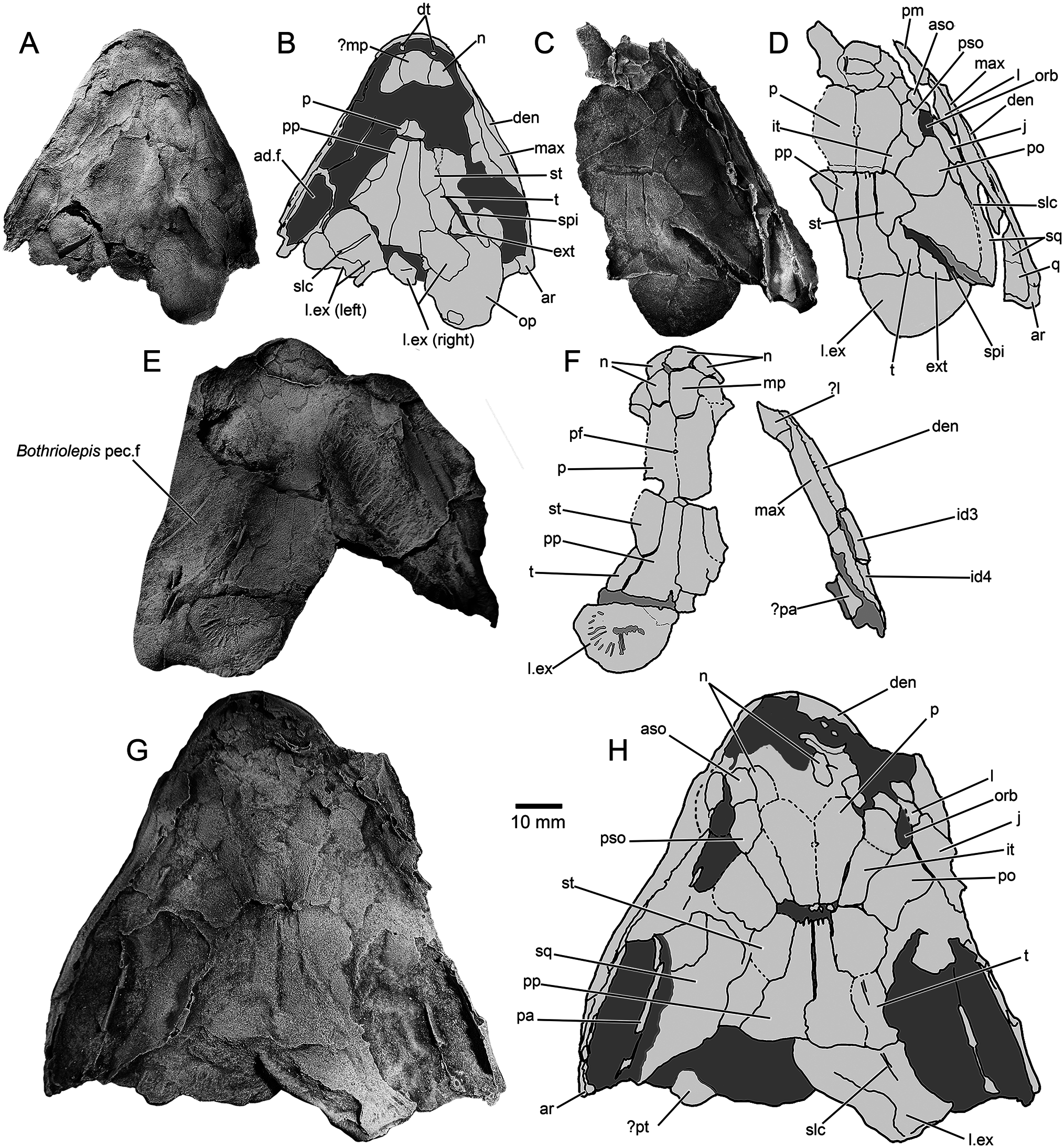
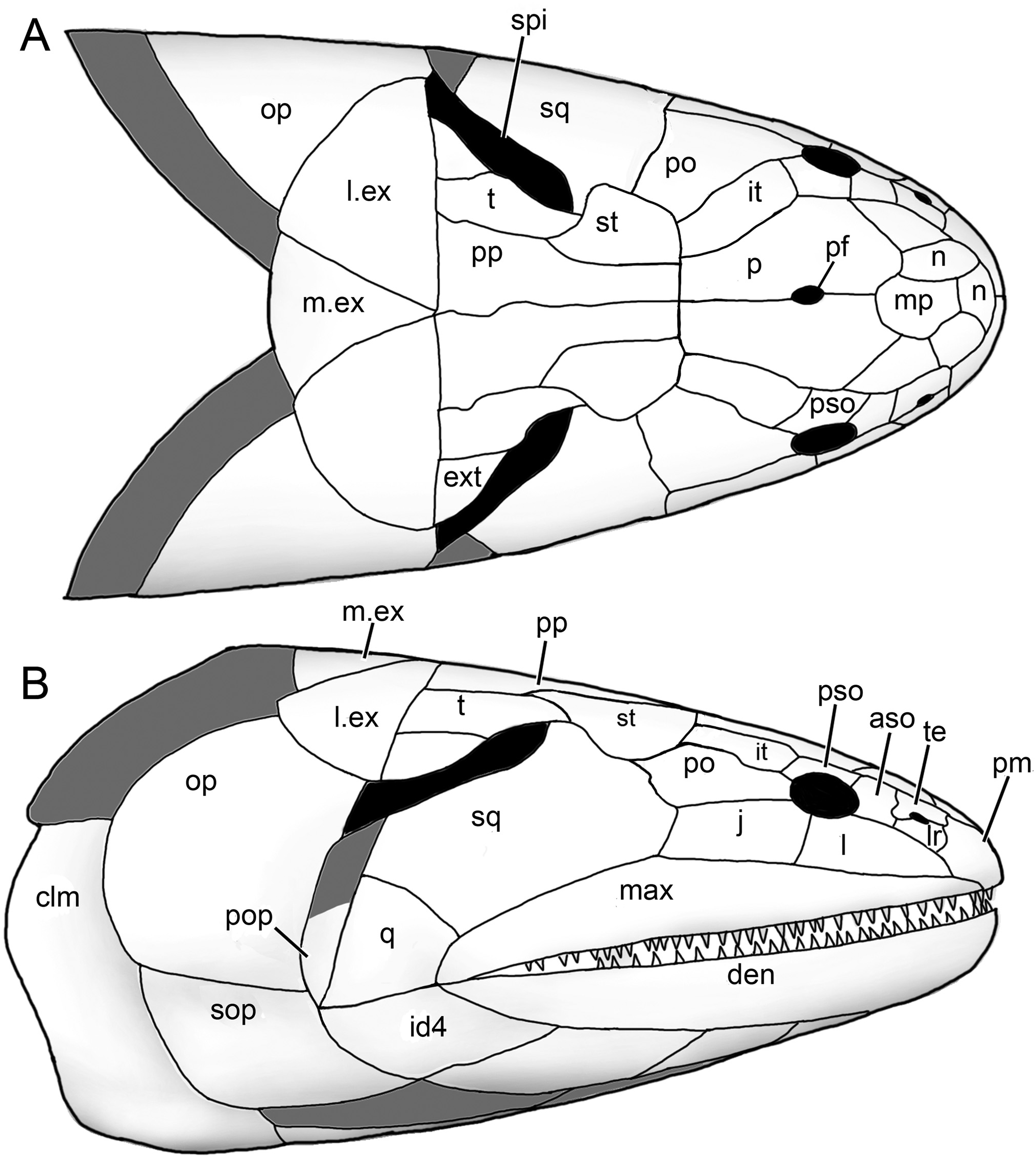
The holotype ( NTM P6410) preserves a head and fusiform body outline with a total length of 26.5 cm ( Fig. 2 View FIGURE 2 ). The posterior end of the fossil terminates shortly before the caudal fin, suggesting an original total length of roughly 30 cm, assuming similar proportions to those of Canowindra (Thomson, 1973) or Gogonasus (Long et al., 2006; Long & Trinajstic, 2017). Assuming minimal distortion of the incompletely preserved skull bones, the holotype skull length is about 4.8 cm from the snout to the posterior margin of the post-parietals. The largest skull, AMF 99704, has a length of 7.6 cm, suggesting a fish 45– 50 cm long ( Fig. 3G–H View FIGURE 3 ). Only a few small patches of cranial dermal ornament are preserved, with faint impressions suggesting wavy ridges on the skull roof and a vermiculate pattern of short ridges and pores on the snout ( Fig. 4A, B View FIGURE 4 ). Sensory canals are also rarely preserved, restricted to short sections of lateral line canals on the lateral extrascapular of NTM P6410 ( Fig. 3A, B View FIGURE 3 ) and AMF 99704 ( Fig. 3G, H View FIGURE 3 ) and on the squamosal of CPC 33948 ( Fig. 3C, D View FIGURE 3 ). A reconstruction of the complete skull, opercular-gular series, and pectoral girdle in articulation is presented in Figure 7 View FIGURE 7 .
Skull
Snout —Various regions of the snout are preserved throughout different specimens of Harajicadectes ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 5 View FIGURE 5 ), with the median post rostral and nasals best shown in NMV P228722 ( Fig. 3E, F View FIGURE 3 ). The median post rostral is an anteroposteriorly elongate hexagon, approximately half the length of the parietals, and is the width of a single parietal. It is laterally bordered on each side by smaller single nasal bones, which closely match the median post-rostral in shape. Lateral abutment of the median post rostral by single nasal bones is also present in Eusthenodon (Jarvik, 1952) and Cladarosymblema (Fox et al., 1995) , although the configuration and number of nasals varies within these taxa. At least two small sub-hexagonal nasals anteriorly straddle the median post-rostral. Both are approximately half the length of the median post rostral but equal in width. A small bone may sit in between these two anterior nasals, and is approximately 1/6 the width of each. A small rectangular anterior tectal and lateral rostral bones are situated between the anterior suborbital and premaxilla ( Figs. 3 View FIGURE 3 , 5 View FIGURE 5 ). A single pair of external nostrils was present, with the only unambiguous nostril visible on CPC 39949B, entirely enclosed by the anterior tectal and lateral rostral ( Fig. 4A, B View FIGURE 4 ).
Skull Roof —The parietal and postparietals of Harajicadectes ( Fig. 3 View FIGURE 3 ) are of roughly equal anteroposterior length. This is a condition also seen in tristichopterids (Schultze & Reed, 2012; Young et al., 2013) and elpistostegalians (Vorobyeva, 1980; Cloutier et al., 2020), whereas more basally-branching forms such as Gogonasus (Long et al., 1997) , Marsdenichthys ( Holland et al., 2010), and canowindrids (Long, 1985b) typically possess postparietals that are significantly longer than the parietals.
The parietal shield ( Fig. 3 View FIGURE 3 ) is well preserved in AMF 99704 and the paratype CPC 39948, while only the parietals are visible in NMV P228722. The parietals are elongate, sub-rectangular bones, being three times as long as wide and slightly smaller or equal in length to the postparietals, a condition seen in some specimens of Gyroptychius (Thomson, 1965) and tristichopterid taxa such as Mandageria (Johanson & Ahlberg, 1997) and Eusthenopteron (Jarvik, 1980) . The anterior margin of each parietal varies between being pointed, to laterally border the median post rostral, or concave to surround the posterior-most nasal bone. The median suture between the parietals is visible in CPC 39948 and NMV P228722, with the latter clearly showing a small circular opening for the pineal foramen, as opposed to the large kite-shaped pineal bone of tristichopterids such as Eusthenodon (Jarvik, 1952) or Mandageria (Johanson & Ahlberg, 1997) . In megalichthyids the pineal foramen is closed (Fox et al., 1995; Janvier et al., 2007; Thomson, 1964). The posterior end of both parietals is wider than the anterior edge of both postparietals. Parietal pit lines are not preserved.
The intertemporal is elongate, being three times as long as wide, and is approximately equal in length to the supratemporal and tabular of the postparietal shield. Towards the anterior end, the intertemporal widens, with a transverse anterior margin. This differs from the narrow, pointed anterior end of the intertemporal in tristichopterids (e.g., Eusthenopteron , Eusthenodon , Mandageria , Cabonnichthys , Edenopteron ; Ahlberg & Johanson, 1997; Jarvik, 1944, 1952; Johanson & Ahlberg, 1997; Young et al., 2019). The anterior end is also pointed in Cladarosymblema (Fox et al., 1995) , whereas an anteriorly broad intertemporal is seen in Marsdenichthys ( Holland et al., 2010) and Medoevia (Lebedev, 1995) . However, in the latter two taxa the intertemporal is relatively shorter compared with Harajicadectes . The ‘posterior postorbital’ in an equivalent position in Canowindra is also broader anteriorly and pointed posteriorly (Long, 1985b).
Both posterior and anterior supraorbitals are short, but broad. The posterior supraorbital has a blunt or rounded posterior end, distinct from the pointed corresponding region of the posterior supraorbital in Eusthenodon (Jarvik, 1952) , Mandageria (Johanson & Ahlberg, 1997) , and Cabonnichthys (Ahlberg & Johanson, 1997) . The right posterior supraorbital on CPC 33948 appears to be either broken or could be two separate bones ( Fig. 3C, D View FIGURE 3 ). A strip of narrow, fluted bone delineates the anterior margin of the postparietals ( Fig. 3C–H View FIGURE 3 ). Each postparietal has a narrow anterior section, and broadens posteriorly so the rear half of the bone is more than double the anterior breadth. The postparietals are joined at a gently undulating median suture, and the posterior margins are relatively straight, as in the Tristichopteridae (Jarvik, 1996; Johanson & Ahlberg, 1997; Young et al., 2013, 2019). In contrast, in Gogonasus (Long et al., 2006) , canowindrids (Long, 1985b, 1987), and the megalichthyid Litoptychius (Schultze & Chorn, 1998) , the posterior margin of the postparietal shield displays some form of curvature or projection. A faint pitline is visible crossing the left postparietal in NMVP 228722.
The supratemporals are located laterally to the anterior portion of each postparietal, with the best-preserved examples visible on CPC 33948 ( Fig. 3C, D View FIGURE 3 ) and AMF 99704 ( Fig. 3G, H View FIGURE 3 ). Each supratemporal is relatively large and roughly semi-circular, exhibiting a wide convex lateral border. This width is over double that of the anterior breadth of a postparietal. It is of roughly equal length to the tabular, in contrast to Glyptopomus (Jarvik, 1950) and Jarvikina (Vorobjeva, 1977) where the supratemporal is approximately double the length of the tabular.
The tabulars are elongate and sub-rectangular, about half the width of the supratemporal. This is distinct amongst tetrapodomorph fish except for the basally-branching rhizodontid Gooloogongia (Johanson & Ahlberg, 2001) . However, unlike Gooloogongia , the tabulars in Harajicadectes flank greatly enlarged external openings for the spiracular chambers. As in rhizodontids and ‘osteolepidids,’ the tabulars are bordered laterally by the extratemporals. These are triangular, and slightly wider than the tabulars. Canowindra also has a triangular extratemporal.
Cranial preservation of the holotype ( Figs. 2 View FIGURE 2 , 3A, B View FIGURE 3 ), CPC 33948 ( Fig. 3C, D View FIGURE 3 ) and CPC 39952 ( Fig. 5E, F View FIGURE 5 ) reveal the natural boundary between the skull roof and the cheek unit. The most conspicuous character is the greatly enlarged spiracular opening, accounting for between 25–30% of the total length of the skull roof. The opening is bordered posteromesially by the tabular and extratemporal, anterolaterally by the squamosal (and preopercular if present). The bony slit appears to have been open-ended posteriorly.
The spiracle is a minute slit in the majority of early osteichthyans (Jarvik, 1980). Among Devonian forms, enlarged spiracular openings are present in the marine tetrapodomorph Gogonasus (Long et al., 2006) , elpistostegalian-grade stem-tetrapods (Brazeau & Ahlberg, 2006; Cloutier et al., 2020; Daeschler et al., 2006) and the actinopterygian Pickeringius (Choo et al., 2019) . In Gogonasus (Long et al., 2006) , the tabulars have ventrally directed laminae forming a sloping wall to the enlarged spiracles, but these (if present) are not displayed in the tabulars of Harijicadectes.
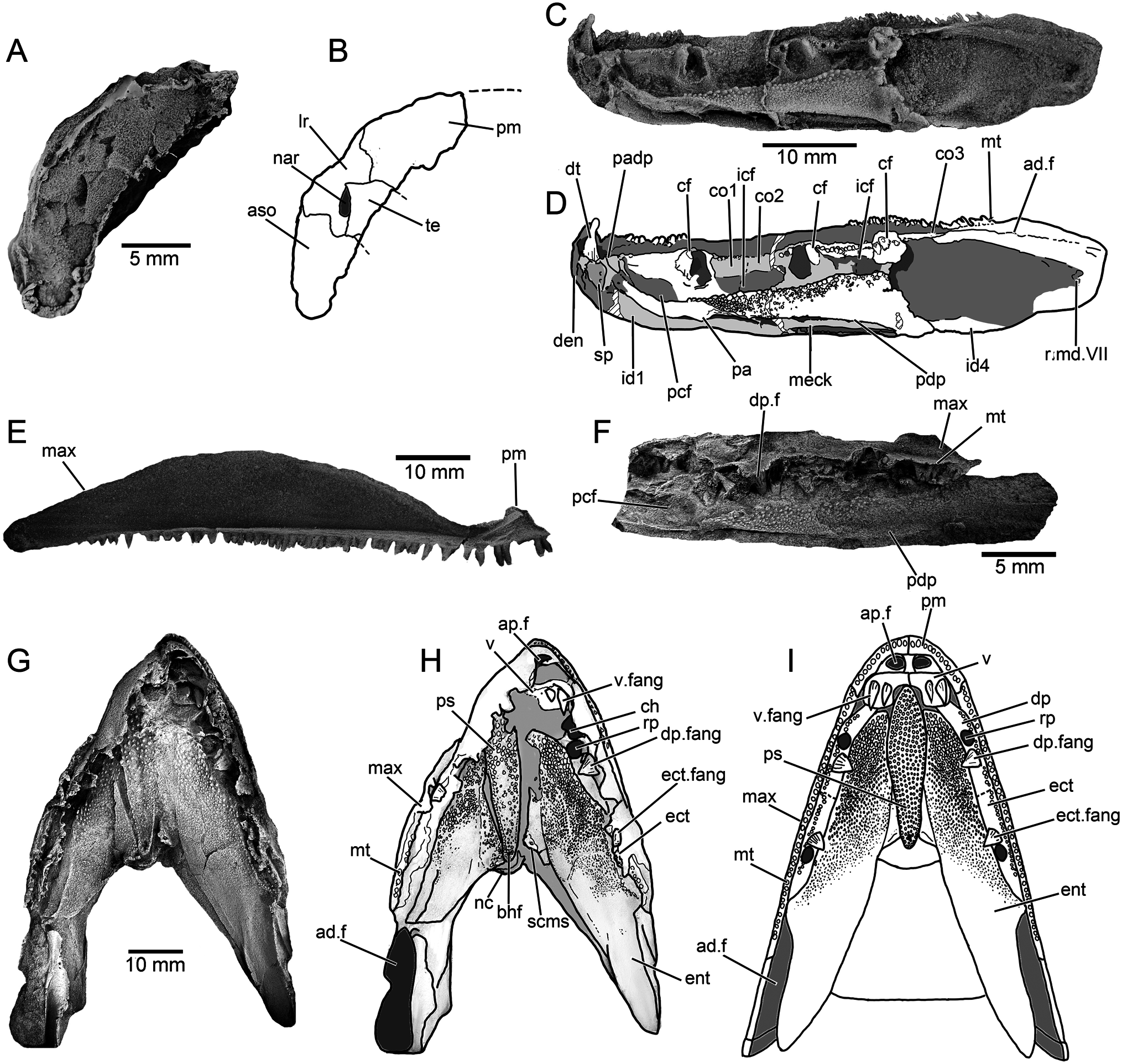
Cheek —The maxilla is preserved in articulation in a number of specimens ( Figs. 3A–F View FIGURE 3 , 5E, F View FIGURE 5 ), and NMV P253965 ( Fig. 4E View FIGURE 4 ) includes a complete disarticulated example in mesial view. It is an elongate bar-like bone, approximately equal in length to the parietal and postparietal combined. While there is individual variation in proportions, in all specimens the bone gently tapers anteriorly to a similar extent to the maxilla of Marsdenichthys ( Holland et al., 2010), but substantially less than that of Megalichthys (Miall, 1885) , Rhizodopsis (Traquair, 1881) , Gogonasus (Long et al., 1997) , and Latvius (Jessen, 1973) . In mesial view ( Fig. 4E View FIGURE 4 ), the bone flares dorsally at the contact with the premaxilla, indicating the presence of a substantial infraorbital overlap surface. Posteriorly, the maxilla tapers slightly towards a rounded tip. Numerous sharp, conical teeth are positioned along the ventral margin except for the posterior 15%. More than 45 teeth are present on NMV P253965. The internal surface of this specimen shows a continuous ridge diverging from the tooth row with an anterodorsally directed flange at the anterior extremity. This feature, which contributes to the posterior margin of the choana, is also present in Cladarosymblema (Fox et al., 1995) , Medoevia (Lebedev, 1995) , and Gogonasus (Long et al., 1997) .
The lachrymal ( Figs. 3C, D, G, H View FIGURE 3 , 5E, F View FIGURE 5 ) has a steep posterior suture with the jugal and has a short, posterodorsal concavity that contributes to the relatively small orbit. Based on CPC 33948 ( Fig. 3C, D View FIGURE 3 ), the jugal is rectangular in shape and of similar size to the lachrymal, with a narrow anterodorsal contribution to the orbit. The postorbital ( Fig. 3C, D, G, H View FIGURE 3 ) is preserved only in CPC 33948 and AMF 99704. Unlike Osteolepis and Gogonasus (Long et al., 1997) the postorbital does not extend posteriorly as far as the spiracular slit.
The squamosal is incompletely known, with partial examples preserved on CPC 33948 ( Fig. 3C, D View FIGURE 3 ), CPC 39952 ( Fig. 5E, F View FIGURE 5 ), and AMF 99704 ( Fig. 3G, H View FIGURE 3 ). It is a large bone, comparable in size to the entire postparietal shield. The entire posterodorsal margin forms most of the anterolateral margin of the spiracular slit, with a short anteromedial concavity to accommodate the lateral curvature of the supratemporal. The quadratojugal, visible only in CPC 39952 ( Fig. 5E, F View FIGURE 5 ), is a triangular bone, roughly 1/7th the size of the squamosal with which it sutures anterodorsally. An unambiguous preopercular has not been identified in the available material.
Extrascapulars —The extrascapular series ( Figs. 3 View FIGURE 3 , 5 View FIGURE 5 ) consists of paired lateral and a single median element. One complete lateral extrascapular is preserved in position in CPC 33948 ( Fig. 3C, D View FIGURE 3 ) and NMVP 228722 ( Fig. 3E, F View FIGURE 3 ) and a partial median extrascapular is visible on CPC 39952. The lateral units are large, semicircular bones, of comparable size to the parietal shield. NMVP 228722 shows ornamental grooves radiating from the ossification center of the bone. The median extrascapular is a triangular bone, about half the size of the laterals, which tapers towards an anterior point.
Palate and Ethmosphenoid — CPC 33949A ( Fig. 4G–I View FIGURE 4 ) preserves the best example of the palatal anatomy. The parasphenoid is an elongate, lanceolate element with a denticulated surface that tapers towards a rounded posterior extremity. The anterior tip is not clearly visible, but the lateral contours and posterior embayment formed by the vomers indicate that it gently tapers towards the front. The denticles are distinctly larger on the anterior half of the bone, with a slightly raised single row of denticles along the posterolateral margins surrounding a central depression that housed the buccohypophyseal foramen.
A worn right vomer is visible in its natural position, bearing two pronounced conical fangs. The vomer consists of a broad, lateral section with a straight posterior margin, lacking the tapering posterior projections present in Eusthenopteron (Jarvik, 1980) and mandageriines (Young et al., 2013, 2019). A narrow anteromedial ramus projects towards a presumed contact with the opposite vomer, forming an embayment to accommodate the anterior tip of the parasphenoid. The vomer of Megalichthys also possesses a medially directed extension (Jarvik, 1966). In that taxon the bone gently tapers towards the midline of the palate, rather than being sharply separated into a narrow medial and broad lateral sections as in Harajicadectes .
The denticulated surfaces of both entopterygoids are preserved, although heavily fractured due to post-mortem compression. A wedge-shaped mass of bone to the right of the parasphenoid (left in palatal view) might represent a discrete element ( Fig. 4G View FIGURE 4 ), perhaps comparable to the ‘accessory vomers’ first described from the Canowindra tristichopterids Mandageria and Cabonnichthys (Ahlberg & Johanson, 1997; Johanson & Ahlberg, 1997). In the absence of an unambiguous counterpart on the left side and without supporting evidence from additional specimens, this is currently interpreted as a fractured section of the entopterygoid. Denticles are larger in the anterior parts of the entopterygoids, particularly along the lateral margins, becoming progressively finer and more scattered towards the posterior, with the rear quarter of the bone being almost devoid of ornamentation. The dermopalatines and ectopterygoids are preserved, but the suture between them is difficult to discern. Each bone bears a single stout, conical fang, mesial to a row of fine denticles.
Small sections of the ethmosphenoid floor are visible adjacent to the parasphenoid, although insufficiently preserved to accurately determine the overall shape. A broken section to the left (right in palatal view) of the parasphenoid and partially underlain by the entopterygoid bears a prominent subcranial muscle scar. A roughened semicircular surface immediately posterior of the parasphenoid likely represents the rim of the notochordal chamber.
Lower Jaw —The labial surface of the mandible is partially visible in the holotype ( Fig. 3A, B View FIGURE 3 ), CPC 39952 ( Fig. 5E, F View FIGURE 5 ), and a variety of isolated heads ( Fig. 3C–H View FIGURE 3 ). The lingual surface is reasonably complete in NMV P253964 ( Fig. 4C, D View FIGURE 4 ), the only specimen that preserves the entire outline of the mandible, and partially visible in NMV P253965A ( Fig. 4F View FIGURE 4 ). NMV P253964 has a straight occlusal margin and a gently convex ventral surface. It tapers only very gently from back to front, so the anterior jaw tip is only marginally narrower than the rear. The occlusal margin has small conical teeth, arranged in at least two rows, from the anterior tip to over the anterior-third of the adductor fossa. A single large, recurved symphysial tusk is present at the anterior of the dentary, 4–5 times the size of the marginal teeth.
The prearticular of NMV P253964, while missing the rear end, is seen to expand posteriorly, gently narrowing anteriorly and bending dorsally, terminating shortly before the median symphysis. Part of the denticulated prearticular plate is preserved, the denticles being substantially larger along the dorsal edge. A highly fragmentary parasymphysial plate is preserved immediately anterior to the prearticular. Three coronoids appear to be present, but it is difficult to discern the sutures between them and the third in the series is poorly preserved. A continuous vertical lamina bearing a row of closely packed denticles runs along the entire coronoid series. Positioned labially to this lamina, each coronoid bears large, stout fangs and adjacent replacement pits.
Body and Fins
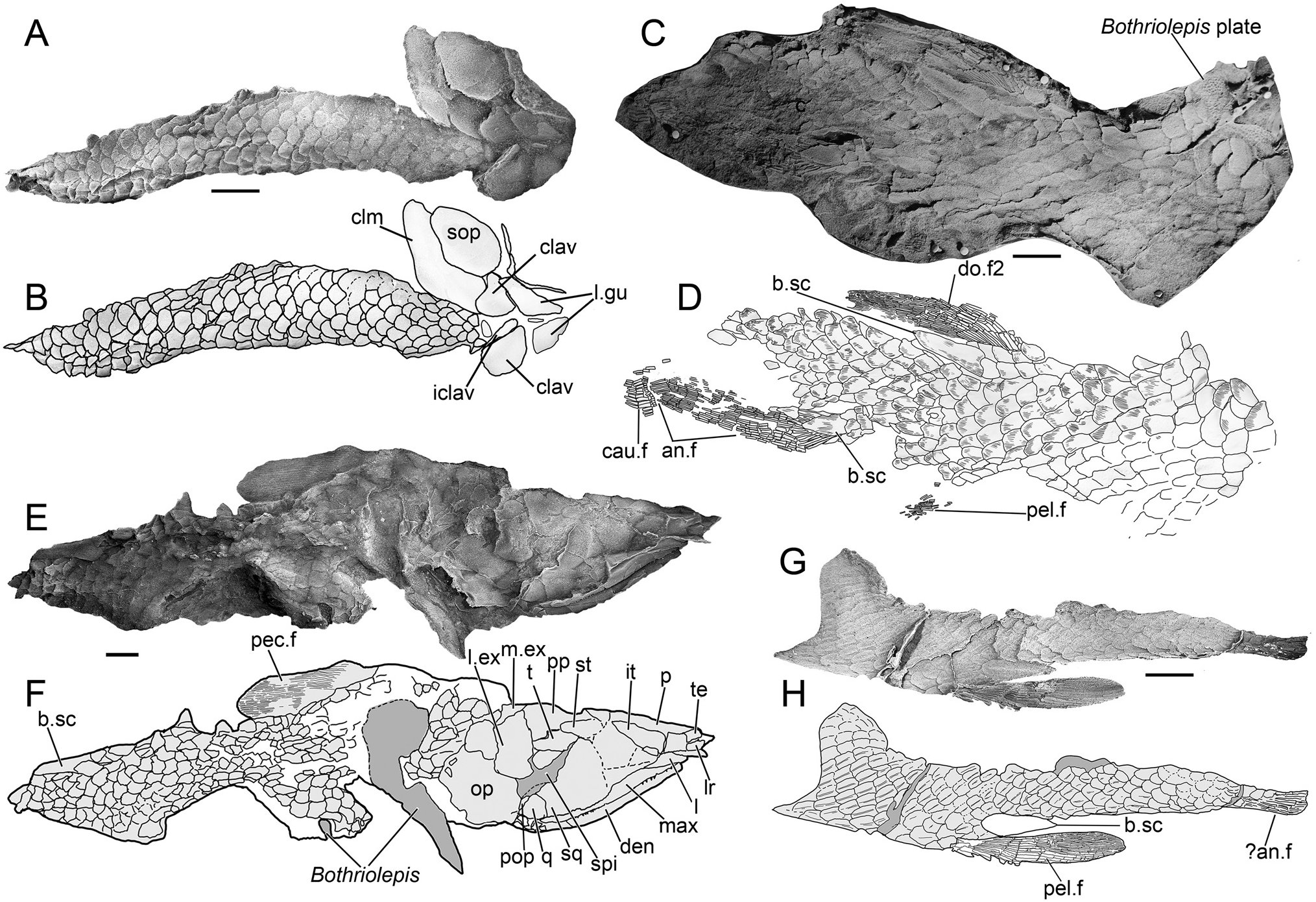
Scales —The scales of Harajicadectes are found in direct association with cranial material in the holotype ( Fig. 1 View FIGURE 1 ) and CPC 39952 ( Fig. 5E, F View FIGURE 5 ), enabling the identification of headless, scale-bearing specimens ( CPC 39951, 39953, 39954, NMVP 229368, NTM P6411) as this taxon. The squamation is cycloid in the anterior and mid-sections of the postcranium ( Fig. 5A, B, E, F View FIGURE 5 ), abruptly shifting to a rhombic shape in the vertical rows immediately behind the level of the second dorsal fin ( Fig. 5D–G View FIGURE 5 ). This shift in scale morphology from rounded to angular is unusual for a tetrapodomorph fish, but is also recorded in Marsdenichthys ( Holland et al., 2010) and Rhizodopsis (Andrews & Westoll, 1970) .
The exposed external surfaces of the scales lack cosmine and are ornamented, as in rhizodontids ( Holland et al., 2007), tristichopterids (Jarvik, 1952), Marsdenichthys ( Holland et al., 2010), Medoevia (Lebedev, 1995) , Rhizodopsis (Schultze & Heidtke, 1986) and Canowindra (Thomson, 1973) . Each scale has approximately 25-35 anteroposteriorly directed sub-parallel ridges ( Fig. 6 View FIGURE 6 ), which appear wider and less densely packed than those of Marsdenichthys ( Holland et al., 2010). This morphology differs from that of Mandageria , Cabonnichthys , and Notorhizodon , in which scale ornamentation is composed of widely separated parallel grooves (Young, 2008; Young et al., 2013). Of a selection of isolated tetrapodomorph scales figured by Burrow and Turner (2012) from Emsian?/Eifelian Cravens Peak Beds, Queensland, two specimens ( ANU V3534 and V3535) display similar ornamentation to Harajicadectes .
In Harajicadectes , the course of these ridges becomes rounded, following the curvature of the scale border. The concentric intersecting lines characteristic of Marsdenichthys and Rhizodopsis ( Holland et al., 2010) are absent.
Fins —No endoskeletal fin elements are preserved in any of the material. A left pectoral fin is present on CPC 39952 ( Fig. 5E, F View FIGURE 5 ) while CPC 39954 displays a probable right pectoral fin with an associated basal scute which appears to have detached and flipped post-mortem to lie against the internal surfaces of squamation from the right side of the body ( Fig. 5G, H View FIGURE 5 ). The fin is paddle-shaped with a rounded tip, with over 19 unsegmented proximal lepidotrichia that transition into finer rays towards the distal tip. The base of the fin is poorly preserved and the disposition of the fleshy lobe and basal scute is unclear. The fin is narrower than the pectoral fin, with over 12 elongate proximal lepidotrichia which progressively divide into over 25 fine distal fin rays. Fragmentary scales, smaller than the adjacent trunk scales, cover the proximal fin rays, suggesting the presence of a fleshy basal lobe, although its size and shape are indeterminate.
CPC 39953 preserves the anal and second dorsal fins along with small fragments of the pelvic and caudal fin. No lepidotrichia of the first dorsal fin are visible, although CPC 39952 preserves a basal scute ( Fig. 5E, F View FIGURE 5 ). The second dorsal and anal fins are elongate and narrow based, with associated basal scutes longer than the fin bases. The anal fin is positioned posterior to the second dorsal fin, at the transition zone from the cycloid scales of the mid-body to the rhombic scales on the posterior. It has a rounded tip; the distal end of the dorsal fin is damaged.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.